
Abstract
Little is known about the Pleistocene climatic context of northern Australia at the time of early human settlement. Here we generate a palaeoprecipitation proxy using stable carbon isotope analysis of modern and archaeological pandanus nutshell from Madjedbebe, Australia’s oldest known archaeological site. We document fluctuations in precipitation over the last 65,000 years and identify periods of lower precipitation during the penultimate and last glacial stages, Marine Isotope Stages 4 and 2. However, the lowest effective annual precipitation is recorded at the present time. Periods of lower precipitation, including the earliest phase of occupation, correspond with peaks in exotic stone raw materials and artefact discard at the site. This pattern is interpreted as suggesting increased group mobility and intensified use of the region during drier periods.
Similar content being viewed by others
Main
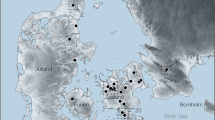
Madjedbebe is a large sandstone rockshelter located on Mirarr country in the Alligator Rivers region, northern Australia, with evidence for human occupation from at least 65 thousand years ago (ka) to the present (Fig. 1b)1. The site provides insights into the behaviour of the first modern humans to reach Sahul (the combined Pleistocene landmass of Australia, New Guinea and the Aru Islands), including evidence for the early use of hafted edge-ground axes, grinding stones, reflective pigments and a broad diet of plant foods1,2. Human use of the site spans several periods of substantial global climate change between Marine Isotope Stages (MIS) 4 and 1, including the last and penultimate glacial stages (MIS2 and MIS4, respectively)3, and the formation of the Kakadu wetlands in the late Holocene4,5. Documenting palaeoprecipitation and subsequent vegetation and resource changes in the Alligator Rivers region is fundamental to our understanding of the adaptive plasticity of populations entering this region for the first time. However, while there are several early (≥45 ka) archaeological sites from this region1,6,7, the best source of palaeoclimate data for this early occupation phase is located ~800 km to its east in the Gulf of Carpentaria (Fig. 1)8,9. This limits our ability to accurately study local human–environment interaction in this region.

Esri, DigitalGlobe, GeoEye, Earthstar, Geographics, CNES/Airbus DS, USDA, USGS, AeroGRID, IGN, and the GIS User Community (a); panel b adapted with permission from ref. 63, Geoscience Australia
a, A map showing the location of Madjedbebe (star) and all Pleistocene-age environmental records for the monsoonal tropics of northern Sahul (crosses)29,59,60,61. b, The location of the modern and archaeological study sites, including Madjedbebe and other early (≥45ka) archaeological sites (white dots), within and near the Alligator Rivers region (dashed line)1,6,7; the Stuart Highway transect from Darwin to Katherine (white line); and the modern extent of the southern distribution of P. spiralis (slashed line)62.
In other parts of the world, stable isotope analysis of archaeological plant material has emerged as a promising way of studying changes in past precipitation and temperature in direct association with records of human occupation and cultural change10,11,12,13. Such work has tended to focus on Eurasian crops and tree species, with limited applicability to other regions. Here we report on the results of stable carbon isotope analysis of modern and archaeological Pandanus spiralis endocarp (the tough ‘nutshell’ of the pandanus drupe, comprised of sclerenchyma tissue; Extended Data Fig. 1). Pandanus has been found in archaeological sites across the tropics, including as part of Melanesian and Pacific foraging and agricultural systems14,15,16,17, and P. spiralis was recovered from almost all occupation layers at Madjedbebe (Fig. 2)2. We demonstrate that stable carbon isotope analysis of this taxon can provide a reliable palaeoprecipitation proxy, and use it in concert with the analysis of soil stable carbon isotopes, and lithic artefact and exotic raw material discard rates from Madjedbebe, to investigate the relationship between environmental change and human mobility and settlement in the past.

a, P. spiralis trees on the Magela Creek floodplain near Madjedbebe in October 2017, with an inset displaying the cephalium, or aggregate fruit of the tree, comprising ~20 polydrupes (see Supplementary Section 1). b, A scanning electron micrograph of an archaeological fragment of P. spiralis endocarp from Phase 7 (C3/5). sl, seed locule; vb, vascular bundle. Scale bar, 1 mm. c, A scanning electron micrograph of an archaeological fragment of P. spiralis mesocarp from Phase 7 (C3/6). Scale bar, 200 µm. See Florin et al.2 for a detailed identification proof. d, The numbers of fragments of P. spiralis endocarp and P. spiralis mesocarp recovered per litre of soil floated from Madjedbebe, versus increasing depth (m) and decreasing archaeological phase. The dark grey area indicates the numbers of P. spiralis endocarp and mesocarp by litres floated, respectively, and the light grey area indicates the numbers of cf. P. spiralis endocarp and mesocarp by litres floated, respectively. The age estimates are based on the modelled mid-point value of the 95% confidence interval for the start date of each phase1.
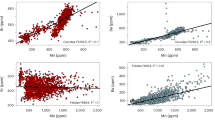
To test whether stable carbon isotope discrimination in P. spiralis endocarp is a useful proxy for the analysis of past fluctuations in mean annual precipitation (MAP), modern P. spiralis drupes were collected from a range of environments in the Northern Territory, and their charred endocarps were analysed for variation in [(13C/12C)sample/(13C/12C)standard] −1 (δ13C) values (Fig. 3; see Extended Data Fig. 2 for the results of charring experiments on the δ13C values of P. spiralis endocarp). Figure 3b depicts δ13C variation between different growth environments (floodplain fringe, seasonal floodways, and open forest and woodland environments) found within a 10-km radius from Madjedbebe rockshelter. Figure 3c documents variation in δ13C values with rainfall within the modern range of distribution of P. spiralis, following a >300-km transect along the Stuart Highway from Darwin to Katherine. Both show a significant correlation between increasing δ13C values and increased water availability (Fig. 3b: one-way analysis of variance, F = 3.572, d.f. = 2, P = 0.032; Fig. 3c: R2 = 0.43, root-mean-square error = 0.75‰, P = <0.001).

