
Abstract
The success of messenger RNA therapeutics largely depends on the availability of delivery systems that enable the safe, effective and stable translation of genetic material into functional proteins. Here we show that extracellular vesicles (EVs) produced via cellular nanoporation from human dermal fibroblasts, and encapsulating mRNA encoding for extracellular-matrix α1 type-I collagen (COL1A1) induced the formation of collagen-protein grafts and reduced wrinkle formation in the collagen-depleted dermal tissue of mice with photoaged skin. We also show that the intradermal delivery of the mRNA-loaded EVs via a microneedle array led to the prolonged and more uniform synthesis and replacement of collagen in the dermis of the animals. The intradermal delivery of EV-based COL1A1 mRNA may make for an effective protein-replacement therapy for the treatment of photoaged skin.
Similar content being viewed by others

Main
Recent developments in messenger RNA-modification techniques have enhanced the therapeutic efficiency of mRNA delivery and its potential for near-term clinical applications, including protein-replacement therapy and vaccination against the severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) virus1,2. However, the intrinsic inability and potential immunogenicity of mRNAs require that they be encapsulated within delivery vehicles. Current mRNA-delivery modalities centre on the usage of lipid nanoparticle (LNP) carriers for encapsulation and transport3,4. However, LNPs pose several major challenges, including cytotoxicity, poor biodistribution, lack of target specificity and immunogenicity. These problems may be caused by the requirement for the surface PEGylation (PEG stands for poly(ethylene glycol)) of LNPs to improve their circulatory half-life and to reduce non-specific clearance5,6. Notably, the administration of LNPs in people has been linked to anaphylaxis, hypersensitivity and autoimmune adverse events7,8. Therefore, the identification of mRNA carriers that can overcome some of these LNP-associated challenges would be helpful for the further development of mRNA-based therapeutics.
Extracellular vesicles (EVs), including exosomes and microvesicles, play a major role in the transport of biomolecules and nucleic acids, including mRNAs, within the human body9,10,11. As a result, in recent years, EVs have emerged as promising carriers for nucleic-acid-based therapeutics owing to their intrinsic biocompatibility, their ability to cross physiological barriers and their low immunogenicity12,13. Unlike LNPs, EVs, including exosomes, are endogenously produced by the body’s cells and lead to lower levels of inflammatory responses. Moreover, strategies to cheaply and easily produce large quantities of exosomes have been developed. We previously reported a cellular nanoporation (CNP) method in which transient nanometric pores were created on the surface of source cells to allow for the large-scale loading of full-transcript mRNAs into secreted EVs14. Here, by using a mouse model of acute photoaging that closely mimics the pathophysiological features of aging-damaged skin in humans15, we show the utility of exosome-based COL1A1 mRNA therapy to replace dermal collagen-protein loss as an anti-aging treatment for photoaged skin. To improve the efficiency of mRNA delivery and retention, we also show that the delivery of collagen mRNA via a hyaluronic acid (HA) microneedle (COL1A1-EV MN) patch allows for a more efficient distribution of mRNA in the dermis, resulting in durable collagen-protein engraftment and in an improved treatment of wrinkles in photoaged skin.
Results
Preparation and in vitro delivery of COL1A1 mRNA-containing EVs
Dermal atrophy owing to irreversible loss of collagen is a hallmark of skin aging16,17. Numerous methods have aimed to restore loss of collagen protein in skin, ranging from over the counter and pharmaceutical approaches (antioxidants18,19,20, retinoids21, peptides22,23) to medical devices (that is, laser therapy24 and synthetic dermal fillers25,26). However, none of these existing technologies have been able to achieve long-term endogenous collagen replacement to maintain skin strength, firmness and elasticity over time27,28,29. Stimulating fibroblasts responsible for synthesizing collagen proteins can also be an effective way for short-term control of skin aging30. However, fibroblasts gradually lose their capacity to proliferate and synthesize collagen as they senesce, resulting in challenges for longer-term methods of collagen replacement for anti-aging treatment31. To overcome these limitations, we aimed to replace collagen protein in a photoaging collagen-depletion model via EV-mediated mRNA delivery. To generate EVs loaded with human collagen I alpha I (COL1A1) mRNA, we employed a CNP technique that involved plating a monolayer of neonatal human dermal fibroblasts (nHDFs) on a nanopore surface and nano-transfecting the cells with a COL1A1-GFP plasmid (Fig. 1a and Supplementary Fig. 1a)14. EVs were isolated from culture media the day after transfection. CNP-treated cells were found to have a 10-fold higher EV number per cell as compared with cells treated with standard bulk electroporation (BEP), which was performed using cuvette-style parallel electrodes as previously described32, or untreated nHDFs in culture (Fig. 1b). The EVs produced by each method were characterized by a size distribution peaking at about 150 nm in diameter as determined by nanoparticle tracking analysis (NTA) and 80% of intensity in the mode peak by dynamic light scattering (DLS) (Fig. 1c and Supplementary Fig. 1b,c) with a polydispersity index (PDI) of 0.12–0.25. Western blot experiments showed that expressions of exosome (CD9, CD63, TSG101) and microvesicle (ARF6) biomarkers in the CNP-treated group were significantly higher than in the untreated group, confirming the increase in secreted EVs (Supplementary Fig. 1d). Kinetic analyses further showed that voltage-optimized EV release peaked at 8 h after CNP induction, with continued secretion noted over the next 24 h (Supplementary Fig. 1e,f). Reverse transcription-quantitative polymerase chain reaction (RT–qPCR) showed that CNP-secreted EVs contained more than 200 times the COL1A1 mRNA than BEP-secreted EVs and 3,000-fold higher COL1A1 mRNA than EVs secreted from non-transfected cells (Fig. 1d). Bioanalyser assessment of gel agarose demonstrated full-length transcribed COL1A1 mRNA at ~4,000 nucleotides (Fig. 1e). EVs prepared by CNP exhibited structural stability when stored for preclinical administration at 4 °C with no changes in appearance, membrane and size properties when evaluated by cryo-electron microscopy (cryo-EM), atomic force microscopy (AFM) and NTA (Supplementary Fig. 2a–c). Furthermore, COL1A1 mRNA encapsulated within EVs was stable both at room temperature and at 4 °C, and also exhibited serum stability, thereby highlighting their potential for future clinical utility (Supplementary Fig. 2d,e).

a, Schematic representation of CNP-generated EVs for targeted nucleic acid delivery. b, EV number per cell produced by untreated nHDFs in PBS buffer as control, BEP or CNP with PBS buffer only in 24 h (n = 3 for each group; *P = 0.048 Control vs BEP; **P = 0.005 Control vs CNP; ##P = 0.002 BEP vs CNP). c, Characterization of EVs by nanoparticle tracking analysis. d, RT–qPCR of COL1A1 mRNA reveals that EVs produced by CNP contain much larger quantities of transcribed mRNAs than EVs produced by BEP and Control groups in 24 h (n = 3 for all groups; **P = 0.002 Control vs BEP; ***P < 0.001 Control vs CNP; ###P < 0.001 BEP vs CNP). e, Gel electrophoresis of total RNA extracted from 3.78 × 108 EVs produced from CNP using COL1A1-GFP plasmid compared with synthetic in vitro-transcribed COL1A1 mRNA as control. f, Proliferation of nHDFs after treatment with different protein concentrations of COL1A1-EVs at 0 h, 24 h and 48 h (n = 4 for all groups; at 24 h: #P = 0.0107, 10 µg ml−1 vs 0 µg ml−1 of protein concentration of COL1A1-EVs after treatment; ##P = 0.002, 20 µg ml−1 vs 0 µg ml−1; ###P < 0.001, 30 µg ml−1 vs 0 µg ml−1; at 48 h: *P = 0.045, 10 µg ml−1 vs 0 µg ml−1 of COL1A1-EV treatment; ***P < 0.001, 20 µg ml−1 vs 0 µg ml−1; ***P < 0.001, 30 µg ml−1 vs 0 µg ml−1). All data are from three independent experiments and are presented as means ± s.e.m. Two-sided Student’s t-tests were used for the comparisons in b and d. One-way ANOVA was used for the comparisons in f. The schematic in a was created with BioRender.com.
To assess the therapeutic potential of COL1A1 mRNA-containing EVs (COL1A1-EVs) in vitro, we treated cultured fibroblasts with COL1A1-EVs for 48 h (Supplementary Fig. 3a). Proliferation of fibroblasts was observed to increase with COL1A1-EV treatment in a dose-dependent manner (Fig. 1f). After treatment, elevated COL1A1 protein expression was observed in fibroblasts treated with COL1A1-EVs, as evidenced by increased co-localization of green fluorescent protein (GFP) with COL1A1 under immunofluorescence microscopy. In contrast, COL1A1 levels were substantially lower without co-localization of GFP in the non-treated group (Extended Data Fig. 1a,b). Successful mRNA delivery into recipient cells was further confirmed by elevated collagen mRNA expression and collagen protein levels measured via quantitative PCR and western blot analysis, respectively, in fibroblasts treated with COL1A1-EVs (Extended Data Fig. 1c,d and Supplementary Fig. 3b). Moreover, pro-collagen I, a precursor of COL1 protein, was significantly increased following COL1A1-EV treatment, indicating improved COL1 protein synthesis by the EV-treated fibroblasts compared with untreated fibroblasts (Extended Data Fig. 1e). Confocal microscopy imaging of cellular uptake of EVs indicated that EV cargo delivery into recipient cells is mediated by clathrin-mediated endocytosis (Supplementary Fig. 3c,d) and is capable of lysosome escape (Supplementary Fig. 4). Taken together, these findings demonstrate that functional COL1A1 mRNA can be stably encapsulated within EVs by CNP and that this COL1A1-EV delivery system can significantly increase COL1A1 protein expression in vitro.
