
Abstract
Pre-eclampsia (typically characterized by new-onset hypertension and proteinuria in the second half of pregnancy) represents a major determinant of the global burden of disease1,2. Its pathophysiology involves placental dysfunction, but the mechanism is unclear. Viral infection can cause organ dysfunction, but its role in placentally related disorders of human pregnancy is unknown3. We addressed this using RNA sequencing metagenomics4,5,6 of placental samples from normal and complicated pregnancies. Here, we show that human herpesvirus 6 (HHV-6, A or B) RNA was detected in 6.1% of cases of pre-eclampsia and 2.2% of other pregnancies. Fetal genotyping demonstrated that 70% of samples with HHV-6 RNA in the placenta exhibited inherited, chromosomally integrated HHV-6 (iciHHV-6). We genotyped 467 pre-eclampsia cases and 3,854 controls and found an excess of iciHHV-6 in the cases (odds ratio of 2.8, 95% confidence intervals of 1.4–5.6, P = 0.008). We validated this finding by comparing iciHHV-6 in a further 740 cases with controls from large-scale population studies (odds ratio of 2.5, 95% confidence intervals of 1.4–4.4, P = 0.0013). We conclude that iciHHV-6 results in the transcription of viral RNA in the human placenta and predisposes the mother to pre-eclampsia.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 digital issues and online access to articles
$119.00 per year
only $9.92 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout


Similar content being viewed by others
Data availability
The sequencing data have been deposited in the European Genome-phenome Archive (EGA) with the following accession numbers: EGAD00001003457 (RNA-seq controls), EGAD00001003507 (RNA-seq FGR cases), EGAD00001003508 (RNA-seq pre-eclampsia cases), EGAD00001004197 (metagenomics) and EGAD00001004592 (SureSelect).
Code availability
Custom scripts for the bioinformatics analyses are available at https://github.com/sung/HHV6-Nat-Micr-2020.
References
GBD 2015 Maternal Mortality Collaborators. Global, regional, and national levels of maternal mortality, 1990–2015: a systematic analysis for the Global Burden of Disease Study 2015. Lancet 388, 1775–1812 (2016).
McClure, E. M. et al. Global Network for Women’s and Children’s Health Research: probable causes of stillbirth in low- and middle-income countries using a prospectively defined classification system. BJOG 125, 131–138 (2018).
Brosens, I., Pijnenborg, R., Vercruysse, L. & Romero, R. The ‘Great Obstetrical Syndromes’ are associated with disorders of deep placentation. Am. J. Obstet. Gynecol. 204, 193–201 (2011).
Gardy, J. L. & Loman, N. J. Towards a genomics-informed, real-time, global pathogen surveillance system. Nat. Rev. Genet. 19, 9–20 (2018).
Houldcroft, C. J., Beale, M. A. & Breuer, J. Clinical and biological insights from viral genome sequencing. Nat. Rev. Microbiol. 15, 183–192 (2017).
Depledge, D. P. et al. Specific capture and whole-genome sequencing of viruses from clinical samples. PLoS ONE 6, e27805 (2011).
Gravel, A., Hall, C. B. & Flamand, L. Sequence analysis of transplacentally acquired human herpesvirus 6 DNA is consistent with transmission of a chromosomally integrated reactivated virus. J. Infect. Dis. 207, 1585–1589 (2013).
Pantry, S. N. & Medveczky, P. G. Latency, integration, and reactivation of human herpesvirus-6. Viruses 9, 194 (2017).
Huang, Y. et al. Human telomeres that carry an integrated copy of human herpesvirus 6 are often short and unstable, facilitating release of the viral genome from the chromosome. Nucleic Acids Res. 42, 315–327 (2014).
GOPEC Consortium. Disentangling fetal and maternal susceptibility for pre-eclampsia: a British multicenter candidate-gene study. Am. J. Hum. Genet. 77, 127–131 (2005).
Tanaka-Taya, K. et al. Human herpesvirus 6 (HHV-6) is transmitted from parent to child in an integrated form and characterization of cases with chromosomally integrated HHV-6 DNA. J. Med. Virol. 73, 465–473 (2004).