a, Samples of P. spiralis collected from different locations along the Stuart Highway, between Darwin and Katherine. MAP is calculated from 24 meteorological sites (see Supplementary Section 2) and displayed as a 10-km buffer surrounding the sampling transect, with blue and yellow indicating higher and lower precipitation, respectively. Additionally, we indicate the mean daily evaporation for June by the fill colour of each dot, which reflects the first month of fruiting of the P. spiralis drupe23. Individual P. spiralis trees are indicated by their specimen number (white numbers; Supplementary Table 3). b, δ13C values of P. spiralis collected from different growth environments (floodplain fringe, seasonal floodways, and open forest and woodland vegetation communities) with the same MAP (~1,510 mm yr−1), near Madjedbebe in the Alligator Rivers region. The grey horizontal band demarcates the interquartile range of the δ13C values of modern P. spiralis from the Alligator Rivers region. Boxes show the interquartile range, the midline shows the median value, and the whiskers extend vertically 1.5 times the interquartile range from the end of the box to the furthest datum within that distance. Data beyond that distance are represented individually as points (‘outliers’). c, A linear relationship is observed between precipitation and δ13C values of P. spiralis with 95% confidence intervals (grey area) and 95% prediction intervals (dashed lines) when removing all sites affected by microclimate effects (blue dots with mean evaporation <5 mm per day). Error bars on data points indicate one standard deviation. Note that the modern δ13C value for P. spiralis collected near Madjedbebe (white dot) falls directly on our best-fit line (sample not used to generate this regression). RMSE, root-mean-square error. The data and R code for this figure are available from ref. 54. Panel a adapted with permission from ref. 63, Geoscience Australia.
These results run counter to many of the observed relationships in C3 plants that link declining MAP and drought conditions to increasing δ13C values of plant tissues18,19. However, these results are in line with expectations of changed C3 carbon isotope discrimination in waterlogged environments20,21. Today, P. spiralis has a wide distribution across northern Australia, where it grows in poorly drained areas of Eucalyptus- and Melaleuca-dominated vegetation communities. This includes along the edges of swamps and billabongs, the fringes of floodplains, and in seasonally wet areas of open forest and woodland. With more than 90% of all precipitation in northern Australia occurring from November to April22, such environments undergo significant seasonal flooding. This means that P. spiralis trees are often waterlogged for several months of the year, including at the beginning of their fruit growth season (P. spiralis fruits mature from June to October)23. The period of waterlogging experienced by P. spiralis plants increases both in regions of higher MAP and in environments more likely to experience prolonged inundation (for example, the fringes of floodplains).
Like other environmental stresses (for example, aridity), waterlogging has been shown to have a negative effect on the photosynthetic rate and, subsequently, the water use efficiency (WUE) of C3 plants20,21. This is understood to be the result of decreased stomata conductance caused by several factors, including decreased root hydraulic conductance, increased oxygen deficiency and subsequent stomata closure. The increased δ13C values evidenced in P. spiralis endocarp growing in environments with higher groundwater availability in this study (for example, with higher MAP or on the fringes of floodplains) are, therefore, probably caused by the effect of prolonged wet season waterlogging experienced by these plants. The only exception to this positive correlation is towards the southern extent of the modern P. spiralis distribution, where δ13C values are higher (that is, WUE is lower; Fig. 3). In terms of precipitation, this is the driest habitat recorded in the transect (~1,150–1,000 mm rainfall per year). However, climate records from this area show significantly reduced evaporation in the early dry season24, which we attribute to cold-air drainage from adjacent high ground (that is, the Arnhem Land Escarpment; Fig. 1b). This reduced evaporation permits standing water to remain longer in this area in June, when P. spiralis drupes are growing despite the reduced precipitation (Extended Data Fig. 3). As such, the lower WUE evidenced near Katherine is also a factor of increased waterlogging during the growing season of P. spiralis drupes. While this means that variability in local topography can cause variability in the effects of MAP on δ13C values of P. spiralis drupes at a particular location, the fit of δ13C values from the direct vicinity of Madjedbebe to the linear relationship observed between precipitation and δ13C values along the Stuart Highway transect suggests that Madjedbebe is an ideal site at which to use this proxy (Fig. 3c).
Figure 4 displays the δ13C values of the archaeological P. spiralis endocarp by archaeological phase from Madjedbebe. Following the Volker-2012a model25, these values have been corrected for changes in the isotopic composition of atmospheric CO2 (δ13CO2ATM) and the partial pressure of CO2 (\(p_{{\mathrm{CO}}_{2}}\)) over the past 65,000 years. Variations in δ13C values in Fig.4a suggest a fluctuating WUE for P. spiralis trees across this period, with decreased WUE during Phase 3 (51.6–28.1 ka), Phase 5 (9.7–8.1 ka) and Phase 7 (about 4 ka to present). Gaps occur in this record in the terminal Pleistocene through to the mid-Holocene owing to decreased recovery of P. spiralis endocarp from this period (Fig. 1c), probably due to reduced preservation. Charred plant remains were recovered at a significantly lower frequency from the alkali environment of the mid- to late Holocene midden (Phases 6 and 7) and the carbonate-enriched sediments directly below it (Phases 4 and 5; see Supplementary Fig. 2 in Florin et al.2). Figure 4b documents variation on a shorter timescale, depicting variations in the corrected δ13C values of the archaeological and modern P. spiralis endocarp over the past 700 years, indicating fluctuations in WUE within Phase 7. On the basis of the modern data, decreased WUE in P. spiralis is most likely a reflection of waterlogging and, therefore, higher precipitation during Phases 3, 5 and 7. In comparison, Phase 2 (65–52.7 ka) and Phase 4 (26.7–13.2 ka) represent periods of higher WUE and, therefore, decreased precipitation. Interestingly, the δ13C values of the modern P. spiralis endocarp are the lowest in the sequence (Fig. 4), suggesting that the lowest recorded precipitation is occurring at present.