In vivo kinetics of COL1A1 mRNA expression and protein translation following COL1A1-EV delivery
To understand the kinetics of EV-mediated COL1A1 mRNA delivery and protein formation in vivo, we delivered 2.7 × 109 copy number of COL1A1 mRNA COL1A1-EVs into the dermis of mice via an insulin needle syringe. The mice were killed over the next 14 d for histological analysis by RNAscope for COL1A1 mRNA quantification. COL1A1 mRNA was found to be significantly elevated in local skin tissue at 12 h after delivery, with notable decreases observed at 24 h and 48 h after injection, and a return to baseline COL1A1 mRNA levels by 96 h (Fig. 2a,b). In vivo translation of COL1A1 mRNA into protein was assessed by immunofluorescence microscopy of explanted tissue over a 30 d period. GFP+ COL1A1+ immunostained grafts were observed as early as 12 h after injection, with peak fluorescence at 4 d after delivery (Fig. 2c–e). GFP+ COL1A1+ protein grafts were observed to decrease in a time-dependent manner from day 4 to day 30 after injection as observed by immunofluorescence microscopy, with the majority of COL1A1-EV-derived protein turning over by day 30. These findings demonstrate that lower doses of COL1A1-EV delivery results in a 3–4 d peak of COL1A1 protein in recipient tissues, decreasing to baseline levels by day 30 after injection.

a, In situ hybridization of human COL1A1 mRNA by RNAscope after intradermal injection of COL1A1-EVs and saline control, measured at 12 h, 24 h, 48 h, 96 h, 7 d, 10 d and 14 d after injection. Scale bar,100 µm. b, Quantification of RNAscope results by average number of brown dots per cell. c, Immunofluorescence over time of COL1A1-EV-derived protein by visualization of co-localized GFP and COL1A1 protein (RFP). Scale bar, 100 µm. d, Fluorescence intensity quantification of COL1A1-GFP protein expression. e, Quantification of GFP+ cells confirms that the COL1A1-EV-derived protein grafts decrease in a time-dependent manner over 30 d. All data are from three independent experiments and are presented as means ± s.e.m.
In vivo therapeutic efficacy of COL1A1-EVs in a collagen-depletion skin-photoaging model
Dermal photoaging is characterized by the destruction of collagen and extracellular-matrix (ECM) proteins in the skin by sun exposure and UV irradiation, which causes the formation of wrinkles33,34. To assess the capacity of COL1A1 mRNA EVs to replace degraded collagen protein in vivo, we employed a previously described photoaging model in which athymic nude mice were treated with UV (311 nm) irradiation over an 8 week period, resulting in dermal wrinkles secondary to collagen depletion35 (Supplementary Fig. 5a). The efficacy of the 8 week UV irradiation course in degrading collagen and elastin fibres in murine skin was confirmed by histological and tissue analysis (Supplementary Fig. 5b–g). We next assessed the therapeutic potential of COL1A1-EVs to replace dermal collagen in UV-irradiated mice by treating the animals with a 5x injection course of either: (1) 2.7 × 109 copy number of COL1A1 mRNA encapsulated in EVs (COL1A1-EVs), (2) 2.7 × 109 copy number of COL1A1 mRNA encapsulated in lipid nanoparticles (COL1A1-LNPs), (3) control EVs without CNP cargo loading (unloaded EVs), (4) 0.05% retinoic acid topical treatment (RA) or (5) saline control. Wrinkle formation was tracked at days 0, 4, 7, 14, 21 and 28 after initiation of treatment, and all animals were killed at day 28 for histology and skin plaster replica analyses36 (Fig. 3a). Microscopic photography of dermal skin wrinkles over the treatment period demonstrated modest decreases in wrinkle number and area by day 28 for the COL1A1-LNPs, unloaded EVs and retinoic acid groups compared with the saline control group (Fig. 3b). In contrast, photoaged mice treated with COL1A1-EVs exhibited a reduction in wrinkle number and area beginning on day 7 after treatment initiation, with a significant reduction from day 14 onward to levels similar to those observed in unirradiated sham controls (Fig. 3c,d). Skin plaster replicas of the dorsal skin taken at day 28 after treatment initiation confirmed the effectiveness of COL1A1-EVs for treating photoaged skin as compared with saline control, retinoic acid, unloaded EV controls and COL1A1-LNPs (Extended Data Fig. 2). Noticeably, COL1A1-LNPs also exhibited a reduction in wrinkle number and area compared with saline control, retinoic acid and unloaded EV controls, although this effect was not as pronounced as with COL1A1-EVs. Skin treated with COL1A1-EVs and COL1A1-LNPs also demonstrated higher elasticity and firmness as measured by a cutometer and an Instron extensometer, respectively (Supplementary Fig. 6).

a, Schematic representation and timeline of 5 low-dose injections of COL1A1-EVs (2.7 × 109 copy number COL1A1 mRNA per dose). Skin tissue was collected at 28 d. b, Wrinkle formation was tracked at days 0, 4, 7, 14, 21 and 28 after initial delivery of COL1A1-EVs, COL1A1-LNPs, unloaded EVs, 0.05% retinoic acid (RA) or saline (n = 4 for all groups). The sham group comprised female nude mice that were not exposed to UV irradiation. Scale bar, 5 mm. c, Total numbers of dorsal-skin wrinkles quantified with custom software (n = 4 for all groups; ※※P = 0.0016 COL1A1-EVs vs Saline at day 7; ※※※P < 0.001 COL1A1-EVs vs Saline at days 14, 21 and 28; **P = 0.0089 COL1A1-LNPs vs Saline at day 21; **P = 0.0097 COL1A1-LNPs vs Saline at day 28; #P = 0.461 Unloaded EVs vs Saline at day 21; ##P = 0.0034 Unloaded EVs vs Saline at day 28; †P = 0.0142 RA vs Saline at day 21; ††P = 0.0069 RA vs Saline at day 28). d, Quantification of wrinkle area on dorsal skin (n = 4 for all groups; ※※P = 0.0063 COL1A1-EVs vs Saline at day 7; ※※※P < 0.001 COL1A1-EVs vs Saline at days 14, 21 and 28; **P = 0.0082 COL1A1-LNPs vs Saline at day 21; ***P < 0.001 COL1A1-LNPs vs Saline at day 28; ††P = 0.0040 RA vs Saline at day 28; ##P = 0.0018 Unloaded EVs vs Saline at day 28). All data are from three independent experiments and are presented as means ± s.e.m. Two-way ANOVA was used for the comparisons in c and d. The schematic in a was created with BioRender.com.
To assess the immunogenic side effects of COL1A1-LNPs and COL1A1-EVs, treated skin was excised from a subgroup of animals and analysed for inflammation 24 h after injection. Skin treated with COL1A1-LNPs exhibited redness and swelling as compared with sham controls and skin treated with COL1A1-EVs (Extended Data Fig. 3a). Flow cytometry and enzyme-linked immunoassay (ELISA) of tissue further revealed massive leucocytic infiltrate dominated by neutrophils in tissue that received COL1A1-LNPs (Extended Data Fig. 3b,c) and high levels of inflammatory cytokines such as TNF-α, IL-6, IL-1β and IFN-γ (Extended Data Fig. 3d–f). By comparison, tissue that received COL1A1-EV treatment did not exhibit a strong inflammatory reaction. These findings suggest that LNPs are substantially more immunogenic than EVs.
After assessment at day 28, a subset of animals was retained for 4 additional weeks to monitor wrinkle reduction duration. Dermal wrinkles were seen to reappear as early as 1 week later, beginning at day 35 after treatment initiation, and by day 56, dermal wrinkles were statistically indistinguishable from pre-treatment levels (Extended Data Fig. 4).
To evaluate dermal collagen engraftment from COL1A1-EV delivery, skin was excised from all groups at day 28 after treatment initiation and evaluated by immunofluorescence microscopy, as well as Masson trichrome staining. Histological analysis and fluorescence intensity quantification revealed that COL1A1 protein expression was significantly restored after COL1A1-EV treatment relative to other groups (Fig. 4a,b). Masson trichrome staining confirmed higher levels of collagen staining and dermal thickness (197.95 ± 24.46 μm COL1A1-EVs vs 95.19 ± 10.66 μm Saline at day 28, P < 0.001) in mice given COL1A1-EVs than in mice given saline control, retinoic acid, unloaded EVs or COL1A1-LNPs (Fig. 4c,d).

