
Abstract
Leveraging the motion and force of individual molecular motors in a controlled manner to perform macroscopic tasks can provide substantial benefits to many applications, including robotics. Nonetheless, although millimetre-scale movement has been demonstrated with synthetic and biological molecular motors, their efficient integration into engineered systems that perform macroscopic tasks remains challenging. Here, we describe an active network capable of macroscopic actuation that is hierarchically assembled from an engineered kinesin, a biomolecular motor, and microtubules, resembling the contractile units in muscles. These contracting materials can be formed in desired areas using patterned ultraviolet illumination, allowing their incorporation into mechanically engineered systems, being also compatible with printing technologies. Due to the designed filamentous assembly of kinesins, the generated forces reach the micronewton range, enabling actuation of millimetre-scale mechanical components. These properties may be useful for the fabrication of soft robotic systems with advanced functionalities.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$259.00 per year
only $21.58 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout





Similar content being viewed by others


Data availability
Plasmids are available from the corresponding author upon request. Source data are provided with this paper.
Code availability
The codes used to simulate the printable active network are available at the following Github repository: https://github.com/takanitta/PrintableActiveNetwork.
References
Yang, G.-Z. et al. The grand challenges of Science Robotics. Sci. Robot. 3, eaar7650 (2018).
Li, Q. et al. Macroscopic contraction of a gel induced by the integrated motion of light-driven molecular motors. Nat. Nanotechnol. 10, 161–165 (2015).
Chen, J. et al. Artificial muscle-like function from hierarchical supramolecular assembly of photoresponsive molecular motors. Nat. Chem. 10, 132–138 (2018).
Vale, R. D. & Milligan, R. A. The way things move: looking under the hood of molecular motor proteins. Science 288, 88–95 (2000).
Goel, A. & Vogel, V. Harnessing biological motors to engineer systems for nanoscale transport and assembly. Nat. Nanotechnol. 3, 465–475 (2008).
Korten, T., Månsson, A. & Diez, S. Towards the application of cytoskeletal motor proteins in molecular detection and diagnostic devices. Curr. Opin. Biotechnol. 21, 477–488 (2010).
Saper, G. & Hess, H. Synthetic systems powered by biological molecular motors. Chem. Rev. 120, 288–309 (2020).
Lin, C. T., Kao, M. T., Kurabayashi, K. & Meyhofer, E. Self-contained, biomolecular motor-driven protein sorting and concentrating in an ultrasensitive microfluidic chip. Nano Lett. 8, 1041–1046 (2008).
Fischer, T., Agarwal, A. & Hess, H. A smart dust biosensor powered by kinesin motors. Nat. Nanotechnol. 4, 162–166 (2009).
Lard, M. et al. Ultrafast molecular motor driven nanoseparation and biosensing. Biosens. Bioelectron. 48, 145–152 (2013).
Aoyama, S., Shimoike, M. & Hiratsuka, Y. Self-organized optical device driven by motor proteins. Proc. Natl Acad. Sci. USA 110, 16408–16413 (2013).
Nicolau, D. V. et al. Parallel computation with molecular-motor-propelled agents in nanofabricated networks. Proc. Natl Acad. Sci. USA 113, 2591–2596 (2016).
Sato, Y., Hiratsuka, Y., Kawamata, I., Murata, S. & Nomura, S. M. Micrometer-sized molecular robot changes its shape in response to signal molecules. Sci. Robot. 2, eaal3735 (2017).
Needleman, D. & Dogic, Z. Active matter at the interface between materials science and cell biology. Nat. Rev. Mater. 2, 17048 (2017).
Koenderink, G. H. & Paluch, E. K. Architecture shapes contractility in actomyosin networks. Curr. Opin. Cell Biol. 50, 79–85 (2018).
Nedelec, F. J., Surrey, T., Maggs, A. C. & Leibler, S. Self-organization of microtubules and motors. Nature 389, 305–308 (1997).
Köhler, S., Schaller, V. & Bausch, A. R. Structure formation in active networks. Nat. Mater. 10, 462–468 (2011).
Wollman, A. J. M., Sanchez-Cano, C., Carstairs, H. M. J., Cross, R. A. & Turberfield, A. J. Transport and self-organization across different length scales powered by motor proteins and programmed by DNA. Nat. Nanotechnol. 9, 44–47 (2014).
Foster, P. J., Fürthauer, S., Shelley, M. J. & Needleman, D. J. Active contraction of microtubule networks. eLife 4, e10837 (2015).
