
Abstract
Much microbiome research has focused on populations that are predominantly of European descent, and from narrow demographics that do not capture the socio-economic and lifestyle differences which impact human health. Here we examined the airway microbiomes of the Orang Asli, the indigenous peoples of Malaysia. A total of 130 participants were recruited from two sites in the north-eastern state of Terengganu in Peninsular Malaysia. Using 16S rRNA sequencing, the nasal microbiome was significantly more diverse in those aged 5–17 years compared to 50+ years (p = 0.023) and clustered by age (PERMANOVA analysis of the Bray–Curtis distance, p = 0.001). Hierarchical clustering of Bray–Curtis dissimilarity scores revealed six microbiome clusters. The largest cluster (n = 28; 35.4%) had a marked abundance of Corynebacterium. In the oral microbiomes Streptococcus, Neisseria and Haemophilus were dominant. Using conventional microbiology, high levels of Staphylococcus aureus carriage were observed, particularly in the 18–65 age group (n = 17/36; 47.2% 95% CI: 30.9–63.5). The highest carriage of pneumococci was in the <5 and 5 to 17 year olds, with 57.1% (4/7) and 49.2% (30/61), respectively. Sixteen pneumococcal serotypes were identified, the most common being the nonvaccine-type 23A (14.6%) and the vaccine-type 6B (9.8%). The prevalence of pneumococcal serotypes covered by pneumococcal conjugate vaccines support introduction into a Malaysian national immunisation schedule. In addition, the dominance of Corynebacterium in the airway microbiomes is intriguing given their role as a potentially protective commensal with respect to acute infection and respiratory health.
Similar content being viewed by others


Introduction
Much microbiome research has, to-date, been focused on populations that are predominantly of European descent, and from demographics that do not capture the socio-economic and lifestyle challenges which impact human health1. The study of non-Western, unindustrialised populations is therefore an important extension of microbiome research so that we may understand human host–microbiota interactions in their broadest sense. Notable endeavours here, which have principally focussed on gut microbiomes, include studies of the Hadza hunter-gatherers of Tanzania2, Amerindians of South America3,4, agriculturalist communities in Burkina Faso5 Malawi and Venezuela6 and the Cheyenne and Arapaho Tribes of North America7. Fewer studies have gone beyond gut microbiomes to sample additional body sites, such as those of the airways. The analysis of salivary microbiota of the Batwa Pygmies8, the Yanomami of the Venezuelan Amazon3 and Guahibo of Venezuela9 have shown the benefits of including these body sites where, respectively, previously undiscovered genera have been identified and novel combinations of taxa described. However, there remains a paucity of data regarding microbiomes of other anatomical sites of the upper respiratory tract (URT). This lack of data requires addressing given the evidence that geographically isolated and/or indigenous populations endure distinct burdens of respiratory infections. For example children of Warao communities in Venezuela have been shown to be at increased risk of pneumococcal disease10, and it is well established that Aboriginal infants are highly susceptible to otitis media caused by Non-typeable Haemophilus influenzae (NTHi)11. With respiratory infectious diseases continuing to be a significant component of global morbidity and mortality12, the interest in the microbiome of the upper airways stems from its’ importance in an individual’s susceptibility to respiratory infection, in part through the presence of resilient taxa which prevent colonisation and/or outgrowth of specific pathobionts13. Understanding the variability in airway microbiomes is a key first step to leveraging these interactions for health. Data from the Gambia during the introduction of the seven valent pneumococcal conjugate vaccine (PCV7) highlighted changes in nasopharyngeal microbiome following implementation; a finding only possible due to the efforts to understand the microbial ecology as PCV7 was being rolled out14. To date no study has undertaken a comprehensive analysis of the URT microbiome of the indigenous populations of Peninsular Malaysia, the Orang Asli.