a, δ13C values of archaeological P. spiralis endocarp, describing fluctuations in climate at Madjedbebe over the last 65,000 years. The vertical dotted lines demarcate the temporal boundaries of the different archaeological phases, based on the modelled mid-point value of the 95% confidence interval for the start and end date of each phase1. The exception is Phase 7, where AMS radiocarbon mid-point ages for the analysed contexts are used (Supplementary Table 10). The grey horizontal band demarcates the interquartile range of the δ13C values of modern P. spiralis from near Madjedbebe. b, A close-up of δ13C values of archaeological and modern P. spiralis from Madjedbebe and its close surrounds over the last 700 years. The vertical dotted lines demarcate the temporal boundaries of the archaeological P. spiralis samples. The grey horizontal band demarcates the interquartile range of the δ13C values of modern P. spiralis from near Madjedbebe. Boxes show the interquartile range, the midline shows the median value, and the whiskers extend vertically 1.5 times the interquartile range from the end of the box to the furthest datum within that distance. Data beyond that distance are represented individually as points (‘outliers’). The data and R code for this figure are available from ref. 54.
Figure 5 compares the pandanus proxy for palaeoprecipitation, and predicted MAP, with the previously published soil δ13C record1, corrected for changing δ13CO2ATM and \(p_{{\mathrm{CO}}_{2}}\), and the discard of lithic artefacts and exotic raw materials at Madjedbebe. The soil δ13C record is probably a product of both environmental and anthropogenic influences, deriving from a combination of vegetation growing within and immediately outside the site, vegetation brought in by natural mechanisms (for example, wind and water) and vegetation brought in by humans for a range of reasons, including subsistence, bedding and fuel26. This makes its interpretation more complex. In northern Australia, an abundance of C4 plants in the environment correlates with seasonal water availability, with a higher abundance of C4 grasses occurring in locations where precipitation during the summer monsoon season is high27,28. Therefore, despite possible anthropogenic influences, the similar peaks (in Phase 3 and, albeit slightly later, in Phase 6) and troughs (in Phases 2 and 4 and at the end of Phase 7) in both records suggest a general agreement in variation in palaeoprecipitation at the site.

From left to right: soil profile; δ13C values of archaeological P. spiralis endocarp from Square C3 grouped by phase and displayed by mean depth of contexts; predicted MAP at Madjedbebe, based on the mean δ13C values of archaeological P. spiralis endocarp and the modern relationship between δ13C and MAP (Fig. 3c), with 95% confidence interval (grey area) and 95% prediction intervals (dashed lines); δ13C of archaeological soil organic matter samples from Square B2, taken at ~5-cm intervals and grouped by phase; number of lithic artefacts per litres floated for Squares E1 to B3 by depth; number of lithic artefacts produced from exotic raw materials per litres floated for Squares E1 to B3 by depth. Dashed lines on the second, fifth and sixth panels are locally estimated scatterplot smoothing (loess) curves. The data and R code for this figure are available from ref. 54. Soil profile in the left panel adapted with permission from ref. 1, Springer Nature Ltd.
The P. spiralis record suggests decreased precipitation during the penultimate (MIS4) and last glacial (MIS2) stages, although the timing and intensity of the latter is in part masked by poor preservation of archaeological P. spiralis endocarp in the terminal Pleistocene/early Holocene. The δ13C values from the soil record suggest that peak aridity at Madjedbebe may have occurred at the height of MIS2 or the Last Glacial Maximum, about 22–18 ka, in the latter half of Phase 4 (Fig. 5). Precipitation then increased in the early and mid-Holocene, consistent with the early Holocene climatic optimum (about 10–6 ka)29,30. Late onset of aridity in tropical northern Australia is corroborated by other palaeoenvironmental records, which show river flows increasing after 30 ka and declining only from 17 to 10 ka (see Supplementary Section 8 for further discussion of the Madjedbebe proxy in comparison to other northern Australian palaeoenvironmental records)31.
As the lowest WUE of P. spiralis is recorded in the modern record, and there is a general correlation between the soil and pandanus δ13C records, rainfall in the vicinity of Madjedbebe may have rarely, if ever, fallen below present levels (~1,500 mm yr−1) in the past 65,000 years. This attests to moderate precipitation in this region, even throughout the drier stages of MIS4 and MIS2. Indeed, evidence for plant food exploitation and fuel use in Phase 2 at Madjedbebe demonstrates that people had access to a range of vegetation communities during early occupation (MIS4), including freshwater and monsoon vine forest patches2,32. The same is probably also true of the Last Glacial Maximum and terminal Pleistocene. Our results indicate the severity of glacial stages, and their effects on local vegetation during this period may be overestimated in this region. This is unsurprising as modelling studies (for example, Yan et al.33) demonstrate that increased seasonal contrasts during the Last Glacial Maximum enhanced the dynamic forcing of the monsoon in northern Australia. This is partially offset by the reduction in available moisture in the atmosphere, but overall a minor increase in monsoon strength is inferred. Further, this is supported archaeologically by the peaks in artefact discard at Madjedbebe during dry phases (Fig. 5c), which suggest increased use of the site during these phases, and are consistent with earlier arguments that human occupation intensified in the Alligator Rivers region during the Last Glacial Maximum owing to its relatively wet environment34. Peaks in exotic raw materials during dry phases (Fig. 5d) also indicate that people were probably more mobile at these times, with decreased abundance of resources requiring them to extend their foraging range and possibly their social networks35,36.
