
Abstract
Somatic cell reprogramming by exogenous factors requires cooperation with transcriptional co-activators and co-repressors to effectively remodel the epigenetic environment. How this interplay is regulated remains poorly understood. Here, we demonstrate that NCoR/SMRT co-repressors bind to pluripotency loci to create a barrier to reprogramming with the four Yamanaka factors (OCT4, SOX2, KLF4 and c-MYC), and consequently, suppressing NCoR/SMRT significantly enhances reprogramming efficiency and kinetics. The core epigenetic subunit of the NCoR/SMRT complex, histone deacetylase 3 (HDAC3), contributes to the effects of NCoR/SMRT by inducing histone deacetylation at pluripotency loci. Among the Yamanaka factors, recruitment of NCoR/SMRT–HDAC3 to genomic loci is mostly facilitated by c-MYC. Hence, we describe how c-MYC is beneficial for the early phase of reprogramming but deleterious later. Overall, we uncover a role for NCoR/SMRT co-repressors in reprogramming and propose a dual function for c-MYC in this process.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout








Similar content being viewed by others

Change history
15 June 2018
In the version of this Article originally published, in Fig. 2c, the ‘+’ sign and ‘OSKM’ were superimposed in the label ‘+OSKM’. In Fig. 4e, in the labels, all instances of ‘Ant’ should have been ‘Anti-’. And, in Fig. 7a, the label ‘0.0’ was misplaced; it should have been on the colour scale bar. These figures have now been corrected in the online versions.
References
Takahashi, K. & Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 126, 663–676 (2006).
Apostolou, E. & Hochedlinger, K. Chromatin dynamics during cellular reprogramming. Nature 502, 462–471 (2013).
Chronis, C. et al. Cooperative binding of transcription factors orchestrates reprogramming. Cell 168, 442–459 (2017).
Soufi, A., Donahue, G. & Zaret, K. S. Facilitators and impediments of the pluripotency reprogramming factors’ initial engagement with the genome. Cell 151, 994–1004 (2012).
Cheloufi, S. et al. The histone chaperone CAF-1 safeguards somatic cell identity. Nature 528, 218–224 (2015).
Mansour, A. A. et al. The H3K27 demethylase Utx regulates somatic and germ cell epigenetic reprogramming. Nature 488, 409–413 (2012).
Singhal, N. et al. Chromatin-remodeling components of the BAF complex facilitate reprogramming. Cell 141, 943–955 (2010).
Di Stefano, B. et al. C/EBPα poises B cells for rapid reprogramming into induced pluripotent stem cells. Nature 506, 235–239 (2014).
Xu, Y. et al. Transcriptional control of somatic cell reprogramming. Trends Cell Biol. 26, 272–288 (2016).
Perissi, V., Jepsen, K., Glass, C. K. & Rosenfeld, M. G. Deconstructing repression: evolving models of co-repressor action. Nat. Rev. Genet. 11, 109–123 (2010).
Mottis, A., Mouchiroud, L. & Auwerx, J. Emerging roles of the corepressors NCoR1 and SMRT in homeostasis. Genes Dev. 27, 819–835 (2013).
Szabó, P. E., Hübner, K., Schöler, H. & Mann, J. R. Allele-specific expression of imprinted genes in mouse migratory primordial germ cells. Mech. Dev. 115, 157–160 (2002).
Wang, Y. et al. Reprogramming of mouse and human somatic cells by high-performance engineered factors. EMBO Rep. 12, 373–378 (2011).
Pijnappel, W. W. et al. A central role for TFIID in the pluripotent transcription circuitry. Nature 495, 516–519 (2013).
Di Stefano, B. et al. C/EBPα creates elite cells for iPSC reprogramming by upregulating Klf4 and increasing the levels of Lsd1 and Brd4. Nat. Cell Biol. 18, 371–381 (2016).
Esteban, M. A. et al. Vitamin C enhances the generation of mouse and human induced pluripotent stem cells. Cell Stem Cell 6, 71–79 (2010).
Gao, Y. et al. Replacement of Oct4 by Tet1 during iPSC induction reveals an important role of DNA methylation and hydroxymethylation in reprogramming. Cell Stem Cell 12, 453–469 (2013).
Stadtfeld, M., Maherali, N., Borkent, M. & Hochedlinger, K. A reprogrammable mouse strain from gene-targeted embryonic stem cells. Nat. Methods 7, 53–55 (2010).
Hanna, J. et al. Direct cell reprogramming is a stochastic process amenable to acceleration. Nature 462, 595–601 (2009).
Blelloch, R., Venere, M., Yen, J. & Ramalho-Santos, M. Generation of induced pluripotent stem cells in the absence of drug selection. Cell Stem Cell 1, 245–247 (2007).