Miura, H. et al. Chromosomally integrated human herpesvirus 6 in the Japanese population. J. Med. Virol. 90, 1636–1642 (2018).
Gravel, A. et al. Inherited chromosomally integrated human herpesvirus 6 as a predisposing risk factor for the development of angina pectoris. Proc. Natl Acad. Sci. USA 112, 8058–8063 (2015).
Sovio, U. et al. Prediction of preeclampsia using the soluble fms-like tyrosine kinase 1 to placental growth factor ratio: a prospective cohort study of unselected nulliparous women. Hypertension 69, 731–738 (2017).
Sultana, Z., Maiti, K., Dedman, L. & Smith, R. Is there a role for placental senescence in the genesis of obstetric complications and fetal growth restriction? Am. J. Obstet. Gynecol. 218, S762–S773 (2018).
Broady, A. J. et al. Protective proteins and telomere length in placentas from patients with pre-eclampsia in the last trimester of gestation. Placenta 50, 44–52 (2017).
Sano, R. & Reed, J. C. ER stress-induced cell death mechanisms. Biochim. Biophys. Acta 1833, 3460–3470 (2013).
Burton, G. J., Yung, H. W., Cindrova-Davies, T. & Charnock-Jones, D. S. Placental endoplasmic reticulum stress and oxidative stress in the pathophysiology of unexplained intrauterine growth restriction and early onset preeclampsia. Placenta 30 (Suppl. A), S43–S48 (2009).
Racicot, K. & Mor, G. Risks associated with viral infections during pregnancy. J. Clin. Invest. 127, 1591–1599 (2017).
Zhang, E. et al. Inherited chromosomally integrated human herpesvirus 6 genomes are ancient, intact, and potentially able to reactivate from telomeres. J. Virol. 91, e01137-17 (2017).
Peddu, V. et al. Inherited chromosomally integrated HHV-6 demonstrates tissue-specific RNA expression in vivo that correlates with increased antibody immune response. J. Virol. 94, e01418-19 (2019).
Marci, R. et al. Presence of HHV-6A in endometrial epithelial cells from women with primary unexplained infertility. PLoS ONE 11, e0158304 (2016).
Caselli, E. et al. HHV-6A infection of endometrial epithelial cells induces increased endometrial NK cell-mediated cytotoxicity. Front. Microbiol. 8, 2525 (2017).
Dekker, G., Robillard, P. Y. & Roberts, C. The etiology of preeclampsia: the role of the father. J. Reprod. Immunol. 89, 126–132 (2011).
Smith, G. C., Pell, J. P. & Walsh, D. Pregnancy complications and maternal risk of ischaemic heart disease: a retrospective cohort study of 129,290 births. Lancet 357, 2002–2006 (2001).
Bellamy, L., Casas, J. P., Hingorani, A. D. & Williams, D. J. Pre-eclampsia and risk of cardiovascular disease and cancer in later life: systematic review and meta-analysis. BMJ 335, 974 (2007).
Pasupathy, D. et al. Study protocol. A prospective cohort study of unselected primiparous women: the pregnancy outcome prediction study. BMC Pregnancy Childbirth 8, 51 (2008).
Sovio, U., White, I. R., Dacey, A., Pasupathy, D. & Smith, G. C. Screening for fetal growth restriction with universal third trimester ultrasonography in nulliparous women in the Pregnancy Outcome Prediction (POP) study: a prospective cohort study. Lancet 386, 2089–2097 (2015).
Gaccioli, F., Lager, S., Sovio, U., Charnock-Jones, D. S. & Smith, G. C. S. The pregnancy outcome prediction (POP) study: investigating the relationship between serial prenatal ultrasonography, biomarkers, placental phenotype and adverse pregnancy outcomes. Placenta 59, S17–S25 (2017).
ACOG. Hypertension in pregnancy. Report of the American College of Obstetricians and Gynecologists’ task force on hypertension in pregnancy. Obstet. Gynecol. 122, 1122–1131 (2013).
