
Abstract
In the BrighTNess trial, carboplatin added to neoadjuvant chemotherapy (NAC) was associated with increased pathologic complete response (pCR) rates in patients with stage II/III triple-negative breast cancer (TNBC). In this matched cohort study, cases with a germline BRCA1/2 mutation (gBRCA; n = 75) were matched 1:2 with non-gBRCA controls (n = 150) by treatment arm, lymph node status, and age to evaluate pCR rates and association of benefit from platinum/PARP inhibitors with validated RNA expression-based immune, proliferation, and genomic instability scores among gBRCA with the addition of carboplatin ± veliparib to NAC. Among the well-matched cohorts, odds of pCR were not higher in gBRCA cancers who received standard NAC with carboplatin (OR 0.24, 95% CI [0.04-1.24], p = 0.09) or with carboplatin/veliparib (OR 0.44, 95% CI [0.10-1.84], p = 0.26) compared to non-gBRCA cancers. Higher PAM50 proliferation, GeparSixto immune, and CIN70 genomic instability scores were each associated with higher pCR rate in the overall cohort, but not specifically in gBRCA cases. In this study, gBRCA carriers did not have higher odds of pCR than non-gBRCA controls when carboplatin ± veliparib was added to NAC, and showed no significant differences in molecular, immune, chromosomal instability, or proliferation gene expression metrics.
Similar content being viewed by others


Introduction
In localized triple-negative breast cancer (TNBC), neoadjuvant chemotherapy (NAC) allows for assessment of pathologic response to chemotherapy. Pathologic complete response (pCR) is associated with improved overall survival compared to residual disease1,2,3,4. To improve pCR rates, studies have attempted to add additional agents to the anthracycline plus taxane NAC backbone. Three studies have shown an improvement in pCR rate with the addition of carboplatin: GeparSixto5, CALGB 406036, and I-SPY27. Expanding on I-SPY2, BrighTNess was a phase III, multicenter, international, randomized, double-blind, placebo-controlled clinical trial, which enrolled 634 patients with stage II/III TNBC 2:1:1 to NAC with: (A) paclitaxel plus carboplatin plus veliparib followed by doxorubicin plus cyclophosphamide (TCV-AC), (B) paclitaxel plus carboplatin followed by doxorubicin plus cyclophosphamide (TC-AC), or (C) paclitaxel followed by doxorubicin plus cyclophosphamide (T-AC). In BrighTNess, the rate of pCR was higher in the TCV-AC (53%) and TC-AC (58%) arms, compared to T-AC (31%)8.
BRCA mutations result in impaired DNA damage repair by homologous recombination, which leads to sensitivity to PARP inhibitors and platinum chemotherapy in metastatic TNBC, as seen in the OlympiAD and TNT trials, respectively9,10. In the BrighTNess trial, 14–16% of patients in each arm had a confirmed deleterious germline BRCA1/2 mutation8. Interestingly, in the subgroup analysis of patients with germline BRCA mutations there was no difference in pCR between BRCA mutated and wildtype patients overall: 51% of patients with a BRCA mutation (gBRCA), achieved a pCR, compared to 48% of patients that did not harbor a BRCA mutation (non-gBRCA)8. There was a trend among patients with germline BRCA mutations toward higher pCR rates compared to T-AC alone (41%) with the addition of carboplatin to paclitaxel (50%), and a further trend toward higher pCR as veliparib was added to carboplatin and paclitaxel (57%)8. In non-gBRCA patients, pCR rates were 29% for T-AC alone, 59% with the addition of carboplatin, and 53% with the addition of veliparib with carboplatin8.
Recently, we published correlative genomic analysis of gene expression for 482 of 634 patients enrolled in BrighTNess11. We found that in multivariable analysis, proliferation and immune signatures were independently associated with pCR but that carboplatin benefit was not significantly different in basal-like vs. non-basal subgroups. Exploratory gene expression immune analyses suggested that tumors with higher inferred CD8+ T-cell infiltration may receive greater benefit with the addition of carboplatin11.