a, Immunofluorescence staining for GFP and COL1A1 (RFP) protein in sham control group, saline control group, RA treatment group, unloaded EV treatment group, COL1A1-LNPs and COL1A1-EV treatment groups. COL1A1-EV-treated mice exhibited GFP+ COL1A1 protein grafts in the dermis and subcutis at 28 d after initiation of treatment. Scale bar, 200 µm. b, Fluorescence intensity of COL1A1 protein (RFP) for all treatment groups (n = 3 for all groups; ※※※P < 0.001 COL1A1-EVs vs Saline; ***P < 0.001 COL1A1-LNPs vs Saline; #P = 0.0386 Unloaded EVs vs Saline; †P = 0.0218 RA vs Saline; +P = 0.0255 COL1A1-EVs vs COL1A1-LNPs). c, Representative Masson trichrome collagen staining of the epidermis, dermis and subcutis for all mouse groups. Collagen is blue. Scale bar, 200 µm. d, Quantification of dermal thickness showing increased collagen fibres in the COL1A1-EVs group (n = 3 for all groups; ※※※P < 0.001 COL1A1-EVs vs Saline; ***P < 0.001 COL1A1-LNPs vs Saline; #P = 0.0437 Unloaded EVs vs Saline; †P = 0.0447 RA vs Saline; +++P < 0.001 COL1A1-EVs vs COL1A1-LNPs). All data are from three independent experiments and are presented as means ± s.e.m. One-way ANOVA was used for the comparisons in b and d.
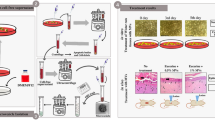
Design of a microneedle-based system for the delivery of EV mRNAs
As wrinkles were found to return to baseline levels in photoaged animals within 1 month of COLA1-EV treatment cessation, we next aimed to improve the duration of protein replacement and the effectiveness of collagen engraftment by designing an HA microneedle formulation (COL1A1-EV MN) to improve EV-mediated mRNA tissue delivery. Custom microneedle patches were prepared by using a micromoulding method in which COL1A1-EVs were mixed with a 15% HA solution dissolved in PBS and cast into the tips of a polydimethylsiloxane (PDMS) mould by maintaining a vacuum for 30 min, after which 1 ml of 15 wt% HA solution was placed into the micromould and then transferred to 4 °C for 4 h for solidification (Fig. 5a). Each needle of the COL1A1-EV microneedle patch was moulded in a conical shape, with a circular diameter of 400 µm at their base and a height of 1,000 µm (Fig. 5b). The mechanical strength of patches with 10%, 15% or 20% HA loaded with EVs was evaluated with a tensile testing machine (Supplementary Fig. 7a). The load fracture force of our 15% COL1A1-EV MN was confirmed to be higher than the minimum average force needed for skin penetration (0.058 N)37 (Supplementary Fig. 7b). Haematoxylin and eosin (H&E) staining confirmed that the microneedles penetrated through the stratum corneum into the dermis (516 ± 76 μm)38 (Fig. 5c). COL1A1-EVs loaded into HA microneedles were found to have stable appearance and membrane integrity as evaluated by transmission electron microscopy (TEM) and AFM, as well as stable COL1A1 mRNA and particle size distribution (Supplementary Fig. 7c–f). For delivery into tissue, COL1A1-EV MN patches were pressed into the dorsal skin of mice, and the microneedle base was removed after 15 min (Supplementary Video 1). During this period, the microneedles dissolved completely, with no visible skin irritation or marking at the site of administration (Fig. 5d and Supplementary Video 2). To compare the tissue distribution of EVs delivered via microneedle with that of EVs delivered by insulin syringe, EVs were labelled with DiI (1,1′-dioctadecyl-3,3,3′,3′-tetramethylindocarbocyanine perchlorate; Beyotime, C1036)39 and administered intradermally under the dorsal skin of recipient mice, with subsequent histological assessment by immunofluorescence microscopy (Fig. 5e and Supplementary Fig. 8a,b). Tissue distribution analysis revealed that syringe needle injection resulted in uneven delivery of EVs with clumping in specific areas of the dermis and subcutis, whereas EVs delivered by microneedle were better dispersed in the dermis and subcutis (Fig. 5f and Supplementary Fig. 8c,d). Assessment of EV membrane integrity by cryo-EM showed less EV membrane lysis in the EVs delivered with the microneedle patch vs the syringe needle (18.1 ± 3.0% COL1A1-EV MN vs 28.3 ± 2.4% needle injection, P = 0.022) (Supplementary Fig. 8e,f). To assess whether the improved tissue distribution and EV membrane protection resulted in improved EV engraftment, we next employed serial in vivo fluorescence imaging of DiI-labelled EVs injected intradermally in mice over a 14 d period. Fluorescence imaging confirmed similar DiI EV signal between syringe-delivered EVs and microneedle-delivered EVs for the first 4 d after injection. However, the COL1A1-EV microneedle group had significantly higher fluorescence signal at day 4 (69.61 ± 6.4% COL1A1-EV MN vs 45.79 ± 2.5% needle injection, P < 0.001) and at day 7 (35.78 ± 5.9% COL1A1-EV MN vs 19.38 ± 3.8% needle injection, P < 0.001), lasting up to day 10 post injection (30.22 ± 2.6% COL1A1-EV MN vs 6.68 ± 1.3% needle injection, P < 0.001) (Fig. 5g,h). Ex vivo organ distribution analyses of DiI probe fluorescence over a 2 week time period revealed similar tissue distribution and signal decay across animals receiving intradermal injection and animals receiving microneedle delivery, suggesting that the reduction in signal intensity is due to metabolism of the DiI probe as shown in other similar studies40,41 (Supplementary Fig. 9a,b). Taken together, these findings indicate that use of the microneedle patch improves long-term retainment of EVs in tissue.

a, Schematic illustration of microneedle fabrication. b, Microscope and scanning electron microscopy images of microneedle arrays. Scale bar, 500 µm. c, H&E-stained section of mouse skin shows penetration of single microneedle. Scale bar, 100 µm. d, Top: time course of HA EV microneedle tips pressed into skin; the microneedles dissolved within 15 min of application. Scale bars, 200 µm. Bottom: skin recovery after HA EV microneedle treatment shows minimal irritation. Scale bars, 5 mm. e, Skin histology of Dil-labelled EVs shows highly concentrated EVs (red) unevenly distributed in the subcutis after syringe needle injection, whereas microneedle-delivered EVs were more evenly distributed in tissue. Yellow dashed lines encircle representative subcutaneous hair bulbs and border representative rod-like portions of the follicle extending upward to the dermis. Scale bar, 100 µm. f, Representative EV distribution analysed by ImageJ software. g, In vivo fluorescence images of nude mice treated with intradermal injection or HA microneedle patch delivery of Dil-labelled EVs on days 1, 2, 4, 7, 10 and 14 after delivery (n = 3 for all groups). h, Quantification of fluorescence intensity over the 14 d treatment period. Data are normalized to the fluorescence intensity at day 1. All data are from three independent experiments and are presented as means ± s.e.m. ***P < 0.001 COL1A1-EV MN delivery group vs needle delivery group. Two-way ANOVA was used for the comparisons in h. The schematic in a was created with BioRender.com.
Therapeutic efficacy of COL1A1-EV delivery via microneedles
We next tested whether use of our custom COL1A1-EV microneedle patch for delivering COL1A1-EVs could result in improved in vivo protein replacement in the skin of photoaged mice (Supplementary Fig. 10). Athymic mice were again subjected to 8 weeks UV irradiation and assigned to 1 of 5 treatment groups: (1) saline control, (2) COL1A1 mRNA EVs delivered by syringe needle, (3) HA microneedle control, (4) unloaded EVs delivered by HA microneedle (Unloaded EV microneedle) and (5) COL1A1 mRNA-loaded EVs delivered by HA microneedle (COL1A1-EV MN). All mice were given a single-dose injection of 22 × 109 copy number of COL1A1 mRNA (or an equivalent vehicle volume for control groups) at day 0 of the treatment timeline. After injection, all mice were monitored via microscopic photography of dorsal skin wrinkles for up to 3 months (Fig. 6a). Relative to syringe needle injection, which reduced wrinkle formation for up to 35 d before a return to pre-treatment baseline, delivery of COL1A1 mRNA by COL1A1-EV MN was found to substantially reduce wrinkle area and number for up to 70 d before a return to baseline levels (Fig. 6b,c). To further confirm these findings, a subset of mice from each group were killed at 1 month, 2 months and 3 months after treatment for skin replica plaster assessment of the dorsal skin and histology (Extended Data Fig. 5a–c). Both needle injection and COL1A1-EV MN treatments reduced wrinkle length and depth for up to 1 month post treatment (Extended Data Fig. 5d,e). However, only mice treated with COL1A1-EV MN showed significantly long-lasting reductions in wrinkle length and depth for up to the 2 month timepoint. To test whether collagen replacement and wrinkle treatment could be maintained for a longer period of time, we subjected additional animal cohorts to serial treatment of COL1A1-EVs delivered by syringe needle and microneedle every 30 d. Both COL1A1-EVs delivered by syringe needle and COL1A1-EV MN were found to reduce wrinkle number and area for as long as the animals received treatment, with COL1A1-EV MN animals demonstrating the most improvement (Extended Data Fig. 6).

