
Abstract
The peritrophic matrix of blood-feeding insects is a chitinous structure that forms a protective barrier against oral pathogens and abrasive particles1. Tsetse flies transmit Trypanosoma brucei, which is the parasite that causes human sleeping sickness and is also partially responsible for animal trypanosomiasis in Sub-Saharan Africa. For this parasite to establish an infection in flies, it must first colonize the area between the peritrophic matrix and gut epithelium called the ectoperitrophic space. Although unproven, it is generally accepted that trypanosomes reach the ectoperitrophic space by penetrating the peritrophic matrix in the anterior midgut2,3,4. Here, we revisited this event using fluorescence- and electron-microscopy methodologies. We show that trypanosomes penetrate the ectoperitrophic space in which the newly made peritrophic matrix is synthesized by the proventriculus. Our model describes how these proventriculus-colonizing parasites can either migrate to the ectoperitrophic space or become trapped within peritrophic matrix layers to form cyst-like bodies that are passively pushed along the gut as the matrix gets remodelled. Furthermore, early proventricular colonization seems to be promoted by factors in trypanosome-infected blood that cause higher salivary gland infections and potentially increase parasite transmission.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 digital issues and online access to articles
$119.00 per year
only $9.92 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others


Data availability
The data and statistics that support the findings of this study are available as Source Data files.
References
Terra, W. R. The origin and functions of the insect peritrophic membrane and peritrophic gel. Arch. Insect Biochem. Physiol. 47, 47–61 (2001).
Ellis, D. S. & Evans, D. A. Passage of Trypanosoma brucei rhodesiense through the peritrophic membrane of Glossina morsitans morsitans. Nature 267, 834–835 (1977).
Evans, D. A. & Ellis, D. S. Recent observations on the behaviour of certain trypanosomes within their insect hosts. Adv. Parasitol. 22, 1–42 (1983).
Evans, D. A. & Ellis, D. S. The penetrative ability of sleeping-sickness trypanosomes. Trans. R. Soc. Trop. Med. Hyg. 72, 653–655 (1978).
Matthews, K. R. The developmental cell biology of Trypanosoma brucei. J. Cell Sci. 118, 283–290 (2005).
Acosta-Serrano, A. et al. The surface coat of procyclic Trypanosoma brucei: programmed expression and proteolytic cleavage of procyclin in the tsetse fly. Proc. Natl Acad. Sci. USA 98, 1513–1518 (2001).
Vassella, E. et al. A major surface glycoprotein of Trypanosoma brucei is expressed transiently during development and can be regulated post-transcriptionally by glycerol or hypoxia. Genes Dev. 14, 615–626 (2000).
Van Den Abbeele, J., Claes, Y., Van Bockstaele, D., Le Ray, D. & Coosemans, M. Trypanosoma brucei spp. development in the tsetse fly: characterization of the post-mesocyclic stages in the foregut and proboscis. Parasitology 118, 469–478 (1999).
Vickerman, K., Tetley, L., Hendry, K. A. & Turner, C. M. Biology of African trypanosomes in the tsetse fly. Biol. Cell 64, 109–119 (1988).
Kleine, F. K. Weitere wissenschaftliche beobachtungen über die entwicklung von trypanosomen in Glossinen. Dtsch. Med. Wochenschr. 35, 924–925 (1909).
Hao, Z., Kasumba, I. & Aksoy, S. Proventriculus (cardia) plays a crucial role in immunity in tsetse fly (Diptera: Glossinidiae). Insect Biochem. Mol. Biol. 33, 1155–1164 (2003).
Billingsley, P. F. & Lehane, M. J. in Biology of the Insect Midgut (eds Lehane, M. J. & Billingsley, P. F.) Ch. 1, 3–30 (Springer, 1996).
Harmsen, R. The nature of the establishment barrier for Trypanosoma brucei in the gut of Glossina pallidipes. Trans. R. Soc. Trop. Med. Hyg. 67, 364–373 (1973).