Based on these data, we hypothesized that patients with BRCA mutated TNBC in the BrighTNess trial may benefit differentially from the addition of carboplatin or veliparib to NAC and may have distinct immune gene expression profiles. To evaluate this, we analyzed the association of pCR rate with treatment arm in a matched cohort study of patients with germline BRCA1/2 (gBRCA) mutations compared to non-gBRCA controls. Using RNA sequencing of pre-treatment biopsies from the BrighTNess trial, we compared molecular subtype (PAM50 and TNBCtype), a measure of tumor proliferation (PAM50 proliferation score), a measure of chromosomal instability (CIN70), and measures of infiltrating immune cells (GeparSixto immune activation signature and relative immune cell abundance by TIMER) between gBRCA and non-gBRCA patients. Then, we determined if any of these measures were associated with pCR, which would indicate a subset of patients that would potentially benefit from the addition of carboplatin or veliparib.
Results
Matched cohort characteristics
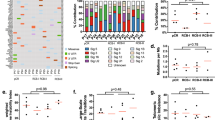
There were 75 BRCA1/2 mutated (gBRCA) cases and 150 non-gBRCA matched controls. BRCA cases and controls did not differ significantly by intentionally matched characteristics (planned treatment arm, lymph node status, age), nor by doxorubicin and cyclophosphamide (AC) administration schedule, Eastern Cooperative Oncology Group performance status, PAM50 subtype, TNBCtype, or rate of pCR (Table 1). Furthermore, BRCA cases did not differ from controls in terms of PAM50 proliferation score, GeparSixto immune activation signature score, CIN70 score, or TIMER-based relative abundance of tumor-infiltrating B cells, CD4 + T-cells, CD8 + T-cells, neutrophils, macrophages, or dendritic cells (Table 2).
BRCA status and treatment response
We assessed interactions between treatment arm and BRCA status. Similar to the overall BrighTNess analysis the entire cohort of mutated and non-mutated patients had higher pCR rates with the addition of carboplatin with or without veliparib compared to T-AC alone (Table 3). Arm A, TCV-AC, (odds ratio [OR]: 4.55; 95% confidence interval [CI]: [1.89, 10.97]) and Arm B, TC-AC, (OR: 5.31; 95%CI: [1.92, 14.66]) had significantly higher odds of pCR compared to Arm C, T-AC; however, there were no significant interactions between treatment arm and BRCA status for the prediction of pCR (Table 3). The pCR rates by treatment arm and BRCA status are shown in Supplementary Table 1. When stratifying by BRCA status, we found that it was the non-gBRCA, not the gBRCA, patients who had higher odds of pCR with the addition of carboplatin with or without veliparib compared to T-AC alone (Supplementary Table 2).
Gene expression features of gBRCA and non-gBRCA TNBCs
PAM50 proliferation score, GeparSixto immune activation signature score, and CIN70 score independently predict pCR (p = 0.007, 0.007, and 0.003, respectively), but there was no significant interaction between the scores and BRCA status (Table 4). Specifically, higher PAM50 proliferation score (OR: 3.14; 95% CI: [1.36, 7.23]), higher GeparSixto immune activation signature (OR: 1.67; 95% CI: [1.15, 2.44]), and higher CIN70 score (OR: 2.15; 95% CI: [1.29, 3.58]) were associated with an increased odds of pCR (Table 4). PAM50 subtype was not significantly associated with pCR. The study was underpowered to assess for association of TNBCtype with pCR. We assessed the associations between the proliferation, GeparSixto, and CIN70 scores, and found that proliferation score and CIN70 score were significantly, positively correlated (Supplementary Fig. 1).