a, Long-term (91 d) observation of 4 treatment groups after a single injection: (1) saline control, (2) COL1A1-EVs (22 × 109 copy number COL1A1 mRNA) delivered by 28G syringe needle, (3) HA microneedle control, (4) unloaded EVs delivered by HA microneedle (Unloaded EV microneedle) and (5) COL1A1-EV MN (22 × 109 copy number COL1A1 mRNA), (n = 4 for all groups). Scale bar, 5 mm b, Quantification of total numbers of wrinkles (n = 4 for all groups; *P = 0.014 COL1A1-EV MN vs Saline at day 7; *P = 0.038 COL1A1-EV MN vs Saline at day 14; *P = 0.012 COL1A1-EV MN vs Saline at day 21; *P = 0.039 COL1A1-EV MN vs Saline at day 28; *P = 0.031 COL1A1-EV MN vs Saline at day 35; *P = 0.03 COL1A1-EV MN vs Saline at day 42; *P = 0.022 COL1A1-EV MN vs Saline at day 49; *P = 0.031 COL1A1-EV MN vs Saline at day 56; **P = 0.008 COL1A1-EV MN vs Saline at day 63; #P = 0.030 Needle injection vs Saline at day 7; #P = 0.041 Needle injection vs Saline at day 14; †P = 0.047 HA microneedle vs Saline at day 14). c, Quantification of total dorsal skin wrinkle area (n = 4 for all groups; *P = 0.016 COL1A1-EV MN vs Saline at day 7; *P = 0.038 COL1A1-EV MN vs Saline at day 14; *P = 0.042 COL1A1-EV MN vs Saline at days 21 and 28; *P = 0.026 COL1A1-EV MN vs Saline at day 42; *P = 0.048 COL1A1-EV MN vs Saline at day 49; #P = 0.033 Needle injection vs Saline at day 7; #P = 0.031 Needle injection vs Saline at day 14; #P = 0.042 Needle injection vs Saline at day 21). All data are from three independent experiments and are presented as means ± s.e.m. Two-way ANOVA was used for the comparisons in b and c.
Histologic analysis of skin samples taken after treatment confirmed the lasting engraftment of GFP+ COL1A1 protein engraftment in the dermis and subcutis of mice in both the needle injection group and the COL1A1-EV microneedle group at 1 month after delivery (Fig. 7a). However, by the 2 month timepoint, only the mice in the COL1A1-EV microneedle cohort demonstrated GFP+ COL1A1 engraftment in the dermis and subcutis (Fig. 7b). At 3 months after delivery, skin samples in all groups failed to exhibit any GFP collagen engraftment (Fig. 7c). COL1A1+ GFP immunofluorescence intensity was quantified, supporting wrinkle microscopy findings which demonstrated effective treatment at days 30–60 and a return to pre-treatment baseline from days 70–90 (Fig. 7d). Immunohistochemical staining of COL1A1 protein and Masson trichrome staining of skin tissues confirmed that the amount of collagen in the dermis correlated with immunofluorescence microscopy findings and that among all cohorts, the COL1A1-EV MN group had the most abundant collagen fibres and highest dermal thickness (180.14 ± 21.46 μm COL1A1-EV MN vs 96.61 ± 14.00 μm Saline at 1 month, P < 0.001; 154.88 ± 8.27 μm COL1A1-EV MN vs 109.25 ± 10.86 μm Saline at 2 months, P < 0.05) (Fig. 7e,f). Taken together, these findings indicate that encapsulation and delivery of COL1A1 mRNA via our COL1A1-EV MN system can prolong collagen protein replacement in photoaged skin for more than twice the duration of syringe needle delivery.

a–c Immunofluorescence staining of GFP and COL1A1 (RFP) demonstrates GFP-positive COL1A1 protein grafts in the skin of mice receiving COL1A1-EVs via 28G needle injection and via COL1A1-EV MN for up to 1 month (28 d) after delivery. At 2 months (63 d), only mice that were treated with COL1A1-EV MN had long-term GFP-positive COL1A1 engraftment. No evidence of GFP-positive collagen protein could be detected in any mice by 3 months (91 d) after delivery. Scale bar, 200 µm. d, Quantification of GFP and COL1A1 (RFP) co-localized fluorescence signal demonstrates long-term COL1A1-EV-derived collagen engraftment in the skin of mice given COL1A1-EV MN vs mice given COL1A1-EVs via 28G needle injection and control groups (n = 3 for all groups; ***P < 0.001 COL1A1-EV MN vs Needle injection at 2 months). e, Immunohistochemical staining of COL1A1 protein and Masson trichrome staining at 1 month (28 d), 2 months (63 d) and 3 months (91 d). Scale bar, 200 µm. f, Quantification of dermal thickness by Masson trichrome staining (n = 3 for all groups; **P = 0.0062 COL1A1-EV MN vs Saline at 1 month; *P = 0.034 COL1A1-EV MN vs Saline at 2 months). All data are from three independent experiments and are presented as means ± s.e.m. Two-way ANOVA was used for the comparisons in d and f.
Discussion
Extracellular vesicles have emerged as a next-generation drug-delivery system because of their inherent properties of biocompatibility, low immunogenicity and ability to be derived from healthy human cells12. Nonetheless, most studies focusing on nucleic-acid delivery so far have encapsulated small molecules in the 10–20 nt range such as microRNAs and small interfering RNA (siRNAs) as the payload, whereas larger nucleic acids such as mRNAs are seldom evaluated owing to the difficulty of loading them into EVs42. With the recent advancement in utility of mRNA-based therapeutics for treating human diseases, there has been increased interest in using EVs as an mRNA-delivery system. Recently, we developed an mRNA-loading technique that allows for the large-scale production of EVs containing intact endogenous mRNA for nucleic-acid therapy14. Here we have shown that mRNA-loaded EVs can be applied for protein-replacement therapy in a model of dermal collagen photodepletion17. We have shown that CNP is able to load high copy numbers of COL1A1 mRNA (~4,000+ nt) into EVs, which cannot be achieved by post-insertion loading methods43,44. In vivo results showed that these COL1A1-EVs can restore COL1A1 protein expression in the mouse skin after photoaging. We also examined the presence of COL1A1 mRNA and protein over time in vivo, and found that the corresponding protein was translated as early as 12 h after delivery with a peak at 4 d, and that this was sustained for several weeks, depending on dose. Several studies have generated ex vivo COL1A1 protein as a collagen filler; here we characterized the in vivo kinetics of exogenous COL1A1 mRNA delivery and protein expression in vivo45.
To adapt our approach for long-term protein replacement, we further developed a microneedle array for the delivery of COL1A1-EVs to allow for an even distribution of EVs into local tissue, with reduced membrane rupture. HA is a critical element of the extracellular matrix and has been verified to have excellent biocompatibility with skin tissues and various biomaterial systems46,47. By integrating EVs into an HA microneedle biomaterial, we were able to extend protein engraftment of COL1A1 to more than 60 d in the skin specimens evaluated. Notably, EVs not loaded with COL1A1 mRNA were also able to achieve a modest decrease in wrinkle number, which is consistent with studies finding that ‘empty’ EVs from certain parental cells are potential therapeutic candidates owing to endogenous cargos (still, the detailed mechanisms underlying such phenomena need further exploration)36.
Compared with the relatively long-lasting expression of DNA-based gene therapies, mRNA therapeutics may help advance gene therapy and lower the risks of adverse events owing to the retention of the mRNAs in the cytoplasm, without penetration into the nucleus48,49. Hence, mRNA-based modalities have advantages over DNA-based and virus-based gene therapies, such as the ability to bypass the traditional transcription process, and no risk of genomic integration (as opposed to the use of some adeno-associated virus vectors, which traditionally has been considered to have a low risk profile50). However, the clinical translation of mRNA therapeutics is still limited. For mRNA products developed using LNPs, the immunogenicity of PEGylation components on the LNP surface as well as some carrier formulations have been linked to inflammation and to numerous adverse safety events51,52. Indeed, when we compared the delivery of COL1A1-LNPs and COL1A1-EVs in vivo, we found that COL1A1-LNPs were able to yield collagen protein and reduce dermal wrinkles, but also caused notable inflammatory infiltrate in local tissue, whereas COL1A1-EVs did not. Several recent studies have shown that EVs are characterized by low levels of immunogenicity, with some preclinical reports and clinical trials (clinicaltrials.gov: NCT05191381, NCT05216562) suggesting that EVs derived from a variety of cell types have immunosuppressive effects, although it should be noted that these studies primarily used mesenchymal stromal cells and dendritic cells as donor cell types36,53,54.