Discussion
As there are little data for palaeoprecipitation levels during MIS4 across northern Australia, let alone in the Alligator Rivers region, this proxy provides valuable environmental context for early settlement of the continent. Modern human populations entering this region 65,000 years ago would have experienced a relatively dry, but not arid, environment, with small pockets of monsoon vine forest and freshwater vegetation available on the Alligator Rivers lowlands2,32. As climate ameliorated in Phase 3, monsoon vine forest environments probably increased in the vicinity of Madjedbebe37. At this time, people frequenting the rockshelter probably reduced their residential range, using fewer exotic raw materials for artefact manufacture. They also ceased the production of certain distinctive lithic artefact types, including thinning flakes, convergent flakes and discoidal core technology1. It is during this phase that human occupation is evidenced in other parts of Sahul17,38,39,40,41,42.
The efficacy of pandanus δ13C as a proxy for palaeoprecipitation is a potential game-changer for palaeoecological and archaeological work in Australia, Melanesia and the Pacific. As in other regions, palaeoprecipitation records from archaeological sites are often lacking. This has meant that the links between climatic changes, human modification of landscape and human behaviour often refer to off-site and distant lake or marine records that may be unreliable indicators of local conditions. Across the tropics, where pandanus and other tree nuts are ubiquitous and well preserved, the isotopic analysis of such food plant remains offers a method for the development of context-specific proxies for past environment and land management, and their comparison to cultural, social and technological changes.
Methods
Charring experiment for modern P. spiralis
To test the effect of carbonization on the δ13C value of P. spiralis, carbon isotope analysis was carried out on the nutshell of 108 drupes from 27 P. spiralis trees under different states of physical pre-treatment. Four drupes from each tree were analysed, each following a different method of physical pre-treatment: dried; charred for 4 h at 400 °C; charred for 2 h at 500 °C; and charred in a multi-temperature open fire. The controlled charring experiments were performed in a low-oxygen state in a kiln, following the protocol set out in Fraser et al.43. Open-fire conditions exceeded 1,300 °C, covering the maximum range of ethnographic and experimental hearths44.
In line with the archaeological specimens, samples also underwent chemical pre-treatment. The charred samples underwent ABA pre-treatment. This procedure consisted of an acid treatment (10 ml of 2 M HCl at 60 °C for 2 h), two base treatments (10 ml of 0.25% NaOH at room temperature for 30 min) and another acid treatment (10 ml of 2 M HCl at room temperature for 2 h). The sample was rinsed three times with Milli-Q water between treatments and six times following the final acid treatment, before it was dried in an oven at 60 °C overnight and powdered with a glass rod.
The dried samples were milled and then underwent α-cellulose pre-treatment45. This procedure consisted of a Soxhlet extraction treatment (2:1 cyclohexane–ethanol mixture for 6 h, ethanol for 6 h, Milli-Q water for 6 h), a lignin extraction treatment (NaClO2 oxidation (15 g l−1) under acidified conditions (pH of ~3) at 60 °C for 2 h), a cellulose extraction treatment (12% NaOH at 60 °C under nitrogen gas, and then 7% NaOH at 60 °C under nitrogen gas) and an acid treatment (2 M HCl at room temperature for 1 h). The sample was then dried in an oven at 60 °C overnight.
The results of this experiment are reported in Extended Data Fig. 2.
Rainfall and environment for modern P. spiralis
Stable carbon isotope analysis was carried out on the endocarp of 201 modern P. spiralis fruits. These fruits came from two areas: Mirarr country near Madjedbebe, Alligator Rivers region; and the edge of the Stuart Highway, between Darwin and Katherine (Figs. 1 and 3). The former allowed for the analysis of variation in the δ13C values of P. spiralis nutshell from environments local to Madjedbebe, including floodplain fringe, seasonal floodways, and open forest and woodland environments. The latter allowed for the analysis of variation in the δ13C values of P. spiralis nutshell across different mean annual rainfall zones, ranging from ~1,700–1,600 mm yr−1 in Darwin to ~1,150–1,000 mm yr−1 in Katherine. Supplementary Table 3 gives details of the site locations, collection dates, MAP, mean daily evaporation and habitat of each of the P. spiralis analysed.
After the initial study examining the effect of carbonization on the δ13C value of P. spiralis, all modern samples were charred at 500 °C for 2 h before analysis and then cut into pieces with a scalpel. The temperature of 500 °C was chosen as it is a commonly prescribed combustion temperature for loss-on-ignition protocols for plant materials46. In line with the archaeological specimens, samples also underwent ABA pre-treatment.
Modern climate
To test whether the 13C isotopic signature of P. spiralis drupes are directly related to water availability, we evaluated two major contributors to the regional water budget, precipitation and evaporation, and produced regional interpolations. All data from the Northern Territory were collected from the Bureau of Meteorology online data page (http://www.bom.gov.au/climate/averages/tables/ca_nt_names.shtml)24. MAP was determined from all collected measurements of daily total precipitation during the full observation time utilizing standard rain gauges. Mean daily evaporation was determined using all data collected for the month of June from a class A evaporation pan. June daily evaporation was selected because P. spiralis fruits from June to October23, with a large amount of their biomass laid down in June (authors’ own observation). Consequently, the 13C isotopic signature preserved within the drupe should reflect this interval.
To create regional precipitation and evaporation maps from meteorological sites, we utilized inverse-distance-weighted (IDW) interpolation. This was chosen as stations along our transect were sparsely available. IDW is a deterministic spatial interpolation technique that assumes spatially known data are strongly correlated to distance such that the local influences of neighbouring sites diminish with increasing distance47. This technique is particularly useful when datasets are too small to utilize more complex interpolation methods (that is, kriging, spline or multiple regression techniques)48. However, IDW is sensitive to site clustering and is optimized when samples are evenly spaced. Therefore, in locations with multiple meteorological stations we selected only one site for inclusion in our analyses. Sites were chosen on the basis of the proximity to the areas of interest and the completeness of climate records, specifically over the last decade.