Torisawa, T., Taniguchi, D., Ishihara, S. & Oiwa, K. Spontaneous formation of a globally connected contractile network in a microtubule-motor system. Biophys. J. 111, 373–385 (2016).
Sanchez, T., Welch, D., Nicastro, D. & Dogic, Z. Cilia-like beating of active microtubule bundles. Science 333, 456–459 (2011).
Rich, S. I., Wood, R. J. & Majidi, C. Untethered soft robotics. Nat. Electron. 1, 102–112 (2018).
Linsmeier, I. et al. Disordered actomyosin networks are sufficient to produce cooperative and telescopic contractility. Nat. Commun. 7, 12615 (2016).
Schuppler, M., Keber, F. C., Kröger, M. & Bausch, A. R. Boundaries steer the contraction of active gels. Nat. Commun. 7, 13120 (2016).
Ross, T. D. et al. Controlling organization and forces in active matter through optically defined boundaries. Nature 572, 224–229 (2019).
Alberts, B. et al. Molecular Biology of the Cell (Norton, 2014).
Svoboda, K. & Block, S. M. Force and velocity measured for single kinesin molecules. Cell 77, 773–784 (1994).
Bagshaw, C. R. Muscle Contraction (Kluwer Academic Publishers, 1993).
Murphy, S. V. & Atala, A. 3D bioprinting of tissues and organs. Nat. Biotechnol. 32, 773–785 (2014).
Alvarado, J., Sheinman, M., Sharma, A., MacKintosh, F. C. & Koenderink, G. H. Force percolation of contractile active gels. Soft Matter 13, 5624–5644 (2017).
Murrell, M., Oakes, P. W., Lenz, M. & Gardel, M. L. Forcing cells into shape: the mechanics of actomyosin contractility. Nat. Rev. Mol. Cell Biol. 16, 486–498 (2015).
Day, R. N. & Davidson, M. W. The fluorescent protein palette: tools for cellular imaging. Chem. Soc. Rev. 38, 2887–2921 (2009).
Dasanayake, N. L. & Carlsson, A. E. Stress generation by myosin minifilaments in actin bundles. Phys. Biol. 10, 036006 (2013).
Lenz, M. Geometrical origins of contractility in disordered actomyosin networks. Phys. Rev. X 4, 041002 (2014).
Nakamura, M. et al. Remote control of myosin and kinesin motors using light-activated gearshifting. Nat. Nanotechnol. 9, 693–697 (2014).
Matsuda, T., Kawakami, R., Namba, R., Nakajima, T. & Gong, J. P. Mechanoresponsive self-growing hydrogels inspired by muscle training. Science 363, 504–508 (2019).
Abramson, A. et al. An ingestible self-orienting system for oral delivery of macromolecules. Science 363, 611–615 (2019).
Iwamoto, H. & Yagi, N. The molecular trigger for high-speed wing beats in a bee. Science 341, 1243–1246 (2013).
Hyman, A. A. Preparation of marked microtubules for the assay of the polarity of microtubule-based motors by fluorescence. J. Cell Sci. Suppl. 14, 125–127 (1991).
Blumenthal, D. K. et al. Identification of the calmodulin-binding domain of skeletal muscle myosin light chain kinase. Proc. Natl Acad. Sci. USA 82, 3187–3191 (1985).
Beckett, D., Kovaleva, E. & Schatz, P. J. A minimal peptide substrate in biotin holoenzyme synthetase-catalyzed biotinylation. Protein Sci. 8, 921–929 (1999).
Castoldi, M. & Popov, A. V. Purification of brain tubulin through two cycles of polymerization-depolymerization in a high-molarity buffer. Protein Expr. Purif. 32, 83–88 (2003).
Hyman, A. et al. Preparation of modified tubulins. Methods Enzymol. 196, 478–485 (1991).
Margossian, S. S. & Lowey, S. Preparation of myosin and its subfragments from rabbit skeletal muscle. Methods Enzymol. 85, 55–71 (1982).
Hess, H., Clemmens, J., Qin, D., Howard, J. & Vogel, V. Light-controlled molecular shuttles made from motor proteins carrying cargo on engineered surfaces. Nano Lett. 1, 235–239 (2001).
Hiratsuka, Y., Miyata, M. & Uyeda, T. Q. P. Living microtransporter by uni-directional gliding of mycoplasma along microtracks. Biochem. Biophys. Res. Commun. 331, 318–324 (2005).
Steinbock, O., Tóth, A. & Showalter, K. Navigating complex labyrinths: optimal paths from chemical waves. Science 267, 868–871 (1995).