Chen, J. et al. Rational optimization of reprogramming culture conditions for the generation of induced pluripotent stem cells with ultra-high efficiency and fast kinetics. Cell Res. 21, 884–894 (2011).
Samavarchi-Tehrani, P. et al. Functional genomics reveals a BMP-driven mesenchymal-to-epithelial transition in the initiation of somatic cell reprogramming. Cell Stem Cell 7, 64–77 (2010).
Li, R. et al. A mesenchymal-to-epithelial transition initiates and is required for the nuclear reprogramming of mouse fibroblasts. Cell Stem Cell 7, 51–63 (2010).
Polo, J. M. et al. A molecular roadmap of reprogramming somatic cells into iPS cells. Cell 151, 1617–1632 (2012).
Stender, J. D. et al. Control of proinflammatory gene programs by regulated trimethylation and demethylation of histone H4K20. Mol. Cell 48, 28–38 (2012).
Huangfu, D. et al. Induction of pluripotent stem cells by defined factors is greatly improved by small-molecule compounds. Nat. Biotechnol. 26, 795–797 (2008).
Guenther, M. G., Barak, O. & Lazar, M. A. The SMRT and N-cor corepressors are activating cofactors for histone deacetylase 3. Mol. Cell Biol. 21, 6091–6101 (2001).
Sun, Z. et al. Deacetylase-independent function of HDAC3 in transcription and metabolism requires nuclear receptor corepressor. Mol. Cell 52, 769–782 (2013).
Zhuang, Q. et al. Class IIa histone deacetylases and myocyte enhancer factor 2 proteins regulate the mesenchymal-to-epithelial transition of somatic cell reprogramming. J. Biol. Chem. 288, 12022–12031 (2013).
Lahm, A. et al. Unraveling the hidden catalytic activity of vertebrate class IIa histone deacetylases. Proc. Natl Acad. Sci. USA 104, 17335–17340 (2007).
Gaspar-Maia, A., Alajem, A., Meshorer, E. & Ramalho-Santos, M. Open chromatin in pluripotency and reprogramming. Nat. Rev. Mol. Cell Biol. 12, 36–47 (2011).
Hatzi, K. et al. A hybrid mechanism of action for BCL6 in B cells defined by formation of functionally distinct complexes at enhancers and promoters. Cell Rep. 4, 578–588 (2013).
Bhaskara, S. et al. Hdac3 is essential for the maintenance of chromatin structure and genome stability. Cancer Cell 18, 436–447 (2010).
Bogani, D. et al. The PR/SET domain zinc finger protein Prdm4 regulates gene expression in embryonic stem cells but plays a nonessential role in the developing mouse embryo. Mol. Cell Biol. 33, 3936–3950 (2013).
Liang, J. et al. Nanog and Oct4 associate with unique transcriptional repression complexes in embryonic stem cells. Nat. Cell Biol. 10, 731–739 (2008).
Saunders, A. et al. The SIN3A/HDAC corepressor complex functionally cooperates with NANOG to promote pluripotency. Cell Rep. 18, 1713–1726 (2017).
Wang, A. et al. Epigenetic priming of enhancers predicts developmental competence of hESC-derived endodermal lineage intermediates. Cell Stem Cell 16, 386–399 (2015).
Barish, G. D. et al. The Bcl6-SMRT/NCoR cistrome represses inflammation to attenuate atherosclerosis. Cell Metab. 15, 554–562 (2012).
Chen, J. et al. H3K9 methylation is a barrier during somatic cell reprogramming into iPSCs. Nat. Genet. 45, 34–42 (2013).
Sridharan, R. et al. Role of the murine reprogramming factors in the induction of pluripotency. Cell 136, 364–377 (2009).
Silva, J. et al. Promotion of reprogramming to ground state pluripotency by signal inhibition. PLoS Biol. 6, e253 (2008).
Chew, J. L. et al. Reciprocal transcriptional regulation of Pou5f1 and Sox2 via the Oct4/Sox2 complex in embryonic stem cells. Mol. Cell Biol. 25, 6031–6046 (2005).
Wernig, M., Meissner, A., Cassady, J. P. & Jaenisch, R. c-Myc is dispensable for direct reprogramming of mouse fibroblasts. Cell Stem Cell 2, 10–12 (2008).
Nakagawa, M. et al. Generation of induced pluripotent stem cells without Myc from mouse and human fibroblasts. Nat. Biotechnol. 26, 101–106 (2008).
Haberland, M., Montgomery, R. L. & Olson, E. N. The many roles of histone deacetylases in development and physiology: implications for disease and therapy. Nat. Rev. Genet. 10, 32–42 (2009).