Willett, K. C. Development of peritrophic membrane in Glossina (tsetse flies) and its relation to infection with trypanosomes. Exp. Parasitol. 18, 290–295 (1966).
Lehane, M. J. & Msangi, A. R. Lectin and peritrophic membrane development in the gut of Glossina m.morsitans and a discussion of their role in protecting the fly against trypanosome infection. Med. Vet. Entomol. 5, 495–501 (1991).
Lehane, M. J., Allingham, P. G. & Weglicki, P. Composition of the peritrophic matrix of the tsetse fly, Glossina morsitans morsitans. Cell Tissue Res. 283, 375–384 (1996).
Rose, C. et al. An investigation into the protein composition of the teneral Glossina morsitans morsitans peritrophic matrix. PLoS Negl. Trop. Med. 8, e2691 (2014).
Rogerson, E. et al. Variations in the peritrophic matrix composition of heparan sulphate from the tsetse fly, Glossina morsitans morsitans. Pathogens 7, e7010032 (2018).
Taylor, A. W. The development of West African strains of Trypanosoma gambiense in Glossina tachinoides under normal laboratory. Parasitology 24, 401–409 (1932).
Yorke, W., Murgatroyd, F. & Hawking, F. The relation of polymorphic trypanosomes, developing in the gut of Glossina, to the peritrophic membrane. Ann. Trop. Med. Parasit. 27, 347–354 (2016).
Freeman, J. C. The presence of trypanosomes in the ecto-peritrophic space of tsetse flies, half an hour after ingestion of the infective blood meal. Trans. R. Soc. Trop. Med. Hyg. 64, 187–188 (1970).
Freeman, J. C. The penetration of the peritrophic membrane of the tsetse flies by trypanosomes. Acta Tropica 30, 347–355 (1973).
Fairbairn, H. The penetration of Trypanosoma rhodesiense through the peritrophic membrane of Glossina palpalis. Ann. Trop. Med. Parasit. 52, 18–19 (1958).
Moloo, S. K., Steiger, R. F. & Hecker, H. Ultrastructure of the peritrophic membrane formation in Glossina Wiedemann. Acta Tropica 27, 378–383 (1970).
Gibson, W. & Bailey, M. The development of Trypanosoma brucei within the tsetse fly midgut observed using green fluorescent trypanosomes. Kinetoplastid Biol. Dis. 2, 1 (2003).
Berriman, M. et al. The genome of the African trypanosome Trypanosoma brucei. Science 309, 416–422 (2005).
Aksoy, E. et al. Mammalian African trypanosome VSG coat enhances tsetse’s vector competence. Proc. Natl Acad. Sci. USA 113, 6961–6966 (2016).
Weiss, B. L., Savage, A. F., Griffith, B. C., Wu, Y. & Aksoy, S. The peritrophic matrix mediates differential infection outcomes in the tsetse fly gut following challenge with commensal, pathogenic, and parasitic microbes. J. Immunol. 193, 773–782 (2014).
Schuster, S. et al. Developmental adaptations of trypanosome motility to the tsetse fly host environments unravel a multifaceted in vivo microswimmer system. eLife 6, e27656 (2017).
Robertson, M. V. Notes on the life-history of Trypanosoma gambiense, with a brief reference to the cycles of Trypanosoma nanum and Trypanosoma pecorum in Glossina palpalis. Phil. Trans. R. Soc. Lond. B 203, 161–184 (1913).
Sharma, R. et al. Asymmetric cell division as a route to reduction in cell length and change in cell morphology in trypanosomes. Protist 159, 137–151 (2008).
Butikofer, P. et al. Phosphorylation of a major GPI-anchored surface protein of Trypanosoma brucei during transport to the plasma membrane. J. Cell Sci. 112, 1785–1795 (1999).
Toh, H. et al. Massive genome erosion and functional adaptations provide insights into the symbiotic lifestyle of Sodalis glossinidius in the tsetse host. Genome Res. 16, 149–156 (2006).