Nakagaki, T., Yamada, H. & Tóth, Á. Maze-solving by an amoeboid organism. Nature 407, 470 (2000).
Tan, J. L. et al. Cells lying on a bed of microneedles: an approach to isolate mechanical force. Proc. Natl Acad. Sci. USA 100, 1484–1489 (2003).
Landau, L. D. & Lifshitz, E. M. Theory of Elasticity (Butterworth-Heinemann, 1986).
Rupp, B. & Nédélec, F. Patterns of molecular motors that guide and sort filaments. Lab Chip 12, 4903–4910 (2012).
Acknowledgements
We thank C. Tatsumi for technical assistance and K. Hirai for carrying out preliminary experiments. We thank H. Hess at Columbia University, Y. Inoue at Kyoto University and T. Q. P. Uyeda at Waseda University for their comments and discussions. This work was supported by the New Energy and Industrial Technology Development Organization (NEDO, No. 16100859-0), MEXT (KAKENHI ‘Molecular Robotics’ JP24104004) and JSPS (KAKENHI JP18H01407, to Y.H.).
Author information
Authors and Affiliations
Contributions
Y.H. and T.N. conceived the experimental work. Y.H. led the experiments. T.N. performed the simulations. Z.D. contributed to the force measurement experiments. Y.W. and K.M. contributed to the TEM measurements and part of the microfabrication. T.N. and Y.H. contributed to the data analysis and interpretation, and wrote the paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Peer review information Nature Materials thanks the anonymous reviewers for their contribution to the peer review of this work.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
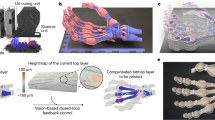
Extended Data Fig. 1 Molecular design of the printable active network.
a, Myosin in muscle sarcomeres. Myosin II has two functional domains, heavy meromyosin (HMM) and light meromyosin (LMM). The main role of HMM is to propel actin filaments, whereas the LMM domain promotes myosin filament formation. In a muscle sarcomere, myosin II forms bipolar filaments and assembles them into a highly-ordered interlocked structure with actin filaments and other proteins. Sarcomeres contract through sliding of the actin filaments caused by myosin conformational changes. b, Design of fusion proteins for the printable active network. CaMLMM is a genetically engineered fusion protein of calmodulin (CaM) and LMM. This fusion protein assembles into filaments and serves as a calcium-activated scaffold for proteins possessing a calmodulin binding sequence. K465m13 is a kinesin I truncation (aa 1–465) with the m13 CaM binding sequence of the myosin light chain kinase (MLCK) at the C-terminus. c, Filament formation of CaMLMM. CaMLMM forms filaments similar to myosin filaments in low-salt conditions. d, Assembly of kinesin filaments triggered by UV irradiation. UV irradiation causes an influx of calcium ions released from caged molecules and results in the decoration of CaMLMM filaments by K465m13 fusion protein units. e, Contraction. Kinesin filaments induce the sliding of microtubules and further dynamic self-assembly into a contractile network.
Extended Data Fig. 2 Biochemical and electron microscopy confirmation of the formation of kinesin filaments.
a-b, Biochemical confirmation of CaMLMM filament formation. The solubility of CaMLMM and myosin II in various NaCl concentrations was measured by sedimentation assays and SDS-PAGE. Representative images of n = 2. Uncropped images with molecular weight marker are provided as Source Data. Both CaMLMM and myosin II were found in soluble fractions only at high NaCl concentrations (200 mM or more). Since myosin II is known to form filaments and precipitate in low ionic strength conditions, this result indicates that CaMLMM behaves similarly to myosin. c, Biochemical confirmation of the binding of K465m13 to the CaMLMM filament under the presence of calcium ions. Co-sedimentation of K465m13 was analysed by SDS-PAGE. In the presence of calcium ions, 77% of K465m13 co-sediments with CaMLMM, while most of the K465m13 is found in the supernatant in samples containing EGTA, a calcium chelating agent. This result indicates that K465m13 is bound to CaMLMM in the presence of calcium as designed. d, TEM image of CaMLMM and kinesin filaments (also see Supplementary Discussion 1). Filamentous objects possessing lengths around 0.8 µm, comparable with synthetic myosin filaments, were observed (upper and lower panels). In K465m13-positive samples, the filamentous objects became slightly thicker than the negative control (lower). Close examination of the CaMLMM filaments with K465m13 revealed globular objects attached to the filaments at intervals of around 40 nm (inset), close to that of myosin heads along myosin filaments28, indicating that the arrangement of the K465m13 fusion protein on the kinesin filaments is similar to that of myosin heads in myosin filaments. Representative images of n = 3. Scale bar: 500 nm.