Future challenges for the clinical application of COL1A1-EV MN include the optimization of microneedle geometry with more densely protected EVs as well as optimized storage conditions because mRNA is subject to rapid degradation when not retained at −80 °C or below55. Ideally, future EV systems can be packaged as ready-to-use aliquots and shipped at appropriate temperatures, which would greatly improve the practicability of COL1A1-EV MN systems for clinical use. We would also aim to expand therapeutic applications in our system to include items such as COL7 for genetic orphan diseases such as epidermolysis bullosa56. We have only delivered mRNA via the microneedle system, yet the system is also suitable for packaging other types of EV cargos, such as miRNAs and siRNAs, and other bioactive therapeutic agents such as peptides and proteins57,58,59. Although many challenges would need to be overcome before the microneedle-based EV-delivery system can be tried in humans, because of the improved biocompatibility and benign side-effect profile of EVs as compared with LNPs and adeno-associated virus (AAV), we believe that the system might constitute a universal nucleic-acid carrier for the treatment of a range of human diseases and conditions.
Methods
Cell culture
nHDFs (PCS-201-010) were purchased from ATCC and cultured in DMEM medium (Thermo Fisher) containing 10% heat-inactivated fetal bovine serum (FBS; 10099141C, Thermo Fisher) at 37 °C in humidified conditions equilibrated with 5% CO2.
Cellular nanoporation and electroporation
For CNP, a single layer of nHDF was seeded on a 1 cm × 1 cm 3D CNP surface for overnight incubation as previously described. Human COL1A1 cDNA (NM_000088.3) plasmid with GFP tag was purchased from Sino Biological (HG11776-ACG). Plasmids pre-loaded in PBS buffer were injected into individual cells via nanochannels using an electric field strength of 250 V cm−1 with 10 pulses at 10 ms per pulse, with a 0.1 s interval. Various electroporation conditions were tested to determine optimal conditions. BEP (gene pulser xcell, Bio-rad) was conducted using an electric field strength of 1,250 V cm−1 with 1 pulse of 20 ms. pCMV-COL1A1-GFP plasmids were prepared at a concentration of 500 ng ml−1 in PBS for transfection.
Collection and purification of EVs secreted by donor cells
Cells were cultured in DMEM medium containing serum. The cell culture medium containing serum was removed when conducting CNP. Cells were then washed with PBS 3 times and cultured in serum-free cell culture medium for 24 h after CNP. EVs were collected from cell culture supernatants. In brief, the cell culture medium (CCM) was centrifuged at 200 × g for 5 min to remove cells and debris, after which it was centrifuged again at 2,000 × g for 30 min. Amicon Ultra-4 centrifugal filter unit (10 kDa, Millipore, 801024) was used to concentrate the CCM. The EV sample was purified using total exosome isolation reagent (Invitrogen, 4478360). EV particle size and number were measured using NanoSight NS300 (Malvern). The RNA yield and size distribution were analysed using an Agilent 2100 Bioanalyzer with an RNA 6000 Pico kit (Agilent Technologies).
RT–qPCR of EV-containing RNA expression levels
The expression of human COL1A1 mRNA in EVs was measured using RT–qPCR following the manufacturer’s recommended protocol. In brief, total RNA from purified EVs was obtained using an RNA purification mini kit (Norgen Biotek, 55000) and a DNA removal kit (Norgen Biotek, 25720). A SuperScript III First-Strand Synthesis system (Invitrogen) was used to synthesize the first-strand complementary DNA, with random hexamers as primers. The expression of genes was measured using TB Green Premix Ex Taq II (Takara, RR820). The primer sequences used were as follows: COL1A1 (human), forward: 5′-CCTGGAAAGAATGGAGATGA-3′ and reverse: 5′-ACCATCCAAACCACTGAAAC-3′; Gapdh (human), forward: 5′-CAGCCTCAAGATCATCAGCA - 3′ and reverse: 5′ - AGAGGCAGGGATGATGTTCT - 3′ (Supplementary Table 1).
In situ hybridization for human COL1A1 mRNA in skin tissue
Nude female mice (10–12 weeks old) were anaesthetized with isoflurane, and the dorsal skin region was injected with 50 μl 2.7 × 109 copy number of CNP COL1A1 mRNA EVs. At predefined timepoints (12, 24, 48, 96 h, 7 d, 10 d, 14 d) following the skin injection, the skin was excised and put in 4% formaldehyde for fixation. Subsequently, the slices were embedded in paraffin further sectioned into 4 μm slices. RNAscope automated in situ hybridization assay for detection of human COL1A1 mRNA was performed using the HybEZ II Hybridization System (Advanced Cell Diagnostics (ACD)), and all in situ hybridization reagents were ACD products. In brief, target retrieval was performed at 95 °C for 15 min using Leica Epitope Retrieval Buffer 2, followed by protease treatment at 42 °C for 15 min. The probe (RNAscope Probe- Hs-COL1A1, 401891, ACD) was hybridized for 2 h at 40 °C followed by RNA scope amplification, and RNAscope 2.5 HD Assay BROWN kit was used for visualization of staining. RNAscope 2.5 LS probe-Rn-Ppib was used as negative control.
Immunofluorescence microscopy
Tissue sections were fixed with 4% paraformaldehyde for 20 min at room temperature and washed 3 times with PBS (Vetec) for 5 min each. Then the tissue was transferred to 0.2% Triton X-100 for 15 min (permeabilized), followed by blocking with BSA for 40 min and the addition of primary antibody (ab34710 and ab6556, Abcam) for overnight blocking at 4 °C. Finally, the secondary antibody (ab6939 and ab6717, Abcam) was added and the tissue sections placed at room temperature for 60 min. Tissue sections were then washed with PBS, added with DAPI (ThermoFisher) for nuclear staining and then mounted for observation.
Immunohistochemical staining and analysis
Tissue sections were fixed in 4% paraformaldehyde for 20 min, washed 3 times with PBS (pH 7.4) for 5 min each and then transferred to a retrieval box containing ethylenediaminetetraacetic acid (EDTA) (pH 9.0) antigen retrieval solution for antigen retrieval in a microwave oven. Subsequently, the sections were incubated in 3% hydrogen peroxide solution at room temperature for 25 min in the dark. After washing with PBS, the tissue was evenly covered with 3% BSA or 10% normal rabbit serum, blocked at room temperature for 30 min, added with primary antibody (ab34710 and ab6556, Abcam) and incubated overnight at 4 °C. Then, a secondary antibody (HRP-labelled) corresponding to the primary antibody was added to cover the tissue sections, which were then incubated at room temperature for 50 min. For 3,3'-diaminobenzidine (DAB) colour development, the slides were placed in PBS (pH 7.4) and washed 3 times with shaking on a de-staining shaker for 5 min each time. After the slices were slightly dried, the freshly prepared DAB colour developing solution was added dropwise in the circle. Developing time was controlled under the microscope. Haematoxylin, haematoxylin differentiation solution and hematoxylin blue-returning solution were sequentially added for nuclei counterstaining. Finally, the glass slides were placed in anhydrous ethanol and xylene for dehydration and sealing.
Mice
Animal work was approved by the Institutional Animal Care and Use Committee (IACUC) of Shenzhen Bay Laboratory (No. D2021-107) or partner laboratories. To create the skin-photoaging model, female nude mice (10–12 weeks old) were subjected to UVB irradiation of the dorsal skin every other day for 8 weeks as follows: mice were anaesthetized with 1.5% isoflurane, and then UV irradiation was delivered with a UV lamp (Philips; emission spectrum 311 nm) positioned 30 cm above the dorsal skin of the mice every other day for 8 weeks. The UV irradiation intensity, represented as the minimal erythemal dose (MED), was set at 1 MED during the first 2 weeks (60 mJ cm−2), elevated to 2 MED (120 mJ cm−2) in the third week, 3 MED (180 mJ cm−2) in the fourth week and 4 MED (240 mJ cm−2) during the fifth to eighth weeks of the experiment. The total irradiated UVB volume was approximately 80 MED. For the syringe-based treatment of photoaged skin, nude mice, after the 8 week irradiation period described above, were assigned to 1 of 6 treatment groups (4 mice each): (1) UVB irradiation + saline, (2) UVB irradiation + 0.05% retinoic acid, (3) UVB irradiation + unloaded nHDF-EVs delivered with a 32G Hamilton syringe, (4) UVB irradiation + 2.7 × 109 copy number COL1A1 mRNA COL1A1-LNPs, (5) UVB irradiation + 2.7 × 109 copy number COL1A1 mRNA COL1A1- EVs delivered with a 32G Hamilton syringe and (6) no UVB exposure (sham). Skin treatments took place on days 0, 4, 7, 14 and 21. The whole back skin was divided into three parts for analysis.
Skin replica
A SILFLO silicone replica and ring locator were purchased from Clinical & Derm. Replicas of the back (dorsal) skin of the mice were obtained at the end of the treatment period. The replicas were analysed by stereomicroscopy (Olympus SZX7), and corresponding images were analysed by ImageJ (NIH).
