
Abstract
Membraneless organelles or condensates form through liquid–liquid phase separation1,2,3,4, which is thought to underlie gene transcription through condensation of the large-scale nucleolus5,6,7 or in smaller assemblies known as transcriptional condensates8,9,10,11. Transcriptional condensates have been hypothesized to phase separate at particular genomic loci and locally promote the biomolecular interactions underlying gene expression. However, there have been few quantitative biophysical tests of this model in living cells, and phase separation has not yet been directly linked with dynamic transcriptional outputs12,13. Here, we apply an optogenetic approach to show that FET-family transcriptional regulators exhibit a strong tendency to phase separate within living cells, a process that can drive localized RNA transcription. We find that TAF15 has a unique charge distribution among the FET family members that enhances its interactions with the C-terminal domain of RNA polymerase II. Nascent C-terminal domain clusters at primed genomic loci lower the energetic barrier for nucleation of TAF15 condensates, which in turn further recruit RNA polymerase II to drive transcriptional output. These results suggest that positive feedback between interacting transcriptional components drives localized phase separation to amplify gene expression.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout





Similar content being viewed by others

Data availability
All data supporting the findings of this study are available from the corresponding author on reasonable request. Source data are provided with this paper.
Code availability
Custom code used to process and analyse the images, as detailed in the Methods, are available upon request.
References
Brangwynne, C. P. et al. Germline P granules are liquid droplets that localize by controlled dissolution/condensation. Science 324, 1729–1732 (2009).
Li, P. et al. Phase transitions in the assembly of multivalent signalling proteins. Nature 483, 336–340 (2012).
Banani, S. F., Lee, H. O., Hyman, A. A. & Rosen, M. K. Biomolecular condensates: organizers of cellular biochemistry. Nat. Rev. Mol. Cell Biol. 18, 285–298 (2017).
Shin, Y. & Brangwynne, C. P. Liquid phase condensation in cell physiology and disease. Science 357, eaaf4382 (2017).
Brangwynne, C. P., Mitchison, T. J. & Hyman, A. A. Active liquid-like behavior of nucleoli determines their size and shape in Xenopus laevis oocytes. Proc. Natl Acad. Sci. USA 108, 4334–4339 (2011).
Berry, J., Weber, S. C., Vaidya, N., Haataja, M. & Brangwynne, C. P. RNA transcription modulates phase transition-driven nuclear body assembly. Proc. Natl Acad. Sci. USA 112, E5237–E5245 (2015).
Falahati, H. & Wieschaus, E. Independent active and thermodynamic processes govern the nucleolus assembly in vivo. Proc. Natl Acad. Sci. USA 114, E3585–E3585 (2017).
Cho, W. K. et al. Mediator and RNA polymerase II clusters associate in transcription-dependent condensates. Science 361, 412–415 (2018).
Sabari, B. R. et al. Coactivator condensation at super-enhancers links phase separation and gene control. Science 361, eaar3958 (2018).
Chong, S. S. et al. Imaging dynamic and selective low-complexity domain interactions that control gene transcription. Science 361, eaar2555 (2018).
Boija, A. et al. Transcription factors activate genes through the phase-separation capacity of their activation domains. Cell 175, 1842–1855 (2018).
McSwiggen, D. T., Mir, M., Darzacq, X. & Tjian, R. Evaluating phase separation in live cells: diagnosis, caveats, and functional consequences. Genes Dev. 33, 1619–1634 (2019).
Peng, A. & Weber, S. C. Evidence for and against liquid–liquid phase separation in the nucleus. Noncoding RNA 5, 1–14 (2019).
Elbaum-Garfinkle, S. et al. The disordered P granule protein LAF-1 drives phase separation into droplets with tunable viscosity and dynamics. Proc. Natl Acad. Sci. USA 112, 7189–7194 (2015).
Nott, T. J. et al. Phase transition of a disordered nuage protein generates environmentally responsive membraneless organelles. Mol. Cell 57, 936–947 (2015).
Smith, J. et al. Spatial patterning of P granules by RNA-induced phase separation of the intrinsically-disordered protein MEG-3. eLife 5, e21337 (2016).
Feric, M. et al. Coexisting liquid phases underlie nucleolar subcompartments. Cell 165, 1686–1697 (2016).