In total, we used 24 precipitation stations and 12 evaporation stations for our analyses (Supplementary Figs. 4 and 5 and Supplementary Tables 1 and 2). Each meteorological station location was placed in ArcGIS (version 10.6) software and projected to the GDA-94 coordinate system. In the ArcGIS Geostatistical Analyst tools, we selected IDW. For both precipitation and evaporation, we elected to use simple interpolation parameters in our model to avoid bias outcomes and to ensure reproducible results. For this reason, we selected a standard neighbour with one sector and equal major and minor semi-axes such that all unknown sites had equally weighted predictions in all directions. The power value of two (p = 2, scaling factor that determines the influence of distance) places a higher weight on closer locations because climate gradients are strong along our transect. We determined the predicted precipitation and evaporation values from all available meteorological stations at a 0.25-km2 resolution for the entire study area. Data were extracted for each sample location using the Spatial Analyst tool Extract Multi Values to Points.
Archaeological P. spiralis
Charred plant macrofossils were recovered from all levels of occupation at Madjedbebe via flotation1,2. Archaeobotanical analysis was carried out on the flot and heavy residue of all excavated hearths, and 100% of the Pleistocene and 50% of the Holocene spits from a 1 × 1-m sediment column (Square C3/C2). Carbon isotope analysis was carried out on all identified specimens of polydrupe Pandanus sp. endocarp that were >2 mm in size (Supplementary Table 4). These specimens were determined to be fragments of P. spiralis both as they were found in conjunction with more fragile P. spiralis mesocarp fragments (Fig. 2b–d); and as Madjedbebe is a lowland site, approximately 10 km from the elevated sandstone plateaus where the other northern Australian polydrupe species, P. basedowii, occurs.
Many of the charred plant macrofossils recovered from Madjedbebe have been shown through previous pre-treatment for accelerated mass spectrometry (AMS) radiocarbon dating to be significantly degraded49, probably owing to the replacement of charcoal by humic substances over time via a process of oxidative degradation50. Therefore, following standard procedure, all specimens underwent ABA pre-treatment to remove any humic, fluvic or carbonate contaminants before analysis51. This treatment was the same as that performed on modern samples. However, as many of the archaeological samples were significantly degraded, they required repetitions of the base stage of pre-treatment to be carried out, in some cases up to 16 times. Between base treatments, the specimens were washed once with Milli-Q water, allowing for the examination of the clarity of the sample and, therefore, the quantity of contaminants still present. Once samples were found to be clear, they underwent a final base treatment before moving to the second stage of acid treatment. Of the 244 archaeological samples analysed, 104 dissolved during pre-treatment (Supplementary Table 4). As δ13C measurements are less sensitive to contamination than 14C measurements and many of the archaeological specimens dissolved during pre-treatment, we recommend that future studies test the necessity of ABA pre-treatment for stable isotope analyses.
Stable carbon isotope analysis
All modern and archaeological samples were weighed (~30–60 µg) into tin capsules for analysis. They were then analysed across 37 runs on an Elementar VarioMICRO Elemental Analyser and an IsoPrime Continuous-Flow Isotope Ratio Mass Spectrometer at the Australian Nuclear Science and Technology Organisation. The δ13C results were normalized to an International Atomic Energy Agency reference material, IAEA C8, with a consensus value of δ13CVPDB = −18.31‰ (refs. 52,53), where VPDB represents Vienna PeeDee Belemnite, and analysed with the commercial reference standards High Organic Sediment Standard OAS (Elemental Microanalysis, catalogue no. B2151) and Protein Standard OAS (Elemental Microanalysis, catalogue no. B2155) as quality control references. Supplementary Tables 5–9 list the results for the standards used. Unless otherwise stated (Supplementary Tables 3 and 4), all results listed are the mean of repeat measurements with the standard deviation of the replicate analyses less than or equal to ±0.3.
Soil analysis
A 2-g sub-sample of sediment from selected contexts (see Supplementary Section 7) was ground to a fine powder and treated with 2 M HCl for 24 h to remove inorganic carbon. National Bureau of Standards NBS-19 was used to normalize the data to the VPDB scale. Analytical precision for replicate measurements of δ13C in NBS-19 was ±0.2‰ (2σ). The results of this analysis were previously reported in Clarkson et al.1.
Adjustment of δ13C for changes in past δ13CO2ATM and \(p_{{\mathrm{CO}}_{2}}\)
To compare the archaeological dataset with the modern isotopic analysis presented in this paper, all archaeological samples were corrected for changes in both δ13CO2ATM and \(p_{{\mathrm{CO}}_{2}}\). This was carried out using a series of offsets calculated for each phase following the Volker-2012a model25. Supplementary Table 4 lists both the original and adjusted δ13C values.
The soil δ13C, initially reported in Clarkson et al.1, was also corrected following the Volker-2012a model25. Both the original and adjusted δ13C values are available from ref. 54.
Lithic analysis
All analysed stone artefacts from Squares E1 to B3 >3 mm in size were counted and identified to raw material type. The three rows closest to the back wall of the site were chosen as they incorporate and surround the flotation squares (C2 and C3). Raw materials not locally found include several varieties of exotic quartzite, silcrete, chert, Oenpelli dolerite, hornfels, basalt, non-local sandstone, a fine-grained vesicular silicate and rock crystal quartz. Although many of these raw materials are of unknown provenance, some are known to be from quite distant sources (such as silcrete and Oenpelli dolerite).
AMS radiocarbon dating
Three charcoal samples from C3/4, C3/5 and C3/7 were identified and pre-treated using the ABA method. The pre-treated samples were combusted and then converted to graphite55. Radiocarbon analysis was carried out using the VEGA AMS Facility at ANSTO56. The radiocarbon results were converted to calendar ages using the SHCal13 data57 and the OxCal program58.
The AMS radiocarbon dates are reported in Supplementary Table 10.
Reporting Summary
Further information on research design is available in the Nature Research Reporting Summary linked to this article.