Preparation of COL1A1-EV MN patches and local tissue delivery
The fabrication of the microneedle patch was performed using a silicone micromould, with each needle cavity being 400 μm in a round base diameter and 1,000 μm in height. These needle cavities were arranged in a 10 × 10 array with 700 μm tip to tip spacing. For the preparation of the microneedle patch, 150 μl of 15 wt% HA solution was mixed with 50 μl EVs, kept under vacuum for 30 min and then transferred to 4 °C until it was deposited in the needle cavity. Finally, 1 ml of 15 wt% HA solution was loaded onto the micromould and the micromould placed in a drying chest with an attached fan to accelerate the evaporation process for solidification. After solidification, the microneedle patch was detached from the silicone mould for further use. To assess COL1A1-EV delivery via the customized microneedle patches, nude mice were irradiated for 8 weeks as described above and assigned to 1 of 5 treatment groups (4 mice each): (1) UVB irradiation + saline, (2) UVB irradiation + 22 × 109 copy number COL1A1 mRNA COL1A1-EVs delivered by 28G Hamilton syringe, (3) UVB irradiation + HA microneedle patch, (4) UVB irradiation + 1010 unloaded nHDF-EVs delivered via microneedle (unloaded EV microneedle) and (5) UVB irradiation + 15% HA microneedle mixed with 22 × 109 copy number COL1A1 mRNA COL1A1- EVs.
Fluorescence in vivo imaging
Microneedle-treated mice were imaged with an in vivo imaging system (IVIS Spectrum, Perkin Elmer) on days 1, 2, 4, 7, 10 and 14. The parameters were set as follows: exposure time 15 s, excitation 570 nm, emission 680 nm, 2F/stop and 13.6 cm field of view in the specified fluorescence imaging times. Quantitative analysis of RFP fluorescence intensity was performed by measuring the average radiation efficiency (photon s−1 cm−2 sr−1 µW−1) in a region of interest. Data were normalized to fluorescence intensity on day 1.
Statistical analysis
Quantitative data are represented as mean ± s.e.m. No data were excluded in this study. To analyse the statistical difference between two groups, two-sided Student’s t-tests were used for the comparisons. For two or more groups, one-way analysis of variance (ANOVA) with Tukey’s multiple-comparison test was used to analyse the statistical difference. Two-way ANOVA was chosen for analysing the statistical difference between data points in groups. P < 0.05 was considered statistically significant. GraphPad Prism 8.3 was used for data analysis.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The main data supporting the results in this study are available within the paper and its Supplementary Information. Source data for the figures are available from figshare at https://figshare.com/articles/dataset/SD_FIGS_xlsx/21514641. The raw and analysed datasets generated during the study are available for research purposes from the corresponding authors on reasonable request.
References
Sahin, U., Kariko, K. & Tureci, O. mRNA-based therapeutics–developing a new class of drugs. Nat. Rev. Drug Discov. 13, 759–780 (2014).
Muik, A. et al. Neutralization of SARS-CoV-2 Omicron by BNT162b2 mRNA vaccine-elicited human sera. Science 375, 678–680 (2022).
Kowalski, P. S., Rudra, A., Miao, L. & Anderson, D. G. Delivering the messenger: advances in technologies for therapeutic mRNA delivery. Mol. Ther. 27, 710–728 (2019).
Wang, C., Zhang, Y. & Dong, Y. Lipid nanoparticle-mRNA formulations for therapeutic applications. Acc. Chem. Res. 54, 4283–4293 (2021).
Qiu, M., Li, Y., Bloomer, H. & Xu, Q. Developing biodegradable lipid nanoparticles for intracellular mRNA delivery and genome editing. Acc. Chem. Res. 54, 4001–4011 (2021).
Lokugamage, M. P. et al. Mild innate immune activation overrides efficient nanoparticle-mediated RNA delivery. Adv. Mater. 32, e1904905 (2020).
Szebeni, J. et al. Applying lessons learned from nanomedicines to understand rare hypersensitivity reactions to mRNA-based SARS-CoV-2 vaccines. Nat. Nanotechnol. 17, 337–346 (2022).
Moghimi, S. M. Allergic reactions and anaphylaxis to LNP-based COVID-19 vaccines. Mol. Ther. 29, 898–900 (2021).
Valadi, H. et al. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 9, 654–659 (2007).
Cheng, L. & Hill, A. F. Therapeutically harnessing extracellular vesicles. Nat. Rev. Drug Discov. 21, 379–399 (2022).
de Jong, O. G. et al. Drug delivery with extracellular vesicles: from imagination to innovation. Acc. Chem. Res. 52, 1761–1770 (2019).
Herrmann, I. K., Wood, M. J. A. & Fuhrmann, G. Extracellular vesicles as a next-generation drug delivery platform. Nat. Nanotechnol. 16, 748–759 (2021).
van Niel, G., D’Angelo, G. & Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 19, 213–228 (2018).
Yang, Z. et al. Large-scale generation of functional mRNA-encapsulating exosomes via cellular nanoporation. Nat. Biomed. Eng. 4, 69–83 (2020).
Sharma, M. R., Werth, B. & Werth, V. P. Animal models of acute photodamage: comparisons of anatomic, cellular and molecular responses in C57BL/6J, SKH1 and Balb/c mice. Photochem. Photobiol. 87, 690–698 (2011).
Varani, J. et al. Decreased collagen production in chronologically aged skin: roles of age-dependent alteration in fibroblast function and defective mechanical stimulation. Am. J. Pathol. 168, 1861–1868 (2006).
Fisher, G. J. W. Z. Pathophysiology of premature skin aging induced by ultraviolet light. N. Engl. J. Med. 337, 1419–1428 (1997).
Buranasudja, V., Rani, D., Malla, A., Kobtrakul, K. & Vimolmangkang, S. Insights into antioxidant activities and anti-skin-aging potential of callus extract from Centella asiatica (L.). Sci. Rep. 11, 13459 (2021).
Wada, N., Sakamoto, T. & Matsugo, S. Mycosporine-like amino acids and their derivatives as natural antioxidants. Antioxidants 4, 603–646 (2015).
Xiong, Z. M. et al. Ultraviolet radiation protection potentials of methylene blue for human skin and coral reef health. Sci. Rep. 11, 10871 (2021).
Bielli, A. et al. Cellular retinoic acid binding protein-II expression and its potential role in skin aging. Aging 11, 1619–1632 (2019).
Song, H., Zhang, S., Zhang, L. & Li, B. Effect of orally administered collagen peptides from bovine bone on skin aging in chronologically aged mice. Nutrients 9, 1209 (2017).
Jeong, S. et al. Anti-wrinkle benefits of peptides complex stimulating skin basement membrane proteins expression. Int. J. Mol. Sci. 21, 73 (2019).
Lee, A. Y. Skin pigmentation abnormalities and their possible relationship with skin aging. Int. J. Mol. Sci. 22, 3727 (2021).
Kim, J. H. et al. Comparative evaluation of the effectiveness of novel hyaluronic acid-polynucleotide complex dermal filler. Sci. Rep. 10, 5127 (2020).
Urdiales-Galvez, F., Martin-Sanchez, S., Maiz-Jimenez, M., Castellano-Miralla, A. & Lionetti-Leone, L. Concomitant use of hyaluronic acid and laser in facial rejuvenation. Aesthetic Plast. Surg. 43, 1061–1070 (2019).
Fisher, G. J., Varani, J. & Voorhees, J. J. Looking older: fibroblast collapse and therapeutic implications. Arch. Dermatol. 144, 666–672 (2008).
Quan, T. et al. Enhancing structural support of the dermal microenvironment activates fibroblasts, endothelial cells, and keratinocytes in aged human skin in vivo. J. Invest. Dermatol. 133, 658–667 (2013).
Shakouri, R. et al. In vivo study of the effects of a portable cold plasma device and vitamin C for skin rejuvenation. Sci. Rep. 11, 21915 (2021).
de Araujo, R., Lobo, M., Trindade, K., Silva, D. F. & Pereira, N. Fibroblast growth factors: a controlling mechanism of skin aging. Skin Pharmacol. Physiol. 32, 275–282 (2019).
Cole, M. A., Quan, T., Voorhees, J. J. & Fisher, G. J. Extracellular matrix regulation of fibroblast function: redefining our perspective on skin aging. J. Cell Commun. Signal. 12, 35–43 (2018).
Shi, J. et al. A review on electroporation-based intracellular delivery. Molecules 23, 3044 (2018).
Todorova, K. & Mandinova, A. Novel approaches for managing aged skin and nonmelanoma skin cancer. Adv. Drug Deliv. Rev. 153, 18–27 (2020).
Choi, J. S. et al. Functional recovery in photo-damaged human dermal fibroblasts by human adipose-derived stem cell extracellular vesicles. J. Extracell. Vesicles 8, 1565885 (2019).
Alpermann, H. & Vogel, H. G. Effect of repeated ultraviolet irradiation on skin of hairless mice. Arch. Dermatol. Res. 262, 15–25 (1978).