As an exploratory, unbiased approach to broadly interrogate diverse pathways involved in cancer and the immune microenvironment, we performed single sample gene set enrichment analysis (GSEA) using Hallmark and Immune Response In Silico (IRIS)12 gene sets. None of the gene sets demonstrated significant differences between BRCA cases and controls after multiple test correction and only seven gene sets demonstrated nominal (p < 0.10) association with BRCA cases: WNT/Beta-catenin signaling (Hallmark), two neutrophil gene sets (IRIS), and two memory B-cell gene sets (IRIS) (Supplementary Tables 3 and 4).
Discussion
The role of carboplatin in NAC regimens for TNBC remains debated. While there is a clear increase in pCR rates with the addition of carboplatin to standard anthracycline-taxane chemotherapy, the long-term benefit of the addition of platinum is less clear5,6,8,13. This had led to uneven uptake of carboplatin in clinical practice. Among patients with germline BRCA mutations and metastatic breast cancer, the TNT, OlymiAD, and Brocade3 trials showed improved response rate and progression-free survival with the addition of carboplatin and/or PARP inhibitors9,10,14. This led to the hypothesis that patients with a germline BRCA mutation may specifically benefit from the addition of platinum chemotherapy and/or a PARP inhibitor to NAC. However, in the neoadjuvant setting, despite small studies suggesting a benefit of platinum chemotherapy in patients with BRCA mutations15,16, GeparSixto and INFORM showed no clear benefit with the addition of carboplatin or cisplatin, respectively17,18, reinforcing the need for greater understanding of the different results of metastatic trials versus neoadjuvant trials.
In this study, we present a large cohort of gBRCA patients compared with well-matched non-gBRCA controls, all of whom received uniform therapy as part of a randomized clinical trial consisting of standard NAC alone or with the addition of carboplatin with or without veliparib. This provides an opportunity to investigate the key questions of 1) whether matched patients with germline BRCA mutations benefit from the addition of carboplatin and/or PARP inhibitor; and 2) whether tumors with germline BRCA mutations have distinct proliferation, immune, and genomic instability gene expression signatures. This study builds on our initial analyses of RNAseq in this cohort that demonstrated carboplatin benefit was not significantly different in basal-like vs. non-basal subgroups but tumors with higher inferred CD8+ T-cell infiltration may receive greater benefit with addition of carboplatin in exploratory gene expression immune analyses11.
We found no significant difference in pCR between gBRCA and non-gBRCA patients among treatment arms in BrighTNess. This analysis does not include long-term follow up data and, notably, also does not include somatic BRCA mutations. We await event-free survival data from the BrighTNess study as well as phase III neoadjuvant trials in early-stage TNBC of NRG-BR003 (NCT024889967) comparing AC-T vs AC-TC as well as NSABP B-56 comparing VCT-AC to TC-AC and T-AC (NCT02032277). However, this cohort establishes that germline BRCA mutation carriers with TNBC do not receive further pCR benefit from the addition of carboplatin with or without veliparib to NAC. Intriguingly, the presence of a gBRCA mutation did not predict an increase in the pCR rate with the addition of either carboplatin or carboplatin and veliparib to neoadjuvant T-AC chemotherapy; in fact, it was in non-gBRCA patients that the addition of carboplatin with or without veliparib significantly increased the pCR rate. We hypothesize that with homologous recombination deficiency, the gBRCA tumors may be already intrinsically highly sensitive to standard anthracycline-taxane chemotherapy, offering less potential benefit with additional therapy. It should be noted that the data in this study do not have implications for the use of adjuvant PARP inhibitors for gBRCA carriers, related to the recently-published OlympiA study19.
The fact that the gBRCA and non-gBRCA cohorts were so well balanced allows for straightforward comparison of pCR in the two groups. However, it is surprising that the gBRCA and non-gBRCA cohorts in this study are so well matched by PAM50 subtype, TNBC subtype, PAM50 proliferation score, GeparSixto score, and proportions of subsets of tumor-infiltrating immune cells. Previous reports have shown that both BRCA1 and 2 tumors tend to be BL1 and BL2 TNBCtypes, BRCA1 tumors tend to be basal-like PAM50 subtype with high tumor inflammation signatures and many tumor-infiltrating immune cells, and BRCA2 tumors tend to be luminal A/B PAM50 subtype20,21,22,23,24,25.