Hirai, H., Tani, T. & Kikyo, N. Structure and functions of powerful transactivators: VP16, MyoD and FoxA. Int. J. Dev. Biol. 54, 1589–1596 (2010).
Soufi, A. et al. Pioneer transcription factors target partial DNA motifs on nucleosomes to initiate reprogramming. Cell 161, 555–568 (2015).
Liang, G., Taranova, O., Xia, K. & Zhang, Y. Butyrate promotes induced pluripotent stem cell generation. J. Biol. Chem. 285, 25516–25521 (2010).
Tonge, P. D. et al. Divergent reprogramming routes lead to alternative stem-cell states. Nature 516, 192–197 (2014).
Okita, K., Ichisaka, T. & Yamanaka, S. Generation of germline-competent induced pluripotent stem cells. Nature 448, 313–317 (2007).
Buganim, Y. et al. Single-cell expression analyses during cellular reprogramming reveal an early stochastic and a late hierarchic phase. Cell 150, 1209–1222 (2012).
Panopoulos, A. D. et al. Aberrant DNA methylation in human iPSCs associates with MYC-binding motifs in a clone-specific manner independent of genetics. Cell Stem Cell 20, 505–517 (2017).
Rais, Y. et al. Deterministic direct reprogramming of somatic cells to pluripotency. Nature 502, 65–70 (2013).
Hnisz, D. et al. Super-enhancers in the control of cell identity and disease. Cell 155, 934–947 (2013).
Yeom, Y. I. et al. Germline regulatory element of Oct-4 specific for the totipotent cycle of embryonal cells. Development 122, 881–894 (1996).
Grez, M., Akgun, E., Hilbergt, F. & Ostertag, W. Embryonic stem cell virus, a recombinant murine retrovirus with expression in embryonic stem cells. Proc. Natl Acad. Sci. USA 87, 9202–9206 (1990).
Raghav, S. K. et al. Integrative genomics identifies the corepressor SMRT as a gatekeeper of adipogenesis through the transcription factors C/EBPβ and KAISO. Mol. Cell 46, 335–350 (2012).
Langmead, B. & Salzberg, S. L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 9, 357–359 (2012).
Li, B.,& Dewey, C. N. RSEM: accurate transcript quantification from RNA-seq data with or without a reference genome. BMC Bioinformatics 12, 323 (2011).
Risso, D., Schwartz, K., Sherlock, G. & Dudoit, S. GC-content normalization for RNA-seq data. BMC Bioinformatics 12, 480 (2011).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550 (2014).
Zhang, Y. et al. Model-based analysis of ChIP-seq (MACS). Genome Biol. 9, R137 (2008).
Kumar, V. et al. Uniform, optimal signal processing of mapped deep-sequencing data. Nat. Biotechnol. 31, 615–622 (2013).
Heinz, S. et al. Simple combinations of lineage-determining transcription factors prime cis-regulatory elements required for macrophage and B cell identities. Mol. Cell 38, 576–589 (2010).
Hutchins, A. P., Jauch, R., Dyla, M. & Miranda-Saavedra, D. glbase: a framework for combining, analyzing and displaying heterogeneous genomic and high-throughput sequencing data. Cell Regen. 3, 1 (2014).
Young, M. D., Wakefield, M. J., Smyth, G. K. & Oshlack, A. Gene Ontology analysis for RNA-seq: accounting for selection bias. Genome Biol. 11, R14 (2010).
Bao, X. et al. The p53-induced lincRNA-p21 derails somatic cell reprogramming by sustaining H3K9me3 and CpG methylation at pluripotency gene promoters. Cell Res. 25, 80–92 (2015).
Shen, Y. et al. A map of the cis-regulatory sequences in the mouse genome. Nature 488, 116–120 (2012).
Whyte, W. A. et al. Master transcription factors and mediator establish super-enhancers at key cell identity genes. Cell 153, 307–319 (2013).
Guttman, M. et al. Ab initio reconstruction of cell type-specific transcriptomes in mouse reveals the conserved multi-exonic structure of lincRNAs. Nat. Biotechnol. 28, 503–510 (2010).