Although the name Orang Asli was only introduced around 1960 by the British, as an ethnic group they are believed to be the first settlers of Peninsular Malaysia, moving from Northern Thailand, Burma and Cambodia 3–8000 years ago. Orang Asli is in fact a catch-all term for three tribal groups, the Senoi, Proto-Malays and Negrito, each of which in turn can be further divided into six ethnic groups15. They number ~179,000, 0.6% of the population of Malaysia but are disproportionately impacted by economic marginalisation and discrimination16. Nearly 80% of the Orang Asli population are beneath the poverty line compared to 1.4% of the national population, and this hardship is reflected in a 20-year lower average life expectancy of just 53 years17. Several studies have highlighted the susceptibility of these populations to specific diseases18,19. Recently a study of 73 adults from the semi-urbanized Temiar, an Orang Asli tribe of Kampong Pos Piah, in Perak, examined salivary microbiomes in the context of obesity—a growing concern and evidence of the epidemiological shifts in disease burden as these communities leave their more traditional existences20. However, there is no published data on the rates and characteristics of respiratory tract infections in the Orang Asli.
Here we examined the microbiology of the upper airway of two Orang Asli communities located in Terengganu state, North-east of Peninsular Malaysia. The nasopharyngeal and nasal (anterior nares) carriage of important pathobionts was determined by culture, and the resistance of these isolates to clinically relevant antibiotics tested. We also present the first investigation of the nasal microbiomes of these indigenous peoples, in addition to their oral microbiomes.
Results
Participant demographics
A total of 130 participants were recruited to the study, with 68 from Kampung Sungai Pergam (Site 1) and 62 from Kampung Berua (Site 2) (Fig. 1). The age and gender distribution are shown in Fig. 1. Of those for whom age was accurately recorded, 49% were female and 45% male. Recruitment of children under five years old was low accounting for only 11.5% of the total population sampled, for which there was no obvious reason. All those with missing age data were adults. A comparison of participants from the two sites is shown in Table 1. A significant difference was seen for Site 1, which had a greater proportion of adults (18 to <85 year olds; p < 0.05) compared to Kampung Berua. In addition, self-reported respiratory illness was higher in both the previous month and 3 months prior to the site visits (p < 0.001). No difference in self-reported smoking or vaccination status was observed.

The location of Orang Asli settlements were in the north-east of Peninsula Malaysia (a) to the south of the Terengganu state’s capital city, Kuala Terengganu (b). An even gender split was achieved (c) but with disproportionate recruitment in older children >5 years of age and adults less than <65 (d).
Carriage of bacterial pathobionts
The carriage prevalence of the four most commonly isolated pathobionts in the anterior nares and nasopharynx are shown in Fig. 2. Here carriage was defined as the isolation of a pathobiont from either the nasal swab used for culture or the nasopharyngeal swab. The most commonly isolated pathobionts were Staphylococcus aureus and Streptococcus pneumoniae both with a total of 39 individuals colonised. The highest carriage for S. aureus was in the 18–65 age group at 47.2% (n = 17/36; 95% CI: 30.9–63.5), followed by 30% (n = 18/60, 95% CI: 18.4–41.6) in the 5 to 17 year olds. In contrast, the highest carriage of pneumococci was in the <5 and 5 to 17 year olds, with 57.1% (4/7) and 49.2% (30/61), respectively. Looking at the latter cases in more detail shows that the average age of a pneumococcal carrier in this age group was ~9 years old (range: 6–13). Examining differences in carriage of S. pneumoniae between female and male participants showed no significant differences (Supplementary Fig. 1). However, for S. aureus there was an increased carriage prevalence in females (57.7% (n = 15/36, 95% CI: 38.7–76.7) in the 18 to <65 years old age group, giving a significant (p = 0.0257) OR of 28.30, albeit it with large 95% CI: 1.50–534.25. The next most carried bacterium was H. influenzae, isolated from seventeen individuals, the majority of which (88.2%) were in the 5 to 17 years old age group. In contrast, Moraxella catarrhalis was rarely isolated (n = 6), as was Neisseria meningitidis (2), Pseudomonas aeruginosa (n = 3), Klebsiella pneumoniae (n = 6) and α-haemolytic Streptococci (n = 7). Carriage and co-carriage are summarised in Supplementary Table 1. Carriage of only S. aureus or S. pneumoniae accounted for 31.5% of the profiles observed. A limited number of co-carriage incidents were found, which included seven and five cases of pneumococci isolation with H. influenzae or S. aureus, respectively.

Point estimates for percentage carriage prevalence of four key upper respiratory tract pathobionts are shown across three age groups <5, 5 to 17 and 18 to 65. Those aged 65+ are not shown due to limited numbers. All ‘Unknowns” were of adult age. Carriage was estimated overall i.e. a positive swab culture from either nasal or nasopharyngeal swab (dark blue), and also separately for each swab type (nasal—black; NP—grey). Error bars show 95% CI intervals.