In the cohort of both gBRCA and non-gBRCA patients, higher PAM50 proliferation score, CIN70 score, and GeparSixto immune signature were associated with higher odds of pCR, which is consistent with previous reports of association of pCR with high proliferation and immune signatures in breast cancer26,27,28,29, but inconsistent with the translational analysis of I-SPY2 which found that CIN70 did not predict response to veliparib with carboplatin30. The strong association found between CIN70 and proliferation reflects that the CIN70 score contains genes associated with proliferation and is consistent with CIN70 correlation with tumor grade31.
Recently, two studies demonstrate that the addition of immunotherapy to T-AC NAC alone32 or plus carboplatin33 further enhance pCR rates. As of yet, no subset analyses of germline BRCA patients from these studies have been reported. As we seek to personalize therapy for breast cancer patients, further research is warranted into the complex interplay between germline BRCA mutation, platinum chemotherapy or PARP inhibitors, and the immune microenvironment and immunotherapy.
This study does have limitations. This is an exploratory secondary analysis of clinical and genomic data from a phase III clinical trial and the power of the analysis may be limited by the small sample size. As these were not prespecified analyses, we pursued a matched cohort study design, which overcomes limitations of gBRCA vs. non-gBRCA studies where receptor subtype are frequently mixed and treatment heterogeneous, and attempted to control for covariates in multivariable analyses. Additionally, DNA metrics of HRD have not yet been fully analyzed (e.g. DNA based LOH scars, RAD51 focus formation, HRDetect mutational signatures), although preliminary analyses of Myriad HRD assay was not associated with differential benefit to addition of carboplatin alone or with veliparib34. Veliparib is established to have relatively less PARP trapping relative to other PARP inhibitors, which my facilitate combination with chemotherapy yet also limits our ability to extrapolate to other PARP inhibitors. Finally, recent FDA approval of immunotherapy as part of neoadjuvant therapy for TNBCs in the US reinforces the importance of validation studies in immunotherapy-containing cohorts.
In conclusion, in the overall cohort of both gBRCA and non-gBRCA patients, higher proliferation score, CIN70 score, and GeparSixto immune signature were associated with higher pCR rate and may be more useful biomarkers of patients who will benefit from the addition of carboplatin to neoadjuvant AC-T chemotherapy. The addition of carboplatin or carboplatin and veliparib to neoadjuvant T-AC chemotherapy was associated with increased pCR rate in non-gBRCA patients but not gBRCA carriers.
Methods
Study population and cohort selection
The BrighTNess trial (NCT02032277, registered January 10, 2014) enrolled 634 patients with stage II/III TNBC and was performed in accordance with The Code of Ethics of the World Medical Association (Declaration of Helsinki). Informed consent was obtained for all human subjects. Patients were randomized 2:1:1 to Arm A: TCV (paclitaxel 80 mg/m 2 IV weekly for 12 doses plus carboplatin AUC 6 IV every 3 weeks for four cycles plus veliparib 50 mg orally twice daily for 12 weeks); Arm B: TC (paclitaxel plus carboplatin plus veliparib placebo); or Arm C: T (paclitaxel plus carboplatin placebo plus veliparib placebo). Then all patients received AC every 2–3 weeks for four cycles, with the schedule selected by the treating physician. We matched non-gBRCA patients 2:1 (N = 150:75) to gBRCA cases by treatment arm, lymph node stage, and 10-year age range to balance the groups and reduce confounding. This cohort represents 225 of the previously assessed 482 patients11. The present analysis was not a preplanned analysis. Clinical data were locked as of January 1, 2018.