Wang, J. et al. A molecular grammar governing the driving forces for phase separation of prion-like RNA binding proteins. Cell 174, 688–699 (2018).
Mitrea, D. M. et al. Nucleophosmin integrates within the nucleolus via multi-modal interactions with proteins displaying R-rich linear motifs and rRNA. eLife 5, e13571 (2016).
Schwartz, J. C., Cech, T. R. & Parker, R. R. Biochemical properties and biological functions of FET proteins. Annu. Rev. Biochem. 84, 355–379 (2015).
Kato, M. et al. Cell-free formation of RNA granules: low complexity sequence domains form dynamic fibers within hydrogels. Cell 149, 753–767 (2012).
Patel, A. et al. A liquid-to-solid phase transition of the ALS protein FUS accelerated by disease mutation. Cell 162, 1066–1077 (2015).
Burke, K. A., Janke, A. M., Rhine, C. L. & Fawzi, N. L. Residue-by-residue view of in vitro FUS granules that bind the C-terminal domain of RNA polymerase II. Mol. Cell 60, 231–241 (2015).
Murthy, A. C. et al. Molecular interactions underlying liquid−liquid phase separation of the FUS low-complexity domain. Nat. Struct. Mol. Biol. 26, 637–648 (2019).
Kwon, I. et al. Phosphorylation-regulated binding of RNA polymerase II to fibrous polymers of low-complexity domains. Cell 155, 1049–1060 (2013).
Suter, D. M. et al. Mammalian genes are transcribed with widely different bursting kinetics. Science 332, 472–474 (2011).
Shin, Y. et al. Spatiotemporal control of intracellular phase transitions using light-activated optodroplets. Cell 168, 159–171 (2017).
Riback, J. A. et al. Composition dependent thermodynamics of intracellular phase separation. Nature 581, 209–214 (2020).
Hein, M. Y. et al. A human interactome in three quantitative dimensions organized by stoichiometries and abundances. Cell 163, 712–723 (2015).
Bracha, D. et al. Mapping local and global liquid phase behavior in living cells using photo-oligomerizable seeds. Cell 175, 1467–1480 (2018).
Sanders, D. W. et al. Competing protein–RNA interaction networks control multiphase intracellular organization. Cell 181, 306–324 (2020).
Lin, Y., Currie, S. L. & Rosen, M. K. Intrinsically disordered sequences enable modulation of protein phase separation through distributed tyrosine motifs. J. Biol. Chem. 292, 19110–19120 (2017).
Sherry, K. P., Das, R. K., Pappu, R. V. & Barrick, D. Control of transcriptional activity by design of charge patterning in the intrinsically disordered RAM region of the Notch receptor. Proc. Natl Acad. Sci. USA 114, E9243–E9252 (2017).
Statt, A., Casademunt, H., Brangwynne, C. P. & Panagiotopoulos, A. Z. Model for disordered proteins with strongly sequence-dependent liquid phase behavior. J. Chem. Phys. 152, 075101 (2020).
Wei, M. T. et al. Phase behaviour of disordered proteins underlying low density and high permeability of liquid organelles. Nat. Chem. 9, 1118–1125 (2017).
Harlen, K. M. & Churchman, L. S. The code and beyond: transcription regulation by the RNA polymerase II carboxy-terminal domain. Nat. Rev. Mol. Cell Biol. 18, 263–273 (2017).
Janke, A. M. et al. Lysines in the RNA polymerase II C-terminal domain contribute to TAF15 fibril recruitment. Biochemistry 57, 2549–2563 (2018).
Boehning, M. et al. RNA polymerase II clustering through carboxy-terminal domain phase separation. Nat. Struct. Mol. Biol. 25, 833–840 (2018).
Kwiatkowski, N. et al. Targeting transcription regulation in cancer with a covalent CDK7 inhibitor. Nature 511, 616–620 (2014).
Wang, Y. B. et al. CDK7-dependent transcriptional addiction in triple-negative breast cancer. Cell 163, 174–186 (2015).
Cisse, I. I. et al. Real-time dynamics of RNA polymerase II clustering in live human cells. Science 341, 664–667 (2013).
Cho, W. K. et al. RNA polymerase II cluster dynamics predict mRNA output in living cells. eLife 5, e13617 (2016).