Data availability
All elements necessary to allow interpretation and replication of the results, including full datasets, are provided in the Supplementary Information. R code and additional data for Figs. 3–5 are available from ref. 54. Archaeobotanical material analysed in this study will be kept in the Archaeology Laboratories of The University of Queensland until 2021. It will then be deposited in a Gundjeihmi Aboriginal Corporation keeping place. The material will be accessible upon request from Gundjeihmi Aboriginal Corporation (gundjeihmi@mirarr.net). The language, images and information contained in this publication include reference to Indigenous knowledge including traditional knowledge, traditional cultural expression and references to biological resources (plants and animals) of the Mirarr people. The source Indigenous knowledge is considered “Confidential Information”; traditional law and custom applies to it and the Mirarr people assert copyright over it in addition to any copyright in the complete work. Any Mirarr-related language, images and information are published with the consent of Gundjeihmi Aboriginal Corporation as the representative of the Mirarr people for the purposes of general education purposes. No further use and absolutely no commercial use is authorized without the prior consent and agreement of the Mirarr people. Please contact Gundjeihmi Aboriginal Corporation to request permission to refer to any Indigenous knowledge in this publication.
References
Clarkson, C. et al. Human occupation of northern Australia by 65,000 years ago. Nature 547, 306–310 (2017).
Florin, S. A. et al. The first Australian plant foods at Madjedbebe, 65,000–53,000 years ago. Nat. Commun. https://doi.org/10.1038/s41467-020-14723-0 (2020).
Nanson, G. C., Price, D. M. & Short, S. A. Wetting and drying of Australia over the past 300 ka. Geology 20, 791–794 (1992).
Clark, R. L. & Guppy, J. C. A transition from mangrove forest to freshwater wetland in the monsoon tropics of Australia. J. Biogeogr. 15, 665–684 (1988).
Woodroffe, C. D., Thom, B. G. & Chappell, J. Development of widespread mangrove swamps in mid-Holocene times in northern Australia. Nature 317, 711–713 (1985).
David, B. et al. Nawarla Gabarnmang, a 45,180±910 cal BP site in Jawoyn country, southwest Arnhem Land Plateau. Aust. Archaeol. 73, 73–77 (2011).
Roberts, R. G. et al. The human colonisation of Australia: optical dates of 53,000 and 60,000 years bracket human arrival at Deaf Adder Gorge, Northern Territory. Quat. Geochronol. 13, 575–583 (1994).
Chivas, A. R. et al. Sea-level and environmental changes since the last interglacial in the Gulf of Carpentaria, Australia: an overview. Quat. Int. 83–85, 19–46 (2001).
Torgersen, T. et al. Late Quaternary environments of the Carpentaria Basin, Australia. Palaeogeogr. Palaeoclim. Palaeoecol. 67, 245–261 (1988).
Aguilera, M., Ferrio, J. P., Pérez, G., Araus, J. L. & Voltas, J. Holocene changes in precipitation seasonality in the western Mediterranean Basin: a multi-species approach using δ13C of archaeobotanical remains. J. Quat. Sci. 27, 192–202 (2012).
Heaton, T. H. E. Spatial, species, and temporal variations in the 13C/12C ratios of C3 plants: implications for palaeodiet studies. J. Archaeol. Sci. 26, 637–649 (1999).
Wallace, M. et al. Stable carbon isotope analysis as a direct means of inferring crop water status and water management practices. World Archaeol. 45, 388–409 (2013).
Wallace, M. P. et al. Stable carbon isotope evidence for Neolithic and Bronze Age crop water management in the Eastern Mediterranean and Southwest Asia. PLoS ONE 10, e0127085 (2015).
Denham, T. P. et al. Origins of agriculture at Kuk Swamp in the highlands of New Guinea. Science 301, 189–193 (2003).
Kirch, P. V. Second millennium B.C. arboriculture in Melanesia: archaeological evidence from the Mussau Islands. Econ. Bot. 43, 225–240 (1989).
Matthews, P. J. & Gosden, C. Plant remains from waterlogged sites in the Arawe Islands, West New Britain Province, Papua New Guinea: implications for the history of plant use and domestication. Econ. Bot. 51, 121–133 (1997).
Summerhayes, G. R. et al. Human adaptation and plant use in highland New Guinea 49,000 to 44,000 years ago. Science 330, 78–81 (2010).
Farquhar, G. D. & Richards, R. A. Isotopic composition of plant carbon correlates with water-use efficiency of wheat genotypes. Aust. J. Plant Physiol. 11, 539–552 (1984).
Kohn, M. J. Carbon isotope compositions of terrestrial C3 plants as indicators of (paleo)ecology and (paleo)climate. Proc. Natl Acad. Sci. USA 107, 19691–19695 (2010).
Ashraf, M. A. Waterlogging stress in plants: a review. Afr. J. Agric. Res. 7, 1976–1981 (2012).
Pezeshki, S. R. Wetland plant responses to soil flooding. Environ. Exp. Bot. 46, 299–312 (2001).
Eamus, D., O’Grady, A. P. & Hutley, L. Dry season conditions determine wet season water use in the wet–dry tropical savannas of northern Australia. Tree Physiol. 20, 1219–1226 (2000).
Cowie, I. D., Short, P. S. & Osterkamp Madsen, M. Floodplain Flora: a Flora of the Coastal Floodplains of the Northern Territory, Australia Vol. 10 (Australian Biological Resources Study, 2000).
Climate Data Online (Bureau of Meteorology, 2020); http://www.bom.gov.au/climate/data/
Hare, V. J., Loftus, E., Jeffrey, A. & Ramsey, C. B. Atmospheric CO2 effect on stable carbon isotope composition of terrestrial fossil archives. Nat. Commun. 9, 252 (2018).
Parker, A. G., Lee-Thorp, J. & Mitchell, P. J. Late Holocene Neoglacial conditions from the Lesotho highlands, southern Africa: Phytolith and stable carbon isotope evidence from the archaeological site of Likoaeng. Proc. Geol. Assoc. 122, 201–211 (2011).