Ethical standards
The trial was performed in accordance with The Code of Ethics of the World Medical Association (Declaration of Helsinki) and was conducted according to the protocol approved by institutional review boards at investigational sites. The full protocol is available as a Supplemental file. Informed consent was obtained for all human subjects. This matched cohort study was approved by the Dana-Farber Cancer Institute Institutional Review Board.
Whole transcriptome gene expression analyses
A pre-treatment biopsy was collected in RNAlater. As detailed previously11, total RNA was extracted and underwent whole transcriptome RNA sequencing (RNAseq) on an Illumina HiSeq 3000 with single end 50 bp reads using RiboZero Gold rRNA depletion at the Washington University McDonnell Genome Institute. Samples with >10 million unique reads were included for further analyses as Reads per Kilobase per Million Reads (RPKM). RNA-seq reads were aligned to the Ensembl release 76 top-level assembly with STAR version 2.0.4b. Gene counts were derived from the number of uniquely aligned unambiguous reads by Subread:featureCount version 1.4.5. Transcript counts were produced by Sailfish version 0.6.3. All gene-level and transcript counts were then imported into the R/Bioconductor package EdgeR and TMM normalization size factors were calculated to adjust samples for differences in library size, resulting in RPKM which were used in downstream analyses. Genes or transcripts not expressed in any sample or less than one count-per-million in the minimum group size minus one were excluded from further analysis.
PAM50 subtype was determined with the ‘Bioclassifier’ package35 after balancing TNBC data with an equal number of estrogen receptor-positive cases from The Cancer Genome Atlas36. TNBCtype was determined with the TNBCtype tool after normalization to fixed upper quantile24,37. PAM50 ‘proliferation signature’ was derived from the ‘Bioclassifier’ package35. The GeparSixto immune signature of genes associated with tumor-infiltrating lymphocytes in GeparSixto38 and CIN70 signature of chromosomal instability31 were calculated as described previously. Proportions of infiltrating immune cell subsets were calculated using the TIMER algorithm39. Single sample Gene Set Enrichment Analysis (ssGSEA)40 was performed using Hallmark and Immune Response In Silico (IRIS)12/ImmuneSigDB41 gene sets.
Statistical analyses
pCR was determined by site pathologists following completion of neoadjuvant therapy, and was defined as absence of cancer cells in the breast and lymph nodes8. Chi-square tests were used to compare cohort characteristics. Student’s t-tests were used to compare the mean of continuous variables where the assumption of equal variances was met. For variables that did not meet this assumption, Welch’s t-tests were used. Logistic regression was performed to compare pCR with PAM50 subtype, proliferation score, GeparSixto score, CIN70 score, treatment arm, BRCA status, and interactions. T-tests were used to compare ssGSEA gene set scores, with multiple test correction using the method of Benjamini-Hochberg. Statistical analyses were performed with SAS version 9.4. Figures were generated with R version 3.5.1.
Reporting summary
Further information on research design is available in the Nature Research Reporting Summary linked to this article.
Data availability
All raw and processed sequencing files are uploaded and available through restricted access in Alliance Standardized Translational Omics Resource (A-STOR) with accession ASTOR_r6252020. Because the study did not meet submission requirements for dbGaP as a non-NIH funded study, the transcript abundance data, deidentified clinical data, and data dictionary are publicly available on NCBI GEO (accession GSE164458).
References
Cortazar, P. et al. Pathological complete response and long-term clinical benefit in breast cancer: the CTNeoBC pooled analysis. Lancet 384, 164–172 (2014).
von Minckwitz, G. et al. Definition and impact of pathologic complete response on prognosis after neoadjuvant chemotherapy in various intrinsic breast cancer subtypes. J. Clin. Oncol. 30, 1796–1804 (2012).
Liedtke, C. et al. Response to neoadjuvant therapy and long-term survival in patients with triple-negative breast cancer. J. Clin. Oncol. 26, 1275–1281 (2008).