Casse, C., Giannoni, F., Nguyen, V. T., Dubois, M. F. & Bensaude, O. The transcriptional inhibitors, actinomycin D and α-amanitin, activate the HIV-1 promoter and favor phosphorylation of the RNA polymerase II C-terminal domain. J. Biol. Chem. 274, 16097–16106 (1999).
Bensaude, O. Inhibiting eukaryotic transcription which compound to choose? How to evaluate its activity? Transcription 2, 103–108 (2011).
Vekilov, P. G. Nucleation. Cryst. Growth Des. 10, 5007–5019 (2010).
Azzalin, C. M., Reichenbach, P., Khoriauli, L., Giulotto, E. & Lingner, J. Telomeric repeat-containing RNA and RNA surveillance factors at mammalian chromosome ends. Science 318, 798–801 (2007).
Wu, B., Chao, J. A. & Singer, R. H. Fluorescence fluctuation spectroscopy enables quantitative imaging of single mRNAs in living cells. Biophys. J. 102, 2936–2944 (2012).
Fukaya, T., Lim, B. & Levine, M. Enhancer control of transcriptional bursting. Cell 166, 358–368 (2016).
Lee, C. F. & Wurtz, J. D. Novel physics arising from phase transitions in biology. J. Phys. D 52, 023001 (2018).
Berry, J., Brangwynne, C. P. & Haataja, M. Physical principles of intracellular organization via active and passive phase transitions. Rep. Prog. Phys. 81, 046601 (2018).
Wei, M.-T., Jedlicka, S. S. & Ou-Yang, H. D. Intracellular nonequilibrium fluctuating stresses indicate how nonlinear cellular mechanical properties adapt to microenvironmental rigidity. Sci. Rep. 10, 5902 (2020).
Söding, J., Zwicker, D., Sohrabi-Jahromi, S., Boehning, M. & Kirschbaum, J. Mechanisms for active regulation of biomolecular condensates. Trends Cell Biol. 30, 4–14 (2019).
Yao, R.-W. et al. Nascent pre-rRNA sorting via phase separation drives the assembly of dense fibrillar components in the human nucleolus. Mol. Cell 76, 767–783 (2019).
Shin, Y. et al. Liquid nuclear condensates mechanically sense and restructure the genome. Cell 175, 1481–1491 (2018).
Kim, N. Y. et al. Optogenetic control of mRNA localization and translation in live cells. Nat. Cell Biol. 22, 341–352 (2020).
Muller, C. B., Eckert, T., Loman, A., Enderlein, J. & Richtering, W. Dual-focus fluorescence correlation spectroscopy: a robust tool for studying molecular crowding. Soft Matter 5, 1358–1366 (2009).
Aaron, J. S., Taylor, A. B. & Chew, T. L. Image co-localization—co-occurrence versus correlation. J. Cell Sci. 131, 1–10 (2018).
Acknowledgements
We thank M. Levine and M. Haataja, as well as J. Eeftens and other members of the Brangwynne laboratory for helpful discussions and comments on this manuscript. This work was supported by the Howard Hughes Medical Institute, and grants from the National Institutes of Health 4D Nucleome Program (U01 DA040601), the Princeton Center for Complex Materials, a National Science Foundation supported MRSEC (DMR 1420541), as well an NSF CAREER award (1253035).
Author information
Authors and Affiliations
Contributions
M.-T.W. and C.P.B. designed research; M.-T.W., Y.-C.C. and Y.S. created constructs and performed experiments; M.-T.W., Y.-C.C, S.F.S. and A.R.S. analysed data; M.-T.W. and C.P.B. wrote the paper; and all authors reviewed and edited the paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
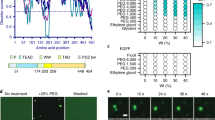
Extended Data Fig. 1 The amino acid sequence of intrinsically disordered regions.
Polar residues are shown in green; positively charged are shown in blue; negatively charged are shown in red; hydrophobic are shown in black; aromatic residues are shown in orange.