Escott, G. M. & Adams, D. J. Chitinase activity in human serum and leukocytes. Infect. Immun. 63, 4770–4773 (1995).
Rogers, M. E. et al. Leishmania chitinase facilitates colonization of sand fly vectors and enhances transmission to mice. Cell Microbiol. 10, 1363–1372 (2008).
Langer, R. C. & Vinetz, J. M. Plasmodium ookinete-secreted chitinase and parasite penetration of the mosquito peritrophic matrix. Trends Parasitol. 17, 269–272 (2001).
Peters, W. Peritrophic Membranes (eds Bradshaw, S. D. et al.) 123–148 (Springer, 1992).
Merzendorfer, H., Kelkenberg, M. & Muthukrishnan, S. in Extracellular Composite Matrices in Arthropods (eds Cohen, E. & Moussian, B.) 255–234 (Springer, 2016).
Vigneron, A. et al. A fine-tuned vector-parasite dialogue in tsetse’s cardia determines peritrophic matrix integrity and trypanosome transmission success. PLoS Pathog. 14, e1006972 (2018).
Ridgley, E. L., Xiong, Z. H. & Ruben, L. Reactive oxygen species activate a Ca2+-dependent cell death pathway in the unicellular organism Trypanosoma brucei brucei. Biochem. J. 340, 33–40 (1999).
Ooi, C.-P., Haines, L. R., Southern, D. M., Lehane, M. J. & Acosta-Serrano, A. Tsetse GmmSRPN10 has anti-complement activity and is important for successful establishment of trypanosome infections in the fly midgut. PLoS Negl. Trop. Dis. 9, e3448 (2015).
Haines, L. R. Examining the tsetse teneral phenomenon and permissiveness to trypanosome infection. Front. Cell Infect. Microbiol. 3, 84 (2013).
Beschin, A., Van Den Abbeele, J., De Baetselier, P. & Pays, E. African trypanosome control in the insect vector and mammalian host. Trends Parasitol. 30, 538–547 (2014).
Oberholzer, M., Lopez, M. A., McLelland, B. T. & Hill, K. L. Social motility in African trypanosomes. PLoS Pathog. 6, e1000739 (2010).
Imhof, S., Vu, X. L., Bütikofer, P. & Roditi, I. A glycosylation mutant of Trypanosoma brucei links social motility defects in vitro to impaired colonization of tsetse flies in vivo. Eukaryot. Cell 14, 588–592 (2015).
Lopez, M. A., Saada, E. A. & Hill, K. L. Insect stage-specific adenylate cyclases regulate social motility in African trypanosomes. Eukaryot. Cell 14, 104–112 (2015).
Shaw, S. et al. Flagellar cAMP signaling controls trypanosome progression through host tissues. Nat. Commun. 10, 803 (2019).
Imhof, S., Knüsel, S., Gunasekera, K., Vu, X. L. & Roditi, I. Social motility of African trypanosomes is a property of a distinct life-cycle stage that occurs early in tsetse fly transmission. PLoS Pathog. 10, e1004493 (2014).
Mehlitz, D., Zillmann, U., Scott, C. M. & Godfrey, D. G. Epidemiological studies on the animal reservoir of Gambiense sleeping sickness. Part III. Characterization of trypanozoon stocks by isoenzymes and sensitivity to human serum. Tropenmed. Parasitol. 33, 113–118 (1982).
Bingle, L. E., Eastlake, J. L., Bailey, M. & Gibson, W. C. A novel GFP approach for the analysis of genetic exchange in trypanosomes allowing the in situ detection of mating events. Microbiology 147, 3231–3240 (2001).
MacGregor, P., Rojas, F., Dean, S. & Matthews, K. R. Stable transformation of pleomorphic bloodstream form Trypanosoma brucei. Mol. Biochem. Parasitol. 190, 60–62 (2013).
Zimmermann, H. et al. A quorum sensing-independent path to stumpy development in Trypanosoma brucei. PLoS Pathog. 13, e1006324 (2017).