Lehmann, B. D. et al. Refinement of triple-negative breast cancer molecular subtypes: implications for neoadjuvant chemotherapy selection. PLoS ONE 11, e0157368 (2016).
von Minckwitz, G. et al. Neoadjuvant carboplatin in patients with triple-negative and HER2-positive early breast cancer (GeparSixto; GBG 66): a randomised phase 2 trial. Lancet Oncol. 15, 747–756 (2014).
Sikov, W. M. et al. Impact of the addition of carboplatin and/or bevacizumab to neoadjuvant once-per-week paclitaxel followed by dose-dense doxorubicin and cyclophosphamide on pathologic complete response rates in stage II to III triple-negative breast cancer: CALGB 40603 (Alliance). J. Clin. Oncol. 33, 13–21 (2015).
Rugo, H. S. et al. Adaptive randomization of veliparib-carboplatin treatment in breast cancer. N. Engl. J. Med. 375, 23–34 (2016).
Loibl, S. et al. Addition of the PARP inhibitor veliparib plus carboplatin or carboplatin alone to standard neoadjuvant chemotherapy in triple-negative breast cancer (BrighTNess): a randomised, phase 3 trial. Lancet Oncol. 19, 497–509 (2018).
Robson, M. et al. Olaparib for metastatic breast cancer in patients with a germline BRCA mutation. N. Engl. J. Med. 377, 523–533 (2017).
Tutt, A. et al. Carboplatin in BRCA1/2-mutated and triple-negative breast cancer BRCAness subgroups: the TNT Trial. Nat. Med. 24, 628–637 (2018).
Filho, O. M., et al. Association of immunophenotype with pathologic complete response to neoadjuvant chemotherapy for triple-negative breast cancer: a secondary analysis of the brightness phase 3 randomized clinical trial. JAMA Oncol (2021).
Abbas, A. R. et al. Immune response in silico (IRIS): immune-specific genes identified from a compendium of microarray expression data. Genes Immun. 6, 319–331 (2005).
Poggio, F. et al. Platinum-based neoadjuvant chemotherapy in triple-negative breast cancer: a systematic review and meta-analysis. Ann. Oncol. 29, 1497–1508 (2018).
Dieras, V. C., et al. Phase 3 study of veliparib with carboplatin and paclitaxel in HER2-negative advanced/metastatic gBRCA-associated breast cancer. in ESMO 2019 Congress (2019).
Byrski, T. et al. Response to neoadjuvant therapy with cisplatin in BRCA1-positive breast cancer patients. Breast Cancer Res. Treat. 115, 359–363 (2009).
Telli, M. L. et al. Phase II study of gemcitabine, carboplatin, and iniparib as neoadjuvant therapy for triple-negative and BRCA1/2 mutation-associated breast cancer with assessment of a tumor-based measure of genomic instability: PrECOG 0105. J. Clin. Oncol. 33, 1895–1901 (2015).
Hahnen, E. et al. Germline mutation status, pathological complete response, and disease-free survival in triple-negative breast cancer: secondary analysis of the geparsixto randomized clinical trial. JAMA Oncol. 3, 1378–1385 (2017).
Tung, N., et al. Cisplatin versus doxorubicin/cyclophosphamide as neoadjuvant treatment in germline BRCA mutation carriers (BRCA carriers) with HER2-negative breast cancer: Results from the INFORM trial (TBCRC 031). in SABCS (San Antontio, TX, 2019).
Tutt, A. N. J. et al. Adjuvant olaparib for patients with BRCA1- or BRCA2-mutated breast cancer. N. Engl. J. Med. 384, 2394–2405 (2021).
Sorlie, T. et al. Repeated observation of breast tumor subtypes in independent gene expression data sets. Proc. Natl Acad. Sci. USA 100, 8418–8423 (2003).
Winter, C. et al. Targeted sequencing of BRCA1 and BRCA2 across a large unselected breast cancer cohort suggests that one-third of mutations are somatic. Ann. Oncol. 27, 1532–1538 (2016).