Acknowledgements
We thank all members of the Laboratory of RNA, Chromatin, and Human Disease for their comments, and Duanqing Pei and Jiekai Chen for their support, advice and/or technical assistance. We also thank J. Auwerx and R. M. Evans for providing materials. This work was supported by the National Key Research and Development Program of China (2016YFA0100102 and 2016YFA0100300), the Strategic Priority Research Program of the Chinese Academy of Sciences (XDA16030502), the National Natural Science Foundation of China (31371513, 31471242, 81571238, 31671537, 31370995 and 31571524), the National Natural Science Foundation of China–Israel Science Foundation Joint Research Scheme (31461143011), the National Natural Science Foundation of China/Research Grants Council Joint Research Scheme (81261160506), the Guangdong Province Science and Technology Program (2013B050800010, 2014A030312001, 2014A030310327, 2016B030229007, 2016A050503037 and 2015A030308007), the Guangzhou Science and Technology Program (201508030027), the Queensland–Chinese Academy of Sciences (Q-CAS) project (GJHZ1242), the Shenzhen Science and Technology Council Basic Research program (JCYJ20150331142757383 and JCYJ20170307110638890) and the Chinese Academy of Sciences-Croucher Funding Scheme for Joint Laboratories (CAS16HU01/CAS16902). The Guangdong Provincial Key Laboratory of Stem Cell and Regenerative Medicine is supported by the Science and Technology Planning Project of Guangdong Province, China (2017B030314056). C.W. is supported by Zhujiang Talents-Overseas Young Talents Postdoctoral Fellowship, M.J.K. and S.K. are supported by the CAS President’s International Fellowships, and D.P.I and M.M.A. are supported by the CAS-TWAS President’s PhD Fellowship.
Author information
Authors and Affiliations
Contributions
M.A.E., Q.Z. and C.B. conceived the original idea, and W.L. and A.P.H. contributed to the idea. Q.Z., W.L., C.B. and Z.H. conducted most of the experiments. A.P.H. and Z.Y. conducted all the bioinformatics analysis. Q.Z., W.L., C.B., A.P.H. and M.A.E. analysed the data. T.A., P.L., X.G., D.P.I., Z.L., M.Z., M.M.A., Z.Y., J.Y., Y.H., H.Z., D.H., J.Z., X.Zhong, X.Zhu, X.F., W.F., Y.L., Y.X., C.W., M.J.K. and S.K. contributed to the experiments and/or the analysis. B.M., M.D.T., H.-F.T., J.C., B.Q., X.B. and S.G. provided relevant advice and/or infrastructural support. M.A.E. supervised the study, with help from A.P.H., and provided most of the financial support. M.A.E. and A.P.H. wrote the manuscript with help from W.L.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Integrated supplementary information
Supplementary Figure 1 NCoR/SMRT co-repressors regulate OSKM reprogramming.
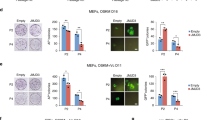
(a) RT-qPCR for Ncor1 and Ncor2 in MEFs transduced with OSKM in serum at different time points and in OG2 ESCs. Data are the mean ± s.e.m. (n=3 biological replicates with 3 technical replicates each) relative to MEFs. (b) Schematic of the Oct4-GFP reporter cassette in OG2 MEFs. The GFP is cloned straight after the ATG of Oct4 CDS and contains an SV40 polyA signal. DE=distal enhancer (deleted in this reporter) and PE=proximal enhancer. The numbers indicate the location of primers. (c) Genomic DNA qPCR using the primers described in panel b. The qPCR suggests that there are around 15 copies of the reporter in OG2 MEFs, whereas the ICR MEFs, which harbor no Oct4-GFP reporter, show no amplification. Data are the mean ± s.e.m. (n=3 biological replicates with 3 technical replicates each). (d) RT-qPCR analysis for detecting the Ncor1/2 knockdown efficiency in MEFs. Data are the mean ± s.e.m. (n=3 biological replicates with 3 technical replicates each) relative to shLuc control. (e) Phase contrast, Oct4-GFP, and immunofluorescence for NANOG for representative OSKM reprogramming experiments in serum+Vc on day 13 after Ncor1/2 knockdown in OG2 MEFs. Scale bar, 100 μm. (f, g) Time course of the appearance of NANOG+ colonies in OG2 MEFs (panel f) and ICR MEFs (with no Oct4-GFP reporter; panel g) transduced with OSKM after Ncor1/2 knockdown compared to shLuc, as measured by immunofluorescence. Data are the mean ± s.e.m. (n=3 biological replicates with 3 replicate items each), P value was calculated using two-tailed unpaired Student’s t-test. (h) As in panel e, but for ICR MEFs transduced with OSKM and shRNAs against Luciferase, Ncor1, Ncor2, or combined Ncor1 and Ncor2. Scale bar, 100 μm. (i, j) Number of GFP+ colonies on day 16 from secondary OG2 MEFs (panel i) or secondary MEFs with a GFP reporter knocked into the endogenous Oct4 locus (panel j), induced to reprogram with OSKM transgenes (left) or doxycycline (Dox, right) in serum+Vc; shRNAs against Luciferase, Ncor1, or Ncor2 were co-transduced. Data are the mean ± s.e.m. (n=3 biological replicates with 3 technical replicates each), P value was calculated using two-tailed unpaired Student’s t-test, ** indicates P value <0.01. (k) Time course of the number of cells in culture in MEFs transduced with OSKM and shRNAs against Luciferase, Ncor1, Ncor2, or combined Ncor1 and Ncor2 in serum+Vc. Data are the mean ± s.e.m. (n=3 biological replicates with 2 technical replicates each), P value was calculated using two-tailed unpaired Student’s t-test. (l) Representative flow cytometry plots of Oct4-GFP+ cells measured in MEFs transduced with OSKM and shRNAs against Luciferase or combined Ncor1 and Ncor2 from day 4 through day 14 of a reprogramming time course in KSR. (m) Number of GFP+ colonies on day 9 of MEFs transduced with OSK or OSKM and shRNAs against Luciferase, Ncor1, Ncor2, or combined Ncor1 and Ncor2 in iCD1. Data are the mean of 3 technical replicates from 1 of 2 representative experiments.