Extended Data Fig. 3 Contraction force of printable active network in experiment and simulation.
a, Time evolutions of bending force of the flexible cantilever under various concentrations of K465m13. After reaching maxima, in many cases, the displacement was suddenly dropped due to rupture of the active network. b, Maximum contraction force produced vs. K465m13 concentration, tubulin concentration, and mean microtubule length. c, Contraction force steadily increased with time until the active network ruptured, which is in agreement with the experimental results shown in a. d, The maximum force increased with (left) motor density; (middle) the number of nodes; and (right) node radius. These results agree with the experimental results in b, assuming that K465m13 concentration, tubulin concentration, and microtubule length correspond to motor density, the number of nodes, and the node radius, respectively. In the node radius simulation shown in (right), the product of the number of the nodes and the radius of individual nodes was kept constant to be consistent with the experimental condition of a constant tubulin concentration. Data points are represented as mean ± s.d. in b (n = 3) and d (n = 5).
Extended Data Fig. 4 Phase diagrams for contraction with various microtubule lengths (LMT).
a. CaMLMM. b. CaMCFP. The critical tubulin concentration required for contraction to occur (red dashed lines) decreases as microtubule length increases. Comparison between CaMLMM and CaMCFP revealed that to induce contraction more tubulin and K465m13 are required with CaMCFP than with CaMLMM. The diameters of chambers are 1 mm. For the length of microtubule, see Supplementary Fig. 7.
Extended Data Fig. 5 Circular network contractions in experiment and simulation.
a-c, Replotted from Fig. 4e and f in the main text. a, Time evolutions of the radii of networks with different initial radii. b, The contraction speed was a linear function of the radius of the contracting networks. c, The contraction rate was inversely proportional to the microtubule length LMT. Marks represent K465m13 concentrations (open diamond: 0.5 µM, triangle: 1.0 µM, square: 2.0 µM and circle: 4.0 µM, respectively). d, Time evolutions of the radii of simulated networks with different initial radii. e, The contraction speed was a linear function of the radii of contracting networks, reproducing b except the acceleration phase at the large radius. f, The contraction rate was ~vkin/Rnode. Assuming that Rnode is comparable with microtubule length, the simulation result is consistent with the experimental one. Data points are represented as mean ± s.d. (n = 5).
Supplementary information
Supplementary Information
Supplementary Discussions 1 and 2, Figs. 1–10, Videos (captions) 1–13 and references.
Supplementary Video 1
Contraction of the printable active network in a chamber. Fluorescence (TRITC-labelled microtubules) and bright-field images of the printable active network in a circular chamber with a diameter of 1.5 mm are shown. After 20 s, the sample was illuminated with 365-nm UV light to release the caged calcium ions, thus initiating the contraction (around 00:02 in the video). The fluorescent and bright-field channels were concatenated into one video. Video speed: ×10.
Supplementary Video 2
Contraction triggered by spatially selective UV irradiation. Upon illumination through the photomasks, the printable active networks assembled in a star or smiley shape and contracted while maintaining their shapes. Two videos for the star and smiley patterns were concatenated into one video. Video speed: ×10.
Supplementary Video 3
Contraction force measurement by the PDMS cantilever. The contraction force of the printable active network was measured by measuring the bending of an elastic PDMS cantilever. The PDMS cantilever (1,000 × 30 × 200 µm3) and an anchor were set in the middle of a chamber (700 µm in depth) using a spacer. The printable active network was generated between the cantilever and the anchor by irradiating a square UV pattern (500 × 3,000 µm2). In this video, the tip of the cantilever moved approximately 600 µm toward the anchor. Video speed: ×10.
Supplementary Video 4
Microgripper closing by the printable active network actuator. A PDMS microgripper consisting of a tip, an arm and two hinges totalling 1.5 mm in size was fabricated. By activation of the printable active network around the arm region, the hinges were bent, and the tip of the gripper closed within 30 s of activation. Fluorescent and bright-field channels were merged into one video. Video speed: ×10.
Supplementary Video 5
Robotic arm bending by the printable active network actuator. Our robotic arm consisted of three elements: two anchors tethering the active network to the body of the robot, a joint and a PDMS frame. Following activation of the printable active network between the anchors, the robotic arm bent within 100 s. Fluorescent and bright-field channels were merged into one video. Video speed: ×10.