Ahn, S. G., Kim, S. J., Kim, C. & Jeong, J. Molecular classification of triple-negative breast cancer. J. Breast Cancer 19, 223–230 (2016).
Laenkholm, A. V., et al. Genomic profiling of tumors from patients with germline BRCA mutations. in ASCO (Chicago, IL, 2018).
Lehmann, B. D. et al. Identification of human triple-negative breast cancer subtypes and preclinical models for selection of targeted therapies. J. Clin. Invest 121, 2750–2767 (2011).
Wen, W. X. & Leong, C. O. Association of BRCA1- and BRCA2-deficiency with mutation burden, expression of PD-L1/PD-1, immune infiltrates, and T cell-inflamed signature in breast cancer. PLoS ONE 14, e0215381 (2019).
Stover, D. G. et al. The role of proliferation in determining response to neoadjuvant chemotherapy in breast cancer: a gene expression-based meta-analysis. Clin. Cancer Res. 22, 6039–6050 (2016).
Gluz, O. et al. Comparison of neoadjuvant Nab-Paclitaxel+Carboplatin vs nab-paclitaxel+gemcitabine in triple-negative breast cancer: randomized WSG-ADAPT-TN trial results. J. Natl Cancer Inst. 110, 628–637 (2018).
Prat, A. et al. Predicting response and survival in chemotherapy-treated triple-negative breast cancer. Br. J. Cancer 111, 1532–1541 (2014).
Gluz, O., et al. Association of molecular subtype, proliferation, and immune genes with efficacy of carboplatin versus gemcitabine addition to taxane-based, anthracycline-free neoadjuvant chemotherapy in early triple-negative breast cancer (TNBC): Results of the randomized WSG ADAPT-TN trial. in ASCO (Chicago, IL, 2017).
Wolf, D. M. et al. DNA repair deficiency biomarkers and the 70-gene ultra-high risk signature as predictors of veliparib/carboplatin response in the I-SPY 2 breast cancer trial. NPJ. Breast Cancer 3, 31 (2017).
Carter, S. L., Eklund, A. C., Kohane, I. S., Harris, L. N. & Szallasi, Z. A signature of chromosomal instability inferred from gene expression profiles predicts clinical outcome in multiple human cancers. Nat. Genet. 38, 1043–1048 (2006).
Mittendorf, E. A. et al. Neoadjuvant atezolizumab in combination with sequential nab-paclitaxel and anthracycline-based chemotherapy versus placebo and chemotherapy in patients with early-stage triple-negative breast cancer (IMpassion031): a randomised, double-blind, phase 3 trial. Lancet 396, 1090–1100 (2020).
Schmid, P. et al. Pembrolizumab for early triple-negative breast cancer. N. Engl. J. Med. 382, 810–821 (2020).
Telli, M. L. et al. Evaluation of homologous recombination deficiency (HRD) status with pathological response to carboplatin +/- veliparib in BrighTNess, a randomized phase 3 study in early stage TNBC. J. Clin. Oncol. 36, 519–519 (2018).
Parker, J. S. et al. Supervised risk predictor of breast cancer based on intrinsic subtypes. J. Clin. Oncol. 27, 1160–1167 (2009).
The Cancer Genome Atlas. Comprehensive molecular portraits of human breast tumours. Nature 490, 61–70 (2012).
Abkevich, V. et al. Patterns of genomic loss of heterozygosity predict homologous recombination repair defects in epithelial ovarian cancer. Br. J. Cancer 107, 1776–1782 (2012).
Denkert, C. et al. Tumor-infiltrating lymphocytes and response to neoadjuvant chemotherapy with or without carboplatin in human epidermal growth factor receptor 2-positive and triple-negative primary breast cancers. J. Clin. Oncol. 33, 983–991 (2015).