Schuster, S. et al. A modification to the life cycle of the parasite Trypanosoma brucei. Preprint at bioRxiv https://doi.org/10.1101/717975 (2019).
Peacock, L., Ferris, V., Bailey, M. & Gibson, W. Multiple effects of the lectin-inhibitory sugars d-glucosamine and N-acetyl-glucosamine on tsetse-trypanosome interactions. Parasitology 132, 651–658 (2006).
Ziegelbauer, K., Quinten, M., Schwarz, H., Pearson, T. W. & Overath, P. Synchronous differentiation of Trypanosoma brucei from bloodstream to procyclic forms in vitro. Eur. J. Biochem. 192, 373–378 (1990).
Deerinck, T. J., Bushong, E. A., Thor, A. & Ellisman, M. H. (eds) NCMIR methods for 3D EM: a new protocol for preparation of biological specimens for serial block face scanning electron microscopy. (National Center for Microscropy and Imaging Research, 2010).
Schneider, C. A., Rasband, W. S. & Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 9, 671–675 (2012).
Acknowledgements
We thank T. Pearson (University of Victoria, Canada) for providing us with anti-GPEET mouse hybridomas; S. Vaughan (Oxford Brookes University) for making available essential TEM protocols; A. Schnaufer for providing us with T. brucei AnTat 1.1 90:13 stabilates and W. Gibson for supplying the T. brucei J10 strain; L. Jeacock for artwork and members of the Acosta Serrano group for discussions. This research was supported by Wellcome Trust project grant 093691/Z/10/Z (awarded to A.A.-S.), Multi-User Equipment grant (for CLSM images 104936/Z/14/Z), MRC Concept in Confidence award MC_PC_17167, GlycoPar-EU FP7 Marie Curie Initial Training Network (no. 608295, to A.C.S. and A.A.-S.) and a PhD studentship from LSTM (to C.R.). J.L. was supported by the Deutscher Akademischer Austauschdienst (DAAD) and M.E. was supported by Deutsche Forschungsgemeinschaft (DFG) grants EN305, SPP1726 (Microswimmers–From Single Particle Motion to Collective Behaviour) and GRK2157 (3D Tissue Models to Study Microbial Infections by Obligate Human Pathogens). M.E. is board member of the Wilhelm Conrad Roentgen Center for Complex Material Systems (RCCM).
Author information
Authors and Affiliations
Contributions
C.R., A.C.-S., N.A.D., L.R.H., M.J.L. and A.A.-S. conceived and designed experiments. C.R., N.A.D., A.J.B., B.M., C.S., L.R.H., M.J.L. and I.A.P. conducted and analysed EM research. A.C.-S., C.R., N.A.D. and M.M. obtained CLSM data. J.L. and M.E. obtained and analysed SEM images. C.R. and A.A.-S. wrote the paper with input from all of the authors.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 TEM analysis of the tsetse PM.
a, Micrographs showing the PM from either eleven days old naïve (left), eleven dpi refractory (middle) or 5 days old naïve (right) flies. All 3 PM layers can be clearly seen; secretions presumably coming from the epithelial cells (*) appear to stick onto PM2 and PM3. Along the length of the PM, PM3 is sometimes observed as a discontinued electron-dense line (arrowheads). The PM within the context of the midgut is highly convoluted in an unfed/starved fly (right). BI, basal infoldings; M, mitochondria; MV, microvilli; N, nucleus; CJ, cell junction; ES, ectoperitrophic space; L, lumen. Scale bars, from left to right, 2 µm, 1 µm, 5 µm. b, Tsetse PM width remains constant throughout infection. Violin plot showing the distribution in width of the PM in naïve compared to infected flies, at either five or eleven dpi. Line represents the mean value, dashed lines indicate both quartiles, circles represent individual measurements, and numbers under each plot indicate the sample size of each group (n=1 experiment). Horizontal lines and p-values indicate statistical significance (unpaired two-tailed t-test, normal distribution and equal variance). Measurements were taken between PM1 and PM3.