Supplementary Figure 2 Validation of iPSCs produced with Ncor1/2 knockdown.
(a) Phase contrast and Oct4-GFP images and immunofluorescence staining for the indicated pluripotency markers (red) for iPSC clones derived from MEFs transduced with OSKM and shRNAs against Ncor1 or Ncor2 in serum+Vc (also for all subsequent panels). Scale bars, 100 μm (phase contrast and GFP fluorescence image) and 50 μm (immunofluorescence). (b) PCR showing integration of the exogenous transgenes in the genome of the indicated iPSC clones, OG2 ESCs were included as control. (c) RT-qPCR showing silencing of exogenous transgenes in the indicated iPSC clones. Data are the mean of 2 technical replicates from 1 experiment relative to MEFs transduced with OSKM on day 6. (d) RT-qPCR showing endogenous expression of the indicated pluripotency genes in the indicated iPSC clones. Data are the mean of 2 technical replicates from 1 experiment. (e) Karyotype analysis of the indicated iPSC clones. (f) Chimeric mice generated with the indicated iPSC clones.
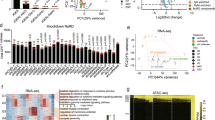
Supplementary Figure 3 RNA-seq analysis of depletion of NCoR/SMRT co-repressors in OSKM reprogramming.
(a) Heatmap showing all genes from the RNA-seq analysis of MEFs transduced with OSKM and shRNAs against Luciferase, Ncor1,and Ncor2. Fold change is relative to MEFs. MEF and ESC data are taken from GSE29278 (same for panels c, d, f). (b) GO analysis showing the up-regulated genes from panel a on day 5 (left panel) and day 9 (right panel). (c) Heatmaps of the genes significantly differentially up-regulated or down-regulated in response to Ncor1/2 knockdown. Genes significantly up- or down-regulated in either Ncor1 or Ncor2 shRNA experiments were merged. (d) Boxplots of the genes in panel c and Fig. 1e, including MEF and ESC data. In general, Ncor1 and Ncor2 up-regulated genes on day 5 of OSKM reprogramming are specific to MEFs. The number of genes is indicated by ‘n’, the boxplots indicate the upper and lower quartiles, the central line is the median, the whiskers indicate 1.5*interquartile range and the circles are outliers. (e) Heatmap showing the fold-change of selected MET and epithelial-to-mesenchymal transition (EMT)-related genes from the RNA-seq analysis. (f) ‘Second wave’ genes, which are associated with the later stage of reprogramming, were mapped onto the reprogramming RNA-seq data. Gene expression was converted into a gene-wise Z-score, and P values were from a two-tailed Mann-Whitney-U test. The number of genes is indicated by ‘n’, the boxplots indicate the upper and lower quartiles, the central line is the median, the whiskers indicate 1.5*interquartile range and the circles are outliers. The 95% confidence intervals for day 5 data: shLuc (−0.44, −0.32), shNcor1 (−0.48, −0.35), and shNcor2 (−0.47, −0.36). And for day 9 data: shLuc (−0.16, −0.01), shNcor1 (0.058, 0.21), and shNcor2 (0.039, 0.18). (g, h) RT-qPCR for the indicated somatic genes on day 5 (panel g) and pluripotency genes on day 9 (panel h) in MEFs transduced with OSKM and shRNAs against Luciferase, Ncor1, Ncor2, or combined Ncor1 and Ncor2 in serum+Vc. Data are the mean ± s.e.m. (n=3 biological replicates for somatic genes and n=4 for pluripotency genes with 3 technical replicates each), P value was calculated using two-tailed unpaired Student’s t-test, * indicates P value <0.05. (i) RT-qPCR for the indicated pluripotency genes on day 13 of MEFs transduced with OSKM and shRNAs against Luciferase, Ncor1, Ncor2, and combined Ncor1 and Ncor2 in serum+Vc, measured relative to OSKM plus shLuc. MEFs and E14 ESCs are included for comparison. Data are the mean ± s.e.m. (n=3 biological replicates with 3 technical replicates each), P value was calculated using two-tailed unpaired Student’s t-test, * indicates P value <0.05.