Extended Data Fig. 2 Statistical tests of the preferential condensation in nucleoplasm or cytoplasm.
a, OptoIDRs concentration outside clusters from individual cells under blue-light-illumination. Solid and open symbols represent C*Cyto and C*Nuc, respectively. The lines indicate the mean and error bars are s.d.s derived from the number of cells (as indicated in the panel) collected from 3 independent experiments. b, The ratio of C*Cyto and C*Nuc along with statistical tests to demonstrate that optoTAF15 has a preferred phase separate in the nucleus as compared to the cytoplasm. Dash line represents that C*Cyto equals to C*Nuc. In the optoTAF15 charged mutation constructs, the ratio C*Cyto over C*Nuc decreases as charged amino acids in optoTAF15 are mutated. Furthermore, the ratio maintains the same level for aromatic group mutation. These data suggest that charge-mediated interactions would contribute to TAF15’s preferential condensation in the nucleoplasm compared within cytoplasm. Data are plotted as calculated ratio of mean ± s.d. derived from the number of cells (as indicated in the panel a). c, Phase behaviors show strong dependence on protein sequence for phenylalanine mutation (Phenylalanine, F to Alanine, A). Steady-state cytoplasmic optoIDR concentration outside clusters (CCyto) from individual cells under blue light illumination as a function of optoIDR initial concentration (C0_Cyto). Solid and open symbols represent individual cells with or without light-activated assemblies respectively; solid line has slope of 1 and y-intercept of 0. d, The molecular interactions, β, along with statistical tests to demonstrate that optoTAF15 has a significantly stronger attractive forces to drive phase separate in the nucleus as compared to the cytoplasm, while optoFUS and optoEWS have little to no significant difference between cytoplasm and nucleus. Data are plotted as calculated ratio of mean ± s.d. derived from 20 of cells. P values are determined statistical significance obtained in pairwise two-sided Fisher’s exact tests in (a, b, and d). Statistical source data are provided in Source Data Extended Data Fig. 2.
Extended Data Fig. 3 Determinant of molecular interactions and inhabitation of transcription kinesis.
a, The molecular interactions, β, of the different constructs. Data are plotted as mean ± s.d. (n = 20 cells collected from 3 independent experiments). b, To assume the interactions are additive, the IDR interaction would be defined by subtracting the contribution of the construct of mCherry tagged Cry2 (βmch::Cry2) from the interaction strength (β) for optoFUS. To compare the contribution from FUSIDR and Cry2, the molecular interactions were systematically analyzed from the different constructs. The data showed that, the attractive forces from FUSIDR are around three-fold higher than the forces from Cry2 in the absence of blue light illumination. c, Fluorescent images of ATP depleted-cells expressing optoIDRs and eGFP-CTD after blue-light illumination. Cells were treated for 30 min in 2 mM sodium azide and 10 mM deoxyglucose to deplete ATP and inhibit phosphorylation. Cell nucleus is outlined by dotted line. Arrowheads point to optoIDR condensates colocalized with recruit CTD. Scale bar same for all images. d, CTD partition coefficients in various optoIDR condensates. These data suggest that each member of the FET family is capable of recruiting unphosphorylated CTD, while an unrelated protein cannot and indicates that EWS is indeed able to recruit CTD. The lines indicate the mean and error bars are s.d.s derived from the number of cells (as indicated in the panel) collected from 3 independent experiments. P values are determined statistical significance obtained in pairwise two-sided Fisher’s exact tests. e, THZ1 treatment to decrease the nucleoplasmic saturation concentration of optoTAF15. Nucleoplasmic saturation concentration as function of time over 3 hours. Black and red points represent untreated and THZ1-treated-cells, respectively. Data are plotted as mean ± s.d. (n = 15 cells collected from 3 independent experiments). Statistical source data are provided in Source Data Extended Data Fig. 3.
Extended Data Fig. 4 Repeated assemble and disassemble of optoIDRs condensates.