Extended Data Fig. 2 CLSM reveals trypanosomes trapped in the PM.
a, Washed, ex vivo PM from a naïve fly (eleven days old) stained with WGA-Rhodamine (top). In eleven dpi flies (middle), eGFP-trypanosomes can be seen (green) in close proximity to the PM, with DAPI (magenta) showing parasite nuclei and kinetoplasts. Scale bar 200 µm. Inset corresponds to the higher magnification of the same area as seen in bottom panel with a 20 µm scale (see Supplementary Video 1). DIC, Differential Interference Contrast. b, Maximum Intensity Projection of a series of z-stacks, and corresponding orthogonal views (top), and 3D reconstruction (bottom) of a trypanosome trapped within the PM (see also Supplementary Video 2). Scale bars 20 µm. n=35 experiments, 10 flies per timepoint.
Extended Data Fig. 3 TEM analysis and SBF-SEM 3D reconstruction of a trypanosome cyst-like body from the anterior midgut.
a, Typical damage found in infected flies is the separation of PM1 from PM2. Usually, the electron dense (PM1) layer appears unbroken, but can be seen peeling away from the second layer (arrowheads). Note that PM1 remains unbroken even when cyst-like bodies contain high parasite numbers. Scale bars left to right 2 µm, 2 µm, 1 µm and 2 µm. ES, ectoperitrophic space. L, lumen. EC, epithelial cell. Number of samples used for TEM analysis see Supplementary Fig. 4. b, SBF-SEM reconstruction of a cyst-like body at eleven dpi (see Fig. 1c, and Supplementary Videos 3 and 4). Slices merged with manual segmentation show breaks or damage to PM1 are absent during a trypanosome infection (top left); trypanosomes are contained within PM1 (top right) and ES-side parasites do not interact with the cyst-like body (bottom left). Bottom right panel shows the length of a partially reconstructed trypanosome within the same cyst-like body. Scale bars 4 µm are representative of the SEM image (not the reconstruction). Reconstruction (n=1).
Extended Data Fig. 4 Analyses of early proventricular infection and formation of cyst-like bodies.
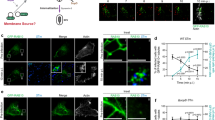
a, Time course of infection up to five dpi with eGFP BSFs (J10 strain) using CLSM. Figure shows representative proventriculi and anterior midguts from each dpi. “Day 0”, proventriculus from a fly dissected 1 hr after receiving an infected meal. Black arrowhead at two dpi shows trypanosomes already in the proventriculus (see Supplementary Video 8). Scale bars 200 µm. b, TEM analysis of an early (five dpi) proventricular invasion by trypanosomes. Trypanosomes (T) can be seen near to the area of cell transition between the foregut (FG) and the proventricular/midgut epithelial cells (EC). At this time point the PV is heavily infected and parasites are located in the lumen (L), the ES and between PM layers. CI, cuticula intima. Scale bars left to right 2 µm, 2 µm and 10 µm. c, SEM analysis of an early (4 dpi) proventricular invasion by trypanosomes. (1), the tsetse PM is secreted by specialised cells near to the dorsal (Dors) side of the proventriculus (scale bar 100 µm). Trypanosomes can be seen within the immature PM (2; scale bar 10 µm) and also in the ectoperitrophic space (3, 4) after the PM has been pressed and hardened. Arrows on a dotted line represent PM path to the MG. Scale bar 10 µm and 1 µm respectively. Ant, anterior. Vent, ventral. FB, fat bodies. MG, midgut. d, TEM analysis of infected proventriculi from flies at eleven dpi. Parasites are contained within PM1 and PM2 or in the ES. No parasites can be seen within the lumen at this time point and the formation of cyst-like bodies can be observed. MV, microvilli. Scale bars left to right 2 µm, 5 µm and 20 µm. CLSM n=2 experiments, 10 flies dissected per timepoint.