Supplementary Figure 4 HDAC screening and analysis of the role of HDAC3 in OSKM reprogramming.
(a) RT-qPCR analysis showing the knockdown efficiency of Hdac members in MEFs. Data are the mean ± s.e.m. (n=3 biological replicates with 3 technical replicates each) relative to shLuc control. (b) Immunoblotting with the indicated antibodies using lysates from MEFs transduced with OSKM and FLAG-tagged HDAC3, HDAC3-YF, and HDAC3-YF-KA on day 7 in serum+Vc. (c) Co-immunoprecipitations of FLAG-tagged NCoR/SMRT and HA-tagged HDAC3 in lysates from HEK293T cells. (d) Phase contrast and Oct4-GFP+ cells in OSKM reprogramming in serum+Vc at the indicated days. Scale bar, 100 μm. (e) Time course measurement of the appearance of Oct4-GFP+ colonies in MEFs transduced with OSKM and HDAC3-YF or empty vector in serum+Vc. Data are the mean ± s.e.m. (n=3 biological replicates with 3 technical replicates each), P value was calculated using two-tailed unpaired Student’s t-test. (f) Representative flow cytometry plots showing the appearance of Oct4-GFP+ cells in MEFs transduced with OSKM and HDAC3-YF or empty vector in KSR. (g) Measurements of cell numbers during a reprogramming time course in serum+Vc. Cells were transduced with OSKM and HDAC3-YF or empty vector. Data are the mean ± s.e.m. (n=3 biological replicates with 2 technical replicates each), P value was calculated using two-tailed unpaired Student’s t-test. (h) RT-qPCR detection of the indicated pluripotency genes in MEFs transduced with OSKM and HDAC3-YF or empty vector on day 13 in serum+Vc. MEFs and E14 ESCs are included as comparisons. Data are the mean ± s.e.m. (n=3 biological replicates with 3 technical replicates each), relative to OSKM and empty vector, P value was calculated using two-tailed unpaired Student’s t-test, * indicates P value <0.05. (i) RT-qPCR analysis showing the expression of the indicated MEF-enriched genes on day 5 in MEFs transduced with OSKM and HDAC3-YF or empty vector in serum+Vc (n=3 biological replicates with 3 technical replicates each), P value was calculated using two-tailed unpaired Student’s t-test. (j) Heatmap of all genes passing quality control from the single-cell RT-qPCR analysis organized into categories according to the type of gene; each column is a single cell. Data were calculated as the ACx (relative expression). (k) Principal component gene loading of the single-cell RT-qPCR data. (l, m) Violin plots of single-cell RT-qPCR expression of pluripotency genes (panel l) or the somatic gene S100a4 (panel m). The red bar represents the mean, and samples sizes are indicated in panel j.
Supplementary Figure 5 HDAC3 deacetylase-null mutant induces H3K27ac change at pluripotency loci in OSKM reprogramming.
(a) Radial plots indicating the genomic distribution of H3K27ac peaks in relation to the TSS. Each radial plot indicates the locations of H3K27ac on the indicated days, the left set of plots are MEFs transduced with OSKM and empty vector in serum+Vc, whereas the right set are MEFs with OSKM and HDAC3-YF. (b) Genome views of H3K27ac tag density at Nanog and Zic2 (note that the 2610035F20Rik gene beside Zic2 is an antisense long intergenic non-coding RNA that shares the TSS with Zic2) in cells transduced with OSKM and HDAC3-YF or empty vector in serum+Vc. (c) ChIP-qPCR analysis of H3K27ac at the Nanog and Zic2 promoters on days 5, 9, and 13 of MEFs tranduced with OSKM and HDAC3-YF or empty vector in serum+Vc. Data are the mean ± s.e.m. (n=3 biological replicates with 3 technical replicates each), P value was calculated using two-tailed unpaired Student’s t-test, ** indicates P value <0.01. (d) ChIP-qPCR for HDAC3-YF binding at Nanog and Zic2 promoters in MEFs transduced with OSKM and HDAC3-YF-FLAG or empty vector on day 9 in serum+Vc. Data are the mean ± s.e.m. (n=3 biological replicates with 3 technical replicates), P value was calculated using two-tailed unpaired Student’s t-test, * indicates P value <0.05. (e) RT-qPCR detection of the expression of Nanog and Zic2 on days 5, 9, and 13 of OSKM reprogramming in serum+Vc. Data are the mean ± s.e.m. (n=3 biological replicates with 3 technical replicates each). (f) Genome views of the level of H3K27ac at the indicated somatic loci (S100a4, Col6a1, and Acta2) in MEFs, OG2 ESCs, and MEFs transduced with OSKM and HDAC3-YF or empty vector in serum+Vc.