a, OptoDDX4 and TRF1-CTD cells, b, Actinomycin D (Act D) and α-amanitin treated optoTAF15 cells, and c, optoTAF15 and TRF1-CTD cells were exposed to blue light activation condition for 1 min to assemble clusters and then incubated in the absence of blue light for 9 min to disassemble clusters. Cell images before and after activation at the end of each cycle are shown. Experiment was repeated independently 3 times with similar results. d, Colocalization for TRF1-CTD various optoIDRs. Colocalization correlation obtained from Pearson’s Correlation results of nuclear pixel intensity (nucleoli excluded) for cells as shown in (a) and (c). Pearson Correlation Coefficient in each frames of successive cycles of activation shows higher colocalization function in optoTAF15 than optoDDX4. Additionally, colocalization correlation of optoIDRs as a function the lag cycle also shows higher colocalization function in optoTAF15 than optoDDX4. Those date indicate that optoTAF15 condensates specifically seed at these tethered loci, TRF1-CTD, and maintain repetitive localization in 8 successive cycles of activation, while optoDDX4 condensates, a control protein, do not seed at TRF1-CTD loci. e, OptoTAF15 and TRF1-CTD cells were exposed to continuous blue light illumination. OptoTAF15 condensates remain stable at telomere tethered CTD sites Experiment was repeated independently 3 times with similar result. Scale bar,, 5μm. Statistical source data are provided in Source Data Extended Data Fig. 4.
Extended Data Fig. 5 Immunofluorescence images of endogenous proteins and image of cell with nascent RNA and with telomeric repeat-containing RNA.
a, In the absence of blue light illumination, optoTAF15 cells were fixed using 4% paraformaldehyde. Then, the Immunofluorescence images were observed under super-resolution Airyscan microscopy. Cell nucleus is outlined by dotted line. Experiment was repeated independently 3 times with similar result. The scale bar is the same for all images. b, In cells expressing optoTAF15 and TRF1-CTD and kept in dark, the EU signal was distributed evenly throughout the nucleoplasm. Image of cell with nascent RNA production labeled by EU incorporation. In cells expressing optoTAF15 and TRF1-CTD under blue light illumination, nascent RNA transcripts were present in optoTAF15 condensates. White arrows indicate light-illuminated TAF15 condensates colocalize TRF1-CTD clusters and enhance local transcription RNA EU clusters. Experiment was repeated independently 3 times with similar result. c, Colocalization for TRF1-CTD with nascent RNA optoTAF15. Degree of colocalization is measured by Pearson’s correlation coefficient of nuclear pixel intensity (nucleoli excluded) between nascent RNA EU versus TRF1-CTD and optoTAF15 versus TRF1-CTD channels; +1 indicates perfect correlation, 0 no correlation, and -1 perfect anti-correlation. Colocalization results indicate that light-illuminated optoTAF15 condensates strongly colocalize with local nascent RNA EU production at TRF1-CTD clusters. Experiment was repeated independently 3 times with similar result. d, White arrows indicate light-illuminated optoTAF15 condensates colocalize TRF1-CTD clusters and enhance local transcription TERRA labeled by RNA fluorescence in situ hybridization at telomeres. e, Colocalization results indicate that light-illuminated optoTAF15 condensates enhance colocalization with transcription TERRA at TRF1-CTD clusters. Experiment was repeated independently 3 times with similar result. Statistical source data are provided in Source Data Extended Data Fig. 5.
Supplementary information
Source data
Source Data Fig. 1
Statistical source data
Source Data Fig. 2
Statistical source data
Source Data Fig. 3
Statistical source data
Source Data Fig. 4
Statistical source data
Source Data Fig. 5
Statistical source data
Source Data Extended Data Fig. 2
Statistical source data
Source Data Extended Data Fig. 3
Statistical source data
Source Data Extended Data Fig. 4
Statistical source data
Source Data Extended Data Fig. 5
Statistical source data
Rights and permissions
About this article

Cite this article
Wei, MT., Chang, YC., Shimobayashi, S.F. et al. Nucleated transcriptional condensates amplify gene expression. Nat Cell Biol 22, 1187–1196 (2020). https://doi.org/10.1038/s41556-020-00578-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41556-020-00578-6
This article is cited by
-
Transcriptional condensates: a blessing or a curse for gene regulation?
Communications Biology (2024)
-
Disordered C-terminal domain drives spatiotemporal confinement of RNAPII to enhance search for chromatin targets
Nature Cell Biology (2024)
-
Heterotypic interactions can drive selective co-condensation of prion-like low-complexity domains of FET proteins and mammalian SWI/SNF complex
Nature Communications (2024)
-
Transcription bodies regulate gene expression by sequestering CDK9
Nature Cell Biology (2024)
-
Asymmetric oligomerization state and sequence patterning can tune multiphase condensate miscibility
Nature Chemistry (2024)