Extended Data Fig. 5 Gradual loss of early proventricular infection by cultured bloodstream form (cBSF) trypanosomes.
a, Flies that had received a bloodmeal containing J10 BSFs (from murine stabilate) were dissected each day and midgut (MG) and proventriculus (PV) infections were scored before preparation for CLSM. Trypanosomes were observed in the PV from two dpi. Two experiments, n=19-22 flies per timepoint. b, Flies were given an infected bloodmeal with either BSFs (Blood), cBSFs that had been adapted to culture from the same BSF stabilate for 9 days (Adapted) and cBSFs that had been in culture for several months (Cultured). Flies were dissected at five dpi and fifteen dpi and infection prevalence was scored for both the proventriculus (PV) and the midgut (MG), or 30 dpi to check salivary gland (SG) infection. Two experiments, n=28-33 flies each timepoint. c, Retrospective analyses of salivary gland (SG) infections using either bloodstream forms (BSFs), cultured procyclic forms (PCFs) or cultured BSFs (cBSFs) for infection. Each point represents an independent experiment (with >100 flies per experiment). Life stages used for all experiments were from AnTat 1.1 90:13 strain. Mean ± SD, numbers on the plots indicate the sample size of each group, horizontal bars and p-values show statistical significance from an unpaired two-tailed t-test, normal distribution and equal variance. d, Flies were given an infected meal with cBSF T. brucei (strain AnTat 1.1 90:13 expressing mNeonGreen) and dissected every day until trypanosomes were observed in the proventriculus. In contrast to BSFs from a murine stabilate, the cultured forms do not invade the proventriculus until at least 15 dpi. The distribution is also different as very few trypanosomes can be seen in the ectoperitrophic space in the anterior midgut. Scale bars 100 µm.
Extended Data Fig. 6 Artistic representation of how trypanosomes enter the ectoperitrophic space via the proventriculus.
Specialised epithelial cells in the proventricular annular pad are responsible for PM (orange) assembly and secretion. Ingested slender and stumpy bloodstream trypanosomes (blue) transform into procyclic forms (green) within the proventricular lumen (1) and then successfully migrate to the ES through a more fluid PM in the proventriculus (2) before it maturates into a fully assembled structure as seen in the midgut. Alternatively, they may become trapped between PM layers forming cyst-like bodies (3). Parasites trapped between PM layers are carried (as on a conveyor belt) through the midgut as the PM continues to be secreted (4). OE and C represent direction of the blood flow from either the oesophagus or crop, respectively.
Extended Data Fig. 7 A selection of TEM micrographs taken from the hindgut of a fly at 11 dpi.
a, An overview of the hindgut of an infected fly. The epithelial cells of the hindgut are distinctive (HGE) and the PM can be seen. In this region, material presumably from degraded and discarded PM can be seen (*). Scale bar 20 µm. b–d, Empty cyst-like body with no visible parasites (*). Scale bars 10 µm, 10 µm and 5 µm, respectively. e, f, Trypanosomes with abnormal morphology in the ectoperitrophic space of the hindgut. Arrowheads indicate the presence of parasites with multiple flagella. Scale bars 5 µm and 10 µm, respectively. For TEM analysis refer to Supplementary Fig. 4.
Extended Data Fig. 8 Trypanosomes are unable to colonise the PV and produce lower MG infections in previously blood fed flies.
a, Representative microscopy images of proventriculi from teneral and refractory tsetse (i.e. blood fed three times before receiving a trypanosome-infected bloodmeal) at three dpi in bright field, a maximum intensity projection of three confocal z-stacks of eGFP-expressing T. brucei strain J-10 (green) and merge. Scale bar 200 μm. b, Infection prevalence rates (percentage) of proventriculi (PV) and midguts (MG) from teneral (green) and refractory (purple) flies at three dpi and five dpi. Sample size is shown as numbers above bars, one experiment.
Supplementary information
Supplementary Information
Supplementary Figs. 1–4.