Andrae, J. W. et al. Initial expansion of C4 vegetation in Australia during the late Pliocene. Geophys. Res. Lett. 45, 4831–4840 (2018).
Murphy, B. P. & Bowman, D. M. J. S. Seasonal water availability predicts the relative abundance of C3 and C4 grasses in Australia. Glob. Ecol. Biogeogr. 16, 160–169 (2007).
Shulmeister, J. & Lees, B. G. Pollen evidence from tropical Australia for the onset of an ENSO-dominated climate at c. 4000 BP. Holocene 5, 10–18 (1995).
Woodward, C. et al. A Holocene record of climate and hydrological changes from Little Llangothlin Lagoon, south eastern Australia. Holocene 24, 1665–1674 (2014).
Reeves, J. M. et al. Palaeoenvironmental change in tropical Australasia over the last 30,000 years – a synthesis by the OZ-INTIMATE group. Quat. Sci. Rev. 74, 97–114 (2013).
Carah, X. A. Regime Change: an Anthracological Assessment of Fuel Selection and Management at Madjedbebe (Malkanunanja II), Mirarr Country, Australia. PhD thesis, Univ. Queensland (2017).
Yan, M., Wang, B., Liu, J., Zhu, A. & Ning, L. Understanding the Australian Monsoon change during the Last Glacial Maximum with a multi-model ensemble. Climate 14, 2037–2052 (2018).
Veth, P. Islands in the interior: a model for the colonization of Australia’s arid zone. Archaeol. Ocean. 24, 81–92 (1989).
Shott, M. J. Technological organization and settlement mobility: an ethnographic examination. J. Anthropol. Res. 42, 15–51 (1986).
Gould, R. A. & Saggers, S. Lithic procurement in Central Australia: a closer look at Binford’s idea of embeddedness in archaeology. Am. Antiquity 50, 117–136 (1985).
Fensham, R. in Flammable Australia: Fire Regimes, Biodiversity and Ecosystems in a Changing World (eds Williams, R. et al.) 173–193 (CSIRO Publishing, 2012).
Bowler, J. M. et al. New ages for human occupation and climate change at Lake Mungo, Australia. Nature 421, 837–840 (2003).
Hamm, G. et al. Cultural innovation and megafauna interaction in the early settlement of arid Australia. Nature 539, 280–283 (2016).
Maloney, T., O’Connor, S., Wood, R., Aplin, K. & Balme, J. Carpenters Gap 1: a 47,000 year old record of indigenous adaption and innovation. Quat. Sci. Rev. 191, 204–228 (2018).
McDonald, J. et al. Karnatukul (Serpent’s Glen): a new chronology for the oldest site in Australia’s Western Desert. PLoS ONE 13, e0202511 (2018).
Veth, P. et al. Early human occupation of a maritime desert, Barrow Island, North-West Australia. Quat. Sci. Rev. 168, 19–29 (2017).
Fraser, R. A. et al. Assessing natural variation and the effects of charring, burial and pre-treatment on the stable carbon and nitrogen isotope values of archaeobotanical cereals and pulses. J. Archaeol. Sci. 40, 4754–4766 (2013).
Bentsen, S. E. Using pyrotechnology: fire-related features and activities with a focus on the African Middle Stone Age. J. Archaeol. Res. 22, 141–175 (2013).
Hua, Q. et al. Radiocarbon in tropical tree rings during the Little Ice Age. Nucl. Instrum. Methods Phys. Res. B 223–224, 489–494 (2004).
Nelson, D. W. & Sommers, L. E. in Methods of Soil Analysis, Part 3: Chemical Methods Vol. 14 (eds Sparks, D. L. et al.) Ch. 34 (Wiley, 1996).
Lu, G. Y. & Wong, D. W. An adaptive inverse-distance weighting spatial interpolation technique. Comput. Geosci. 34, 1044–1055 (2008).
Longley, P. A., Goodchild, M. F., Maguire, D. J. & Rhind, D. W. Geographic Information Systems and Sciences 2nd edn (Wiley, 2005).
Clarkson, C. et al. Reply to comments on Clarkson et al. (2017) ‘Human occupation of northern Australia by 65 000 years ago’. Aust. Archaeol. 84, 84–89 (2018).
Ascough, P. L., Bird, M. I., Francis, S. M. & Lebl, T. Alkali extraction of archaeological and geological charcoal: evidence for diagenetic degradation and formation of humic acids. J. Archaeol. Sci. 38, 69–78 (2011).
Roberts, P. et al. Calling all archaeologists: guidelines for terminology, methodology, data handling, and reporting when undertaking and reviewing stable isotope applications in archaeology. Rapid Commun. Mass Spectrom. 32, 361–372 (2018).
Clercq, M. L., Van Der Plicht, J. & Gröning, M. New 14C reference materials with activities of 15 and 50 pMC. Radiocarbon 40, 295–297 (2016).
Gonfiantini, R., Stichler, W. & Rozanski, K. in Reference and Intercomparison Materials for Stable Isotopes of Light Elements 13–29 (IAEA, 1995).
Marwick, B. & Florin, S. A. Pandanus nutshell generates a palaeoprecipitation record for human occupation at Madjedbebe, northern Australia. OSF https://doi.org/10.17605/OSF.IO/9TN4F (2020).
Hua, Q. et al. Progress in radiocarbon target preparation at the ANTARES AMS Centre. Radiocarbon 43, 275–282 (2001).
Fink, D. et al. The ANTARES AMS facility at ANSTO. Nucl. Instrum. Methods Phys. Res. B 223–224, 109–115 (2004).
Hogg, A. G. et al. SHCal13 Southern Hemisphere calibration, 0–50,000 years cal BP. Radiocarbon 55, 1889–1903 (2013).