Supplementary Figure 6 NCoR and SMRT binding in ESCs and in reprogramming.
(a) Radial plots of the genome-wide distribution of NCoR or SMRT binding sites relative to the TSS. (b) GO analysis of groups 5, 7, and 15. The list of genes for GO was generated by taking all genes within 2 kb and the closest gene within 20 kb of the NCoR/SMRT binding sites and removing duplicate genes. (c) Genome views of NCoR and SMRT binding at Klf2. Black dots under the tracks indicate that NCoR or SMRT was detected as bound at that location, also for panel d. (d) NCoR and SMRT binding to the developmental-related and pro-differentiation genes Sox7 (endoderm), Tcf15 (mesoderm), and Pou4f2 (ectoderm). (e) Knockdown efficiency of shRNAs (pRetroSuper) against Ncor1 and Ncor2 in OG2 ESCs. Data are the mean ± s.e.m. (n=3 biological replicates with 3 technical replicates each) relative to shLuc control. (f) Morphology and Oct4-GFP expression in OG2 ESCs (passage 5 after transduction) transduced with shRNAs (pRetroSuper) against Luciferase, Ncor1, or Ncor2. Scale bar is 100 μm. (g) Knockdown efficiency of shRNAs (pLKO) against Ncor1 and Ncor2 in E14 ESCs 72 hours after transduction. Data are the mean of 3 technical replicates from 1 experiment relative to shLuc control. (h) RT-qPCR data for the expression of selected pluripotent genes and developmental-related genes after knockdown (pLKO) of Ncor1 or Ncor2 in E14 ESCs. Data are the mean of 3 technical replicates from 1 experiment relative to shLuc control.
Supplementary Figure 7 Genome loci bound by NCoR/SMRT show higher levels of H3K27ac when HDAC3-YF is included in a reprogramming assay and in ESCs.
(a) Violin plots for the normalized tag density within a 1 kb window of all H3K27ac peaks for each ChIP-seq experiment. The red bar is mean and the green bar is the median. Each violin contains all of the peaks for the indicated condition. The number of peaks in each violin (from left-to-right): 10887, 11005, 11050, 9901, 11159, 12525, and 20717. (b) Heatmaps showing NCoR/SMRT reprogramming-specific peaks (group 5), ESC-specific peaks (group 7), and NCoR/SMRT peaks common to both reprogramming and ESCs (group 15). Each row is an NCoR/SMRT binding site. Data were converted into a Z-score to emphasize relative changes, and the Z-score was generated from the standard deviation of all NCoR/SMRT binding sites. (c) Twenty-three selected ‘second wave’ genes with no evidence of NCoR/SMRT binding within 20 kb of the TSS were analyzed for the levels of H3K27ac on day 9 or day 13 of an OSKM reprogramming time course for cells transduced with HDAC3-YF or empty vector. Tag density of H3K27ac were measured in a 1 kb window centered on the TSS of the indicated gene, scores were converted to a Z-score based on the total variance of all peaks. (d) Genome view for the H3K27ac levels at the Gdf3 locus, a second wave gene, not bound by NCoR/SMRT within 20 kb of the TSS. (e) Violin plots for the normalized tag density of H3K27ac in ESC-specific NCoR/SMRT binding sites (group 7) for an OSKM reprogramming time course co-transduced with HDAC3-YF or empty vector. The green bar represents the median, n is the number of transcripts bound by NCoR/SMRT in ESCs, and the P value was calculated using two-tailed Mann-Whitney-U test.
Supplementary Figure 8 c-MYC cooperates with NCoR/SMRT to derail reprogramming and this can be overcome by conjugating the VP16 co-activator domain to OCT4 or SOX2.