Hu, S. et al. Needle-free injection of exosomes derived from human dermal fibroblast spheroids ameliorates skin photoaging. ACS Nano 13, 11273–11282 (2019).
Kim, J. D., Kim, M., Yang, H., Lee, K. & Jung, H. Droplet-born air blowing: novel dissolving microneedle fabrication. J. Control. Release 170, 430–436 (2013).
Abd, E. et al. Skin models for the testing of transdermal drugs. Clin. Pharmacol. 8, 163–176 (2016).
Zheng, T. et al. Plasma exosomes spread and cluster around β-amyloid plaques in an animal model of Alzheimer’s disease. Front. Aging Neurosci. 9, 12 (2017).
Li, Z. et al. Exosome-based Ldlr gene therapy for familial hypercholesterolemia in a mouse model. Theranostics 11, 2953–2965 (2021).
Cao, H. et al. In vivo real-time imaging of extracellular vesicles in liver regeneration via aggregation-induced emission luminogens. ACS Nano 13, 3522–3533 (2019).
Kamerkar, S. et al. Exosomes facilitate therapeutic targeting of oncogenic KRAS in pancreatic cancer. Nature 546, 498–503 (2017).
O’Brien, K., Breyne, K., Ughetto, S., Laurent, L. C. & Breakefield, X. O. RNA delivery by extracellular vesicles in mammalian cells and its applications. Nat. Rev. Mol. Cell Biol. 21, 585–606 (2020).
Yin, H. et al. Non-viral vectors for gene-based therapy. Nat. Rev. Genet. 15, 541–555 (2014).
Zhang, Z. et al. COL1A1: a potential therapeutic target for colorectal cancer expressing wild-type or mutant KRAS. Int. J. Oncol. 53, 1869–1880 (2018).
Yu, J. et al. Microneedle-array patches loaded with hypoxia-sensitive vesicles provide fast glucose-responsive insulin delivery. Proc. Natl Acad. Sci. USA 112, 8260–8265 (2015).
Wu, T. et al. Microneedle-mediated biomimetic cyclodextrin metal organic frameworks for active targeting and treatment of hypertrophic scars. ACS Nano 15, 20087–20104 (2021).
Paunovska, K., Loughrey, D. & Dahlman, J. E. Drug delivery systems for RNA therapeutics. Nat. Rev. Genet. 23, 265–280 (2022).
van Haasteren, J., Li, J., Scheideler, O. J., Murthy, N. & Schaffer, D. V. The delivery challenge: fulfilling the promise of therapeutic genome editing. Nat. Biotechnol. 38, 845–855 (2020).
Mullard, A. Gene therapy community grapples with toxicity issues, as pipeline matures. Nat. Rev. Drug Discov. 20, 804–805 (2021).
Shi, D. et al. To PEGylate or not to PEGylate: immunological properties of nanomedicine’s most popular component, polyethylene glycol and its alternatives. Adv. Drug Deliv. Rev. 180, 114079 (2022).
Knop, K., Hoogenboom, R., Fischer, D. & Schubert, U. S. Poly(ethylene glycol) in drug delivery: pros and cons as well as potential alternatives. Angew. Chem. Int. Ed. Engl. 49, 6288–6308 (2010).
Qian, X. et al. Immunosuppressive effects of mesenchymal stem cells-derived exosomes. Stem Cell Rev. Rep. 17, 411–427 (2021).
Kim, S. H. et al. Exosomes derived from IL-10-treated dendritic cells can suppress inflammation and collagen-induced arthritis. J. Immunol. 174, 6440–6448 (2005).
Chaudhary, N., Weissman, D. & Whitehead, K. A. mRNA vaccines for infectious diseases: principles, delivery and clinical translation. Nat. Rev. Drug Discov. 20, 817–838 (2021).
Peking, P., Koller, U. & Murauer, E. M. Functional therapies for cutaneous wound repair in epidermolysis bullosa. Adv. Drug Deliv. Rev. 129, 330–343 (2018).
Vader, P., Mol, E. A., Pasterkamp, G. & Schiffelers, R. M. Extracellular vesicles for drug delivery. Adv. Drug Deliv. Rev. 106, 148–156 (2016).
Born, L. J., Harmon, J. W. & Jay, S. M. Therapeutic potential of extracellular vesicle-associated long noncoding RNA. Bioeng. Transl. Med. 5, e10172 (2020).
Pi, F. et al. Nanoparticle orientation to control RNA loading and ligand display on extracellular vesicles for cancer regression. Nat. Nanotechnol. 13, 82–89 (2018).
Acknowledgements
We thank C. Wogan of the Division of Radiation Oncology, MD Anderson Cancer Center, for editorial assistance.
Author information
Authors and Affiliations
Contributions
A.S.L. and F.L. conceived the work; A.S.L., W.J., F.L., Z.Y. and B.Y.S.K. supervised the research; A.S.L., J.S., L.J.L. and K.J.K. developed the technology; A.S.L., Y.Y., Y.T., F.L., W.J., Z.Y., L.T. and B.Y.S.K. designed the experiments; A.S.L., L.J.L., Z.Y., Y.T., Y.Y., W.J., F.L., B.Y.S.K., K.J.K., J.S., B.S., K.H., D.L., T.G., L.T., W.-J.L. and E.B. provided intellectual input; A.S.L., L.J.L., Z.Y., W.J., J.S., S.D., E.B. and B.Y.S.K. wrote the manuscript, with input from all authors; Y.Y., Y.T., J.S., K.J.K., Y.T., A.P.E., J.C., C.-L.C., W.-H.H. Y.L., Z.L., Y.Z., H.Z., X.L., Y.W. and J.H. conducted experiments; Y.Y., Y.T., Z.Y. and A.P.E. prepared figures and videos.
Corresponding authors
Ethics declarations
Competing interests
A.S.L. and L.J.L. are consultants and shareholders of Spot Biosystems, Ltd. J.S. and K.J.K are employees of Spot Biosystems, Ltd.
Peer review
Peer review information
Nature Biomedical Engineering thanks Sun Hwa Kim, Chuanbin Wu and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 In vitro delivery of COL1A1 mRNA-containing EVs.
a, Fluorescence images of serum-starved nHDFs treated with COL1A1-GFP EVs and protein translated from delivered COL1A1-GFP mRNA after 48 h. Scale bar, 100 µm. b, Fluorescence intensity of cells treated with COL1A1-EVs (n = 3 for all groups, ***P < 0.001 Control vs COL1A1-EVs) in 48 h. c, RT-qPCR shows higher collagen mRNA transcript levels after in vitro delivery of COL1A1 mRNA from EVs (n = 3 for all groups, ***P < 0.001 Control vs COL1A1-EVs) in 48 h. d, Western blots show elevated COL1A1 protein in treated fibroblasts (n = 3 for all groups, **P = 0.001 Control vs COL1A1-EVs).e, Pro-collagen I collected from supernatant and detected by ELISA (n = 3 for all groups, ***P < 0.001 Control vs COL1A1-EVs) in 48 h. All data are from three independent experiments and are presented as means ± SEM; two-sided Student’s t tests were used for the comparisons in (b–e).
Extended Data Fig. 2 Skin plaster assessment of dorsal skin after COL1A1-EV treatment.
a, Microscopic observations of dorsal skin and skin replicas. Scale bar, 5 mm. b, Mean wrinkle depth in skin replicas (n = 4 for all groups, ※※※P < 0.001 COL1A1-EVs vs Saline; **P = 0.0025 COL1A1-LNPs vs Saline; +++P < 0.001 COL1A1-EVs vs COL1A1-LNPs). c, Mean wrinkle length analysed on skin replicas (n = 4 for all groups, #P = 0.0203 Unloaded EVs vs Saline; *P = 0.0405 COL1A1-LNPs vs Saline; ※※※P < 0.001 COL1A1-EVs vs Saline; ++P = 0.0015 COL1A1-EVs vs COL1A1-LNPs).All data are from three independent experiments and are presented as means ± SEM. One-way analysis of variance (ANOVA) was used for the comparisons in (b, c). NS, not significant.
Extended Data Fig. 3 Assessment of in vivo immunogenicity of COL1A1-LNPs and COL1A1-EVs.
a, Skin samples from the mice injected with a single dose injection of 22E9 copy number COL1A1 mRNA in COL1A1-EVs and COL1A1-LNPs were harvested after 24 h. Skin samples of mice were analysed by flow cytometry, for b, leukocyte cell percentage, and c, neutrophil percentage (n = 3 for all groups, **P = 0.0034 COL1A1-LNPs vs sham for %CD45 + cells; ***P < 0.001 COL1A1-LNPs vs Sham for %neutrophils among CD45 + cells; NS, not significant). d, Protein quantification via ELISA for IFN-γ, IL-1β, IL-6 and TNF-α shows elevation of inflammatory cytokines in the COL1A1-LNPs group as compared to COL1A1-EVs (n = 3 for all groups, **P = 0.0074 COL1A1-LNPs vs Sham for IFN-γ; #P = 0.0333 COL1A1-EVs vs Sham and ***P < 0.001 COL1A1-LNPs vs Sham for IL-1β; ***P < 0.001 COL1A1-LNPs vs Sham for IL-6; *P = 0.0146 COL1A1-LNPs vs Sham for TNF-α; NS, not significant). e, Representative immunostaining images for TNF-α and (f) IL-6 after injected with COL1A1-EVs and COL1A1-LNPs. Scale bar, 100 µm. All data are from three independent experiments and are presented as means ± SEM. One-way ANOVA was used for the comparisons in (b–d).