Hiby, S. E. et al. Combinations of maternal KIR and fetal HLA-C genes influence the risk of preeclampsia and reproductive success. J. Exp. Med. 200, 957–965 (2004).
Pellett, P. E. et al. Chromosomally integrated human herpesvirus 6: questions and answers. Rev. Med. Virol. 22, 144–155 (2012).
Hill, J. A. et al. Outcomes of hematopoietic cell transplantation using donors or recipients with inherited chromosomally integrated HHV-6. Blood 130, 1062–1069 (2017).
Moustafa, A. et al. The blood DNA virome in 8,000 humans. PLoS Pathog. 13, e1006292 (2017).
Mouammine, A. et al. rs73185306 C/T is not a predisposing risk factor for inherited chromosomally-integrated human herpesvirus 6A/B. J. Infect. Dis. 221, 878–881 (2020).
Gardosi, J., Mongelli, M., Wilcox, M. & Chang, A. An adjustable fetal weight standard. Ultrasound Obstet. Gynecol. 6, 168–174 (1995).
Langmead, B. & Salzberg, S. L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 9, 357–359 (2012).
Kim, D. et al. TopHat2: accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 14, R36 (2013).
Krueger, F. Trim Galore!: a wrapper tool around Cutadapt and FastQC to consistently apply quality and adapter trimming to FastQ files, with some extra functionality for MspI-digested RRBS-type (Reduced Representation Bisufite-Seq) libraries. (Babraham Bioinformatics, 2012); http://www.bioinformatics.babraham.ac.uk/projects/trim_galore/
Wood, D. E. & Salzberg, S. L. Kraken: ultrafast metagenomic sequence classification using exact alignments. Genome Biol. 15, R46 (2014).
Li, H. & Durbin, R. Fast and accurate short read alignment with Burrows–Wheeler transform. Bioinformatics 25, 1754–1760 (2009).
Carver, T., Harris, S. R., Berriman, M., Parkhill, J. & McQuillan, J. A. Artemis: an integrated platform for visualization and analysis of high-throughput sequence-based experimental data. Bioinformatics 28, 464–469 (2012).
Lager, S. et al. Detecting eukaryotic microbiota with single-cell sensitivity in human tissue. Microbiome 6, 151 (2018).
Mori, Y., Akkapaiboon, P., Yang, X. & Yamanishi, K. The human herpesvirus 6 U100 gene product is the third component of the gH–gL glycoprotein complex on the viral envelope. J. Virol. 77, 2452–2458 (2003).
Brown, J. R. et al. Norovirus whole-genome sequencing by SureSelect target enrichment: a robust and sensitive method. J. Clin. Microbiol. 54, 2530–2537 (2016).
Nurk, S. et al. Assembling single-cell genomes and mini-metagenomes from chimeric MDA products. J. Comput. Biol. 20, 714–737 (2013).
Telford, M., Navarro, A. & Santpere, G. Whole genome diversity of inherited chromosomally integrated HHV-6 derived from healthy individuals of diverse geographic origin. Sci. Rep. 8, 3472 (2018).
Wood, M. L. & Royle, N. J. Chromosomally integrated human herpesvirus 6: models of viral genome release from the telomere and impacts on human health. Viruses 9, 184 (2017).
Boratyn, G. M. et al. BLAST: a more efficient report with usability improvements. Nucleic Acids Res. 41, W29–W33 (2013).
Fagerland, M. W., Lydersen, S. & Laake, P. Recommended confidence intervals for two independent binomial proportions. Stat. Methods Med. Res. 24, 224–254 (2015).
Lydersen, S., Fagerland, M. W. & Laake, P. Recommended tests for association in 2 × 2 tables. Stat. Med. 28, 1159–1175 (2009).
Nyaga, V. N., Arbyn, M. & Aerts, M. Metaprop: a Stata command to perform meta-analysis of binomial data. Arch. Public Health 72, 39 (2014).