Supplementary Video 1
The z-stack of trypanosome cyst-like body. GFP-expressing trypanosomes can be observed within the layers of an isolated PM from a fly at 9 d.p.i; 79 stacks of 0.5 µm. Parasite DNA is shown in magenta. Taken under ×63 oil-immersion objective.
Supplementary Video 2
CLSM of embedded trypanosomes. Maximum intensity projection of an isolated PM (orange) from a fly at 8 d.p.i. Trypanosomes (green) can be seen trapped on the ectoperitrophic side of the PM by PM material.
Supplementary Video 3
Serial sections of a cyst-like body from a fly at 11 d.p.i.; 474 serial sections of a trypanosome-filled pocket in the PM in the anterior midgut. Sections were cut at 100 nm and scanned under SBF-SEM.
Supplementary Video 4
Reconstruction of a cyst-like body from a fly at 11 d.p.i.; SBF-SEM and manual segmentation showing parasites are contained and lying parallel within PM1 and PM2 and are not seen crossing the PM.
Supplementary Video 5
Serial sections of PV ROI1 from a fly at 5 d.p.i.; 458 serial sections in a region of high parasitaemia in the PV of a fly at 5 d.p.i. Sections were cut at 40 nm and scanned under SBF-SEM.
Supplementary Video 6
Serial sections of PV ROI2 from a fly at 5 d.p.i.; 457 serial sections in a second region of high parasitaemia in the PV of a fly at 5 d.p.i. Sections were cut at 40 nm and scanned under SBF-SEM.
Supplementary Video 7
Reconstruction of PV ROI2 from a fly at 5 d.p.i. SBF-SEM and manual segmentation showing that the cuticular intima (yellow) of the foregut is intact; there appears to be no penetration of PV cells by trypanosomes and no indication of attachment. Four trypanosomes were also reconstructed. The scale bar is representative of the SEM image and not the reconstructed image.
Supplementary Video 8
PV from a fly at 2 d.p.i. PV shown in differential interference contrast with trypanosomes in green, indicated by the line and circles. Trypanosomes are clearly inside the tissue and are different from those swimming freely outside.
Supplementary Video 9
Annotated CLSM of naive PV. The z-stack of a naive PV showing the origin of the PM (orange), F-actin structure of the PV (white) and nuclei (magenta).
Supplementary Video 10
3D representation of naive PV. 3D render of a PV from a naive fly (as shown in Supplementary Video 9).
Supplementary Video 11
Annotated CLSM of infected PV. The z-stack of a PV from an infected fly at 5 d.p.i. The stains are the same as described in Supplementary Video 9 but expressing eGFP to show trypanosome distribution.
Supplementary Video 12
3D representation of infected PV. 3D render of a PV from a fly at 5 d.p.i. (as shown in Supplementary Video 11).
Supplementary Data
Statistical source data.
Source data
Source Data Fig. 3
Statistical source data.
Source Data Fig. 4
Statistical source data.
Source Data Extended Data Fig. 1b
Statistical source data.
Source Data Extended Data Fig. 5
Statistical source data.
Source Data Extended Data Fig. 8
Statistical source data.
Rights and permissions
About this article

Cite this article
Rose, C., Casas-Sánchez, A., Dyer, N.A. et al. Trypanosoma brucei colonizes the tsetse gut via an immature peritrophic matrix in the proventriculus. Nat Microbiol 5, 909–916 (2020). https://doi.org/10.1038/s41564-020-0707-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41564-020-0707-z
This article is cited by
-
Minimal resin embedding of SBF-SEM samples reduces charging and facilitates finding a surface-linked region of interest
Frontiers in Zoology (2023)
-
Cyclic AMP signalling and glucose metabolism mediate pH taxis by African trypanosomes
Nature Communications (2022)
-
A multi-adenylate cyclase regulator at the flagellar tip controls African trypanosome transmission
Nature Communications (2022)
-
The holobiont transcriptome of teneral tsetse fly species of varying vector competence
BMC Genomics (2021)