Bronk Ramsey, C. Bayesian analysis of radiocarbon dates. Radiocarbon 51, 337–360 (2009).
Field, E., McGowan, H. A., Moss, P. T. & Marx, S. K. A late Quaternary record of monsoon variability in the northwest Kimberley, Australia. Quat. Int. 449, 119–135 (2017).
Rowe, C. et al. Vegetation over the last glacial maximum at Girraween Lagoon, monsoonal northern Australia. Quat. Res. https://doi.org/10.1017/qua.2020.50 (2020).
van der Kaars, S. Palynology of eastern Indeonesian marine piston-cores: a late Quaternary vegetational and climatic record for Australasia. Palaeogeogr. Palaeoclim. Palaeoecol. 85, 239–302 (1991).
Occurrence records download on 2020-11-07. Atlas of Living Australia https://doi.org/10.26197/ala.ffefeb96-7b7c-421b-8cf2-0968c938f982 (2020).
Hutchinson, M. F., Stein, J. L., Stein, J. A., Anderson, H. & Tickle, P. K. GEODATA 9 second DEM and D8: Digital Elevation Model Version 3 and Flow Direction Grid 2008 (Geoscience Australia, 2008); https://ecat.ga.gov.au/geonetwork/srv/eng/catalog.search#/metadata/66006
Acknowledgements
We are grateful to the custodians of Madjedbebe, the Mirarr Senior Traditional Owners (Y. Margarula and M.N.) and our research partner, Gundjeihmi Aboriginal Corporation, for permission to carry out this research and publish this paper. We are also grateful to J. O’Brien and D. Vadiveloo for assistance in the field. We thank X. Carah for implementing the archaeobotanical recovery programme at Madjedbebe and for his assistance alongside M. Harding, K. Norman, A. Rogers, E. Hayes and K. Connell in the collection of modern pandanus drupes for isotopic analysis. We also thank M. Smith and M. Forbes for their thoughtful comments on this manuscript; any errors are, however, our own. The archaeobotanical and isotopic research conducted by S.A.F. was supported by an Australian Institute of Nuclear Science and Engineering Postgraduate Research Award (11877), a Wenner-Gren Dissertation Fieldwork Grant (9260), a Dan David Scholarship, an Australian Research Council Research Training Program scholarship and an ARC Centre of Excellence for Australian Biodiversity and Heritage Irinjili Research Training Program Internship for Women. The initial fieldwork and excavation of Madjedbebe was funded by an ARC Discovery Project (DP110102864) obtained by C.C., B.M., R.F., L.A.W. and M. Smith. P.R. thanks the Max Planck Society for funding.
Funding
Open access funding provided by Max Planck Society.
Author information
Authors and Affiliations
Contributions
S.A.F., P.R., A.S.F. and C.C. conceptualized the project and wrote the main text, with specialist assistance from J.S., N.R.P., C.E.L., B.M., L.A.B. and Q.H. S.A.F. and A.S.F. undertook the archaeobotanical analysis. S.A.F. undertook the modern collection, with M.N. and Dj.Dj. within the Alligator Rivers region. M.N. and Dj.Dj. provided cultural knowledge. L.A.B. and S.A.F. undertook the isotopic analysis. Q.H. and S.A.F. undertook the radiocarbon analysis. C.C. undertook the lithic analysis. B.M., N.R.P., S.A.F., C.C., A.S.F. and P.R. created the figures. S.A.F. obtained the funding for the plant collection, and isotopic and AMS radiocarbon analysis. C.C., R.F., B.M. and L.A.W. obtained the funding for and conducted the excavations. All authors edited the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Peer review information Nature Ecology & Evolution thanks Peter Veth and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Peer reviewer reports are available.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Schematic of a drupe from a polydrupe or syncarpous Pandanus spp.
Note that portions of both the endocarp and the mesocarp are found archaeologically. Figure adapted from ref. 2, Springer Nature Ltd.
Extended Data Fig. 2 Results of the modern Pandanus spiralis charring experiment.
Compares the δ13C values of endocarp from the same trees prepared with different physical pre-treatment methods. There is a significant difference between the dried and charred δ13C values (One-way ANOVA, F = 2.81, df = 111, p = 0.043). This may be caused by the variation in chemical pre-treatment methods used (soxhlet extraction pre-treatment, rather than ABA pre-treatment, was used for the dried specimens; see Methods) or by the process of charring itself. However, there is little difference between the δ13C values of the charred specimens (One-way ANOVA, F = 0.12, df = 83, p = 0.883). This is despite the fact that one charring method occurred on an open fire, which reached temperatures > 1300 °C. Therefore, as the archaeological specimens are also charred and have been chemically pre-treated using the same method as the modern specimens, any change in δ13C through charring is not expected to affect the results.
Extended Data Fig. 3 Modern precipitation and evaporation data from Darwin, Pine Creek and Katherine.
a) Map of the Stuart Highway transect from Darwin to Katherine (white line), showing proximity to Madjedbebe; b) and c) Mean annual precipitation and daily evaporation data, respectively, from Darwin, Pine Creek and Katherine. Panel a adapted with permission from ref. 63, Geosciences Australia, and panel b and c adapted with permission from ref. 30, Bureau of Meteorology.
Supplementary information
Supplementary Information
Supplementary Notes 1–8, Figs. 1–5, Tables 1–10 and references.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Florin, S.A., Roberts, P., Marwick, B. et al. Pandanus nutshell generates a palaeoprecipitation record for human occupation at Madjedbebe, northern Australia. Nat Ecol Evol 5, 295–303 (2021). https://doi.org/10.1038/s41559-020-01379-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41559-020-01379-8
This article is cited by
-
The history of phytolith research in Australasian archaeology and palaeoecology
Vegetation History and Archaeobotany (2023)
-
65,000-years of continuous grinding stone use at Madjedbebe, Northern Australia
Scientific Reports (2022)