(a) Heatmap showing the overlap of NCoR/SMRT bound sites (day 9 OSKM reprogramming) versus c-MYC (48 h OSKM reprogramming and pre-iPSCs). The heatmap was generated by creating a non-redundant list of overlapping peak summits within 400 bp. Each row of the heatmap is a genomic locus either bound (red) or not bound (white) by the indicated factor. Only sites bound by NCoR/SMRT (day 9 OSKM reprogramming) are shown. The number of sites bound by the different factors and time points is indicated. Data are taken from GSE90895, the same for panels b-e. (b) GO analysis of the ChIP-seq peaks co-bound by NCoR/SMRT and c-MYC in 48 h OSKM reprogramming or pre-iPSCs (panel a). Gene sets for GO analysis were collected by annotating all genes within 2 kb and the closest gene within 20 kb of a binding site, duplicate genes were removed. (c) NCoR/SMRT and c-MYC are bound to the Klf2 locus during reprogramming and in ESCs. Genomic view of NCoR/SMRT and c-MYC binding during reprogramming (day 9 OSKM reprogramming, 48 h OSKM reprogramming and pre-iPSCs) and in ESCs. (d) Pileup tag density plots for NCoR/SMRT bound sites in day 9 OSKM reprogramming versus the binding sites for OCT4, SOX2, KLF4, and c-MYC in ESCs (left panel). Plots are centered on NCoR/SMRT binding sites, the tag density of the indicated OSKM factors is shown. Data are normalized to library size. Pair-wise Venn overlap for NCoR/SMRT bound sites in day 9 OSKM reprogramming versus OSKM bound sites in ESCs (right panel). Peaks were considered overlapping if their summits were within 400 bp. (e) As in panel d, but comparing NCoR/SMRT bound sites in ESCs to OSKM bound sites in ESCs. (f) Flow cytometry on day 6 after co-transducing the Oct4 distal enhancer-driven DsRed reporter in MEFs with empty vector, OSK or OSKM in serum+Vc. Percent indicates the proportion of cells positive for DsRed. (g) Number of Oct4-GFP+ colonies on day 16 in MEFs transduced with OSKM, treated with TSA across the indicated reprogramming windows in serum+Vc. Data are the mean of 3 technical replicates from 1 of 2 representative experiments. (h) Number of Oct4-GFP+ colonies on day 16 in MEFs transduced with OSKM and inducible HDAC3-YF treated with doxycycline across the indicated reprogramming windows in serum+Vc. Data are the mean ± s.e.m. (n=3 biological replicates with 3 technical replicates each), P value was calculated using one-tailed unpaired Student’s t-test. (i) Phase contrast and fluorescence images for detecting Oct4 distal enhancer-driven DsRed reporter in MEFs co-transduced with OSKM and the indicated engineered reprogramming factors OCT4-VP16 and SOX2-VP16 on day 6 in serum+Vc. Scale bar, 100 μm. (j) Number of GFP+ colonies on day 18 (serum) or day 16 (serum+Vc) in MEFs transduced with either full length versions of OSKM, or OVP16SKM, OSVP16KM, OVP16SVP16KM, where the VP16 domain is fused to the indicated reprogramming factor. Data are the mean ± s.e.m. (n=3 biological replicates with 3 technical replicates each), P value was calculated using two-tailed unpaired Student’s t-test, ** indicates P value <0.01. (k) Schematic representing how c-MYC directs the NCoR/SMRT-HDAC3 co-repressor complex to conflicting purposes during reprogramming. At somatic loci, recruitment of NCoR/SMRT is beneficial in the earlier stages of reprogramming, as it seemingly works to suppress the somatic gene program, which may be independent of HDAC3. However, at pluripotency loci and potentially other loci, recruitment of NCoR/SMRT-HDAC3 is deleterious, working to restrain reprogramming.
Supplementary Figure 9
Unprocessed scans of blots for all figures in the manuscript.
Supplementary information
Supplementary Information
Supplementary Figures 1–9, Supplementary Table legends, and Supplementary References.
Supplementary Table 1
shNcor1/2-differentially regulated genes.
Supplementary Table 2
TaqMan primers used for single-cell RT–qPCR.
Supplementary Table 3
Acx values for all primers used in the single-cell RT–qPCR.
Supplementary Table 4
Nearest gene to NCoR/SMRT ChIP-seq peaks.
Supplementary Table 5
shRNA target sequences and PCR primers.
Supplementary Table 6
Statistics source data.
Rights and permissions
About this article

Cite this article
Zhuang, Q., Li, W., Benda, C. et al. NCoR/SMRT co-repressors cooperate with c-MYC to create an epigenetic barrier to somatic cell reprogramming. Nat Cell Biol 20, 400–412 (2018). https://doi.org/10.1038/s41556-018-0047-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41556-018-0047-x
This article is cited by
-
VGLL1 cooperates with TEAD4 to control human trophectoderm lineage specification
Nature Communications (2024)
-
A spatiotemporal atlas of mouse liver homeostasis and regeneration
Nature Genetics (2024)
-
Restricting epigenetic activity promotes the reprogramming of transformed cells to pluripotency in a line-specific manner
Cell Death Discovery (2023)
-
Oct4 cooperates with c-Myc to improve mesenchymal-to-endothelial transition and myocardial repair of cardiac-resident mesenchymal stem cells
Stem Cell Research & Therapy (2022)
-
Changes in chromatin accessibility landscape and histone H3 core acetylation during valproic acid-induced differentiation of embryonic stem cells
Epigenetics & Chromatin (2021)