Di Meglio, A. et al. Time trends in incidence rates and survival of newly diagnosed stage IV breast cancer by tumor histology: a population-based analysis. Breast Cancer Res Treat. 157, 587–596 (2016).
Subramanian, A. et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl Acad. Sci. USA 102, 15545–15550 (2005).
Godec, J. et al. Compendium of immune signatures identifies conserved and species-specific biology in response to inflammation. Immunity 44, 194–206 (2016).
Acknowledgements
Research reported in this work was supported by the Alliance Foundation Trials; AbbVie Inc; Conquer Cancer Foundation of ASCO [Young Investigator Award to K.C.], Alliance Foundation [to D.G.S.], and Breast Cancer Research Foundation [to D.G.S.]. AbbVie was provided a draft of the manuscript prior to publication; however, they were not involved in the writing of the report or in the decision to submit the paper for publication. Dr. Metzger and Dr. Stover had full access to all the data in the study and had final responsibility for the decision to submit for publication. The content is solely the responsibility of the authors and does not necessarily represent the official views of the Alliance Foundation Trials, LLC. https://acknowledgments.alliancefound.org. The authors would like to acknowledge Catherine Carson, Celia Garr, Katherine Tyson, and Ashley Little for clinical support making this research possible. Otto Metzger-Filho, Katharine Collier, and Sarah Asad contributed equally. Charles E. Geyer Jr., Sibylle Loibl, and Daniel G. Stover jointly supervised this work.
Author information
Authors and Affiliations
Contributions
Conceptualization: O.M.F., K.C., S.A., P.J.A., C.E.G., S.L., D.G.S. Data curation: M.W., J.B., M.C. Formal Analysis: K.C., S.A., D.G.S. Funding acquisition: P.J.A., D.M., C.E.G., S.L. Investigation: J.O.S., M.U., H.S.R., J.B.H., M.G., W.M.S., G.V.M., P.R., N.W., C.D., W.F.S., C.E.G., S.L. Methodology: K.C., S.A., D.G.S. Project administration: D.G.S. Software: L.L., L.C. Supervision: O.M.F., C.E.G., S.L., D.G.S. Validation: D.G.S. Visualization: K.C., S.A., J.B., M.C., L.L., L.C., D.G.S. Writing—original draft: O.M.F., K.C., S.A., P.J.A., C.E.G., S.L., D.G.S. Writing—review & editing: All authors.
Corresponding author
Ethics declarations
Competing interests
C. Denkert reports stock and other ownership interests in Sividon Diagnostics (until 2016); honoraria from Novartis and Roche; consulting or advisory roles for MSD Oncology, Daiichi Sankyo, Molecular Health, AstraZeneca, and Merck; research funding (paid to institution) from Myriad Genetics and Roche; travel, accommodations, expenses from Roche; and patents, royalties, other intellectual property from VMScope digital pathology software (Patent applications WO2015114146A1 and WO2010076322A1- therapy response; Patent application WO2020109570A1 - cancer immunotherapy).
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Metzger-Filho, O., Collier, K., Asad, S. et al. Matched cohort study of germline BRCA mutation carriers with triple negative breast cancer in brightness. npj Breast Cancer 7, 142 (2021). https://doi.org/10.1038/s41523-021-00349-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41523-021-00349-y
This article is cited by
-
The impact of germline BRCA pathogenic variants in locally advanced, triple negative breast cancer treated with platinum-based neoadjuvant chemotherapy
Breast Cancer Research and Treatment (2024)
-
Post(neo)adjuvante Therapiekonzepte – Möglichkeiten der Therapieoptimierung
Der Gynäkologe (2022)
-
Outcomes of patients with HER2-negative metastatic breast cancer after platinum- and non-platinum-based first-line chemotherapy among patients with and without pathogenic germline BRCA1/2 mutations
British Journal of Cancer (2022)
-
Management of early-stage triple-negative breast cancer: recommendations of a panel of experts from the Brazilian Society of Mastology
BMC Cancer (2022)