Extended Data Fig. 4 Return of dermal wrinkles to baseline after treatment with low dose COL1A1-EVs.
a, Wrinkles were tracked on days 0, 4, 7, 14, 21, 28, 35, 42, 49, and 56 d after the indicated treatments (5 low-dose injections of COL1A1-EVs (2.7E9 copy number COL1A1 mRNA), COL1A1-LNPs (2.7E9 copy number COL1A1 mRNA), unloaded EVs, 0.05% retinoic acid [RA], saline). n = 4, Scale bar, 5 mm. Female nude mice that were not exposed to UV comprised the sham group. b, Numbers of wrinkles on the dorsal skin of the mice over time. (n = 4 for all groups; **P = 0.008 COL1A1-EVs vs Saline at day 7; **P = 0.004 COL1A1-EVs vs Saline at day 21, **P = 0.001 COL1A1-EVs vs Saline at day 35; ***P < 0.001 COL1A1-EVs vs Saline at days 14, 28, 42, and 49; †P = 0.025 RA vs Saline at day 21; ††P = 0.007 RA vs Saline at day 28; ##P = 0.0071 Unloaded EVs vs Saline at day 14; ##P = 0.004 Unloaded EVs vs Saline at day 21; ##P = 0.002 Unloaded EVs vs Saline at day 28; #P = 0.015 Unloaded EVs vs Saline at day 35; ※※P = 0.0053 COL1A1-LNPs vs Saline at day 14; ※※P = 0.0041 COL1A1-LNPs vs Saline at day 21; ※P = 0.017 COL1A1-LNPs vs Saline at day 28; ※P = 0.022 COL1A1-LNPs vs Saline at day 35). c, Total wrinkle area (n = 4 for all groups, *P = 0.012 COL1A1-EVs vs Saline at day 7; ***P < 0.001 COL1A1-EVs vs Saline at days 14, 21, 28 and 35; *P = 0.015 COL1A1-EVs vs Saline at day 42; ††P = 0.008 RA vs Saline at day 14; ††P = 0.007 RA vs Saline at days 21 and 35;††P = 0.005 RA vs Saline at day 28; #P = 0.012 Unloaded EVs vs Saline at day 14; #P = 0.035 Unloaded EVs vs Saline at day 21; ##P = 0.002 Unloaded EVs vs Saline at day 28; ※P = 0.062 COL1A1-LNPs vs Saline at day 14; ※P = 0.039 COL1A1-LNPs vs Saline at day 21; ※※P = 0.0027 COL1A1-LNPs vs Saline at day 28; ※※P = 0.046 COL1A1-LNPs vs Saline at day 35). All data are from three independent experiments and are presented as means ± SEM. Two-way ANOVA was used for the comparisons in (b, c).
Extended Data Fig. 5 Evaluation of long term COL1A1-EV MN dermal wrinkle treatment by skin replica plaster.
a–c, Microscopic observation of dorsal skin and skin replica at 1 month, 2 months, and 3 months after treatment. Scale bar, 5 mm. d, e Quantification of mean wrinkle length (n = 4 for all groups, ***P < 0.001 COL1A1-EV MN vs Saline at 1 month and 2 months; ###P < 0.001 Needle injection vs Saline at 1 month; ††P = 0.031 HA MN vs Saline at 1 month;※※P = 0.029 Unloaded EV MN vs Saline at 1 month) and mean wrinkle depth (n = 4 for all groups, ***P < 0.001 COL1A1-EV MN vs Saline at 1 month; **P = 0.001 COL1A1-EV MN vs Saline at 2 months; Needle injection vs Saline not significant at 1 month; HA MN vs Saline not significant at 1 month) from skin replicas. All data are from three independent experiments and are presented as means ± SEM. Two-way ANOVA was used for the comparisons in (d, e).
Extended Data Fig. 6 Maintenance of wrinkle treatment via serial injection of COL1A1-EVs and COL1A1-EV MN.
a, After 8 weeks of UV irradiated photoaging, wrinkles were tracked for mice treated every 30 days with 1) saline, 2) COL1A1-EVs, and 3) COL1A1-EV MN on days 0, 4, 7, 14, 21, 28, 49, 70, and 91 (COL1A1-EVs, COL1A1-EV MN, Saline). n = 4, Scale bar, 5 mm. b, Total wrinkle number (n = 4 for all groups; *P = 0.019 COL1A1-EV MN vs Saline at day 4; #P = 0.034 COL1A1-EVs vs Saline, *P = 0.031 COL1A1-EV MN vs Saline at day 7; #P = 0.030 COL1A1-EVs vs Saline, **P = 0.008 COL1A1-EV MN vs Saline at day 14; **P = 0.008 COL1A1-EV MN vs Saline at day 21; *P = 0.025 COL1A1-EV MN vs Saline at day 28; **P = 0.006 COL1A1-EV MN vs Saline at day 35; #P = 0.031 COL1A1-EVs vs Saline, *P = 0.030 COL1A1-EV MN vs Saline at day 42; #P = 0.044 COL1A1-EVs vs Saline, *P = 0.016 COL1A1-EV MN vs Saline at day 49; *P = 0.016 COL1A1-EV MN vs Saline at day 56; #P = 0.017 COL1A1-EVs vs Saline, *P = 0.020 COL1A1-EV MN vs Saline at day 63; *P = 0.018 COL1A1-EV MN vs Saline at day 70; #P = 0.011 COL1A1-EVs vs Saline, **P = 0.006 COL1A1-EV MN vs Saline at day 77; #P = 0.028 COL1A1-EVs vs Saline, *P = 0.037 COL1A1-EV MN vs Saline at day 84; #P = 0.043 COL1A1-EVs vs Saline, *P = 0.043 COL1A1-EV MN vs Saline at day 91) and c, wrinkle area on the dorsal skin of the mice during 90 day study window (n = 4 for all groups; #P = 0.012 COL1A1-EVs vs Saline, **P = 0.005 COL1A1-EV MN vs Saline at day7; #P = 0.010 COL1A1-EVs vs Saline, *P = 0.048 COL1A1-EV MN vs Saline at day 14; #P = 0.023 COL1A1-EVs vs Saline, *P = 0.021 COL1A1-EV MN vs Saline at day 21; *P = 0.022 COL1A1-EV MN vs Saline at day 28; *P = 0.046 COL1A1-EV MN vs Saline at day 35; #P = 0.019 COL1A1-EVs vs Saline, **P = 0.009 COL1A1-EV MN vs Saline at day 42; #P = 0.042 COL1A1-EVs vs Saline, *P = 0.030 COL1A1-EV MN vs Saline at day 49; *P = 0.030 COL1A1-EV MN vs Saline at day63; *P = 0.029 COL1A1-EV MN vs Saline at day 70; #P = 0.048 COL1A1-EVs vs Saline, *P = 0.022 COL1A1-EV MN vs Saline at day 77; #P = 0.040 COL1A1-EVs vs Saline, *P = 0.027 COL1A1-EV MN vs Saline at day 84; *P = 0.045 COL1A1-EV MN vs Saline at day 91). All data are from three independent experiments and are presented as means ± SEM. Two-way ANOVA was used for the comparisons in (b, c).
Supplementary information
Supplementary Information
Supplementary methods, results and discussion, materials, figures, tables and references.
Supplementary Video 1
Microneedles delivering HA EVs on mouse skin.
Supplementary Video 2
Ex vivo time course of the dissolution of the tips of microneedles delivering HA EVs into the skin.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
You, Y., Tian, Y., Yang, Z. et al. Intradermally delivered mRNA-encapsulating extracellular vesicles for collagen-replacement therapy. Nat. Biomed. Eng 7, 887–900 (2023). https://doi.org/10.1038/s41551-022-00989-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41551-022-00989-w
This article is cited by
-
Recent advances in extracellular vesicles for therapeutic cargo delivery
Experimental & Molecular Medicine (2024)
-
Unraveling the surface marker signature of cell-derived vesicles via proteome analysis and nanoparticle flow cytometry
Scientific Reports (2024)
-
Sirolimus-loaded exosomes as a promising vascular delivery system for the prevention of post-angioplasty restenosis
Drug Delivery and Translational Research (2024)
-
Engineered extracellular vesicles carrying let-7a-5p for alleviating inflammation in acute lung injury
Journal of Biomedical Science (2024)
-
mRNA-based vaccines and therapeutics: an in-depth survey of current and upcoming clinical applications
Journal of Biomedical Science (2023)