Supplementary Video 6
Insect robot imitating walking by sequential activation of the printable active network actuator. The 2-mm insect-like robot was composed of three body segments with pairs of appendages connected by two joints. By activation around the left legs, the body of the robot was steered towards the left. After 90 s, rupture was caused by the restoring force of the two joints, and the main body returned to its original position. Next, the same activation was repeated on the right side. Fluorescent and bright-field channels were merged into one video. Video speed: ×10.
Supplementary Video 7
Micromanipulation and assembly of cogwheels by the printable active network actuator. To demonstrate the manipulation and assembly of micro-objects by the printable active network actuator, cogwheel structures with dimensions in the range 0.4–0.5 mm were used. By generating a ring-like active network around the two cogwheels, the cogwheels were brought into close proximity. Finally, the cogwheels meshed together. Fluorescent and bright-field channels were merged into one video. Video speed: ×10.
Supplementary Video 8
Accumulation of microbeads into a star shape. Microbeads with diameters of 15 µm (resin of RESOURCE Q, GE Healthcare) were dispersed in a solution of the printable active network. By activating the printable active network in a star-shaped area, the microbeads clustered in this area; 1 min after activation, approximately 300 microbeads had accumulated into a star-shaped structure. Video speed: ×10.
Supplementary Video 9
Demonstration of maze solving using the printable active network. Maze solving using a printable active network is demonstrated. The initial uniformly distributed active network contracted upon UV irradiation, leaving a connection between the start and goal anchors (around 00:12 in the video). Video speed: ×10.
Supplementary Video 10
Formation and coalescence of microtubule asters. In the non-contraction phase just below the critical concentrations, isolated asters of microtubules and coalescence between two asters were observed. The polarity-marked microtubule (minus ends: rhodamine) showed that the minus ends are pointing outwards. Two videos observed with ×20 and ×40 objective lenses were concatenated into one video. Scale bar: 50 µm. Video speed: ×50.
Supplementary Video 11
Simulation of the contraction of the printable active network in a circular chamber. Simulation of the contraction of the printable active network in a circular chamber with a radius of 15 µm. White dots represent node centres and blue lines represent the links connecting two nodes. Video speed: ×1.
Supplementary Video 12
Simulation of the contraction of the printable active network actuator between two anchors. A printable active network actuator with a random node array (upper), and one with a grid node array (lower). The densities of the nodes were 0.25 µm−2. The initial size of the printable active network was 100 × 30 µm2. Both ends, each with a width of 10 µm, were anchored. White dots represent the centres of nodes and the lines represent the links connecting two nodes. The magnitudes of the forces acting on the links are mapped in different colours, as shown in the colour bar. Video speed: ×1.
Supplementary Video 13
Simulation of the reversible contraction of the active network actuator. A printable active network actuator with a bidirectional motor protein. The direction of the motor protein was switched every 1 s. The initial size of the active network was 100 × 30 µm2. Both ends, each with a width of 10 µm, were anchored. The anchor on the right was connected to a spring. Magenta dots represent the centres of nodes and white lines represent the links connecting two nodes. Video speed: ×1.
Supplementary Data 1
Numerical data used to generate the graphs in the Supplementary figures.
Supplementary Data 2
DNA and amino acid sequences of the proteins used in this research.
Source data
Source Data Fig. 4
Numerical data used to generate graphs.
Source Data Fig. 5
Numerical data used to generate graphs.
Source Data Extended Data Fig. 2
Numerical data used to generate graphs and uncropped gel images.
Source Data Extended Data Fig. 3
Numerical data used to generate graphs.
Source Data Extended Data Fig. 5
Numerical data used to generate graphs.
Rights and permissions
About this article

Cite this article
Nitta, T., Wang, Y., Du, Z. et al. A printable active network actuator built from an engineered biomolecular motor. Nat. Mater. 20, 1149–1155 (2021). https://doi.org/10.1038/s41563-021-00969-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41563-021-00969-6
This article is cited by
-
Collective Molecular Machines: Multidimensionality and Reconfigurability
Nano-Micro Letters (2024)
-
Hierarchical supramolecular structure comprising reduction-responsive DNA microspheres and semi-artificial glycopeptide-based micro-asters
Polymer Journal (2023)
-
Competing instabilities reveal how to rationally design and control active crosslinked gels
Nature Communications (2022)
-
The rate of microtubule breaking increases exponentially with curvature
Scientific Reports (2022)
-
Linking path and filament persistence lengths of microtubules gliding over kinesin
Scientific Reports (2022)