Dominguez, G. et al. Human herpesvirus 6B genome sequence: coding content and comparison with human herpesvirus 6A. J. Virol. 73, 8040–8052 (1999).
Acknowledgements
We are grateful to the following: the participants of the POP, case–control and GOPEC studies; L. Bibby, S. Ranawaka, K. Holmes, J. Gill, C. L. Soh and R. Millar for technical assistance; the UCL Pathogen Genomics Unit; the Sanger Institute core sequencing facility and the Pathogen Informatics team. This work was supported by the Women’s Health theme of the NIHR Cambridge Biomedical Research Centre, the Medical Research Council (MR/K021133/1 to G.C.S.S., D.S.C.-J., J.P. and S.J.P. and G1100221 to G.C.S.S. and D.S.C.-J.) and the UCL/UCLH NIHR Biomedical Research Centre (to J.B. and C.V.). S.L., M.C.d.G., J.D., S.J.P., J.P., D.S.C.-J. and G.C.S.S. received grants from the Medical Research Council (UK); F.G., U.S., S.G., E.C., D.S.C.-J. and G.C.S.S. received grants from the National Institute for Health Research (UK); U.S. received grants from the Stillbirth & Neonatal Death Society (Sands); and J.P. received grants from the Wellcome Trust and grants from Pfizer.
Author information
Authors and Affiliations
Contributions
G.C.S.S. had the original idea. G.C.S.S., D.S.C.-J., J.B., J.P. and S.J.P. supervised the study. G.C.S.S., D.S.C.-J., J.B., J.P., S.J.P., F.G., S.L. and M.C.d.G. designed the experiments. F.G. and J.D. performed the RNA-seq experiments. F.G. and E.C. performed the cord and parental DNA genotyping experiments. F.G. analysed the placental RNA samples by RT–qPCR. S.L. performed the qPCR experiments on placental DNA samples and the DNA-seq experiments. M.C.d.G., S.G. and C.V. analysed the RNA-seq and DNA-seq data. U.S. performed statistical analyses. A.M. and A.S. provided samples and clinical information for the case–control study. C.D. and W.K.L. provided samples and clinical information for the GOPEC study. All authors reviewed the paper prior to submission.
Corresponding author
Ethics declarations
Competing interests
J.D. reports being an employee of GlaxoSmithKline and A.S. reports being an employee of Robinson College (Cambridge, UK). J.P. reports personal fees from Next Gen Diagnostics Llc., outside the submitted work; S.J.P. reports personal fees from Specific and personal fees from Next Gen Diagnostics, outside the submitted work; D.S.C.-J. reports grants from GlaxoSmithKline Research and Development Limited, outside the submitted work; G.C.S.S. reports grants and personal fees from GlaxoSmithKline Research and Development Limited, personal fees and non-financial support from Roche Diagnostics Ltd, outside the submitted work; D.S.C.-J. and G.C.S.S. report grants from Sera Prognostics Inc. and non-financial support from Illumina Inc., outside the submitted work. A.M., W.K.L., C.D., C.V. and J.B. have nothing to disclose.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Characteristics of the study groups in the Pregnancy Outcome Prediction (POP) study.
Data are expressed as median (IQR) or n (%) as appropriate. The overall rate of pre-eclampsia in these participants was 6.4%. For fields where there is no category labelled ‘missing’, data were 100% complete. Maternal age was defined as age at recruitment. All other maternal characteristics were defined by self-report at the 20 weeks questionnaire, from examination of the clinical case record, or linkage to the hospital’s electronic databases. Socio-economic status was quantified using the Index of Multiple Deprivation (IMD) 2007, which is based on census data from the area of the mother’s postcode. Stillbirths (n=8) and spontaneous preterm deliveries (n=100) were included in the analysis, while miscarriages (n=7) and terminations of pregnancy (n=11) were excluded. Abbreviations: non-cases denote patients without pre-eclampsia; FTE denotes full time education; BMI denotes body mass index; DM denotes diabetes mellitus.
Extended Data Fig. 2 Placental RNA-seq reads mapped to HHV-6A and HHV-6B genomes.
Placental HHV-6 positive samples identified by RNA-seq. Reads were identified by Kraken as aligning to the HHV-6 genomes and mapped with BWA to the HHV-6A or HHV-6B reference genomes (Supplementary Methods); note that the total number of reads/sample recognized by the two software is not always identical. CON denotes a healthy pregnancy without FGR or pre-eclampsia (see Methods); FGR denotes fetal growth restriction; PE denotes a patient with pre-eclampsia; HHV-6A denotes human herpesvirus 6, variant A; HHV-6B denotes human herpesvirus 6, variant B; DRL: direct repeat left; DRR: direct repeat right. Repetitive regions are in Italic. HHV-6A and HHV-6B genomes have been described by Dominguez G et al53.
Extended Data Fig. 3 HHV-6 detection in fetal and parental samples.
HHV-6A and HHV-6B representative signals in cord (a) and parental (b) DNA samples detected using a multiplex qPCR approach. These analyses were performed in 5,061 and 86 samples, respectively, and each sample was analyzed in triplicate. qPCR amplification curves for the HHV-6A and HHV-6B 9 U67/68 genes are represented in green and red, respectively; RNase P curves are in blue and confirmed presence of DNA in the wells. ciHHV-6 corresponds to a high HHV-6 DNA signal in the sample measured by qPCR, that is within 4 cycles of the RNase P signal. HHV-6 non-integrated corresponds to a HHV-6 DNA signal in the sample detected at more than 4 Ct higher compared to the RNase P signal. Negative samples lack HHV-6A or HHV-6B DNA signal. c) RT-qPCR amplification plot of placental RNA samples showing detection of the HHV-6 U100 gene. Eight representative samples are shown, two with viral transcript amplification (total n=48 samples, each analyzed in triplicate). Five negative controls (samples without reverse transcriptase enzyme in the RT reaction) lacked U100 amplification (not shown). U100 curves are in black and RNase P curves are in blue. Rn (normalized reporter value) represents the fluorescence of the reporter dye normalized to the signal of the passive reference dye for a given reaction. The ΔRn is the Rn value of an experimental reaction minus the Rn value of the baseline signal generated by the instrument. This parameter indicates the magnitude of the fluorescent signal generated in the qPCR assay. ciHHV-6 denotes chromosomally integrated human herpesvirus 6; HHV-6A denotes human herpesvirus 6, variant A; HHV-6B denotes human herpesvirus 6, variant B; RNase P denotes the human positive control gene RPPH1.
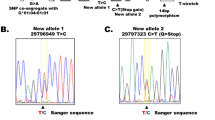
Extended Data Fig. 4 Identification of informative HHV-6B SNPs.
DNA-seq reads of 2 randomly selected samples were compared to the HHV-6B reference genome. *Informative SNP sites, that is SNPs present in just one of the two analyzed samples (gapped vertical red lines). **SNPs present in both analyzed samples, that is sites concordantly different from the reference genome, were considered not informative (continuous vertical red lines). Throughout the 162kb HHV-6B genome 187 SNPs were classified as informative.
Supplementary information
Supplementary Information
Supplementary Tables 1 and 2, and supplementary references.
Rights and permissions
About this article

Cite this article
Gaccioli, F., Lager, S., de Goffau, M.C. et al. Fetal inheritance of chromosomally integrated human herpesvirus 6 predisposes the mother to pre-eclampsia. Nat Microbiol 5, 901–908 (2020). https://doi.org/10.1038/s41564-020-0711-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41564-020-0711-3
This article is cited by
-
The immediate-early protein 1 of human herpesvirus 6B interacts with NBS1 and inhibits ATM signaling
EMBO Reports (2024)
-
Disease Burden of Primary HHV-6B Infection in Immunocompetent Children
Current Clinical Microbiology Reports (2024)
-
Genomic investigations of unexplained acute hepatitis in children
Nature (2023)
-
Maternal natural killer cells at the intersection between reproduction and mucosal immunity
Mucosal Immunology (2021)