Pneumococcal serotypes
Serotyping of pneumococcal isolates revealed 16 different serotypes (Fig. 3). To avoid repeated sampling bias, if isolates recovered from nose and nasopharyngeal swabs in a single individual were the same serotype this was only counted once. The most common serotypes were the nonvaccine serotype (non-VT) 23A (14.6%) and the vaccine serotype (VT) 6B (9.8%). Overall, five of the 16 pneumococcal serotypes would be covered by Prevenar 7® (Pfizer, PCV7) (4, 6B, 14, 19F and 23F) with two additional serotypes covered by Prevenar 13® (Pfizer, PCV13) (3 and 6A). Only four of the serotypes (4, 14, 19F and 23F) are included in Synflorix® (GSK, PCV10). Six isolates were not serotypeable. All but one of the vaccine-type pneumococci (a single 23F) were isolated from the site at Kampung Berua (Site 2).

Bar plot with 95% CI showing prevalence of serotypes that are included in PCV7 (black), additional types covered by PCV13 (grey) or nonvaccine serotypes (white).
Antibiotic resistance
High levels of resistance were observed for S. aureus where 73.3% (n = 44) of isolates were resistant to penicillin, 33.0% (n = 20) to tetracycline and 18.3% (n = 11) to ciprofloxacin (Supplementary Fig. 2). However, all isolates were sensitive to cefoxitin, and all but one to chloramphenicol. Three of the H. influenzae isolates (15%) were resistant to penicillin, all of which were negative for β-lactamase activity. Of the S. pneumoniae, 17.9% (n = 12) were resistant to tetracycline. No resistance was seen for M. catarrhalis against any antibiotic tested. Acinetobacter baumannii were sensitive to meropenem and ciprofloxacin (n = 14). Similarly, all K. pneumoniae were sensitive to all antibiotics tested including ceftazidime and meropenem.
Upper respiratory tract microbiomes
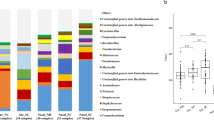
The microbiome of the nasal (anterior nares) and oral (whole mouth) was examined using 16S rRNA sequencing of the V4 hypervariable region. Here, the mean number of sequences per sample, excluding negative controls, was 64,694 with a maximum of 1,072,721. Negative control samples had a median of 1077 sequences, ranging from 692 to 1700. The frequency and classification of ASVs identified at phylum and genus level can be found in Supplementary Fig. 3. Sixteen nasal and two oral samples were excluded based on low numbers of ASVs (<1000). Eleven of these samples had ASVs only classified as Bacteria. The remaining five (comprising two oral and three nasal samples) also had a sequencing depth of <1000 reads. Alpha diversity (Fig. 4A, B) was similar between all ages apart from nasal samples from those aged 5–17 and 50–65 where a significantly greater diversity was observed in the younger group (p = 0.0023), although notably fewer samples were available from the older participants. Nasal samples are shown to cluster by age using DPCoA (Fig. 4C—left column) and PERMANOVA analysis of the Bray–Curtis distance showed that this was a significant difference (p = 0.001). In contrast, again using age as the grouping factor, this was not seen for oral samples (Fig. 4C—left column). Examining the most abundant phyla (Supplementary Fig. 4) showed nasal samples were dominated, as expected, by Firmicutes, Proteobacteria and Actinobacteria. For oral samples Bacteroidetes was a much more prevalent phyla in addition to Fusobacteria, although those same three phyla, which dominated the nasal samples were also present at high levels here as well. Hierarchical clustering of Bray–Curtis dissimilarity scores from nasal samples revealed six clusters that could be characterised by the dominance of only one or two genera (Fig. 5). The largest cluster (n = 28; 35.4%) contained individuals with a marked abundance of Corynebacterium, accounting for an average ASV relative abundance of 73.2% (range: 59.5–87.2%). The second largest cluster (n = 16; 20.3%) also had a substantial proportion of Corynebacterium, in this case with Dolosigranulum as significant co-carried genus. Here, on average, Corynebacterium accounted for 48.9% and Dolosigranulum 31.6% of the relative abundance; combined these Genera accounted for between 71.5 and 89.4% of the ASV relative abundance. Two clusters were then definable by the presence of Moraxella, one with and the other without Haemophilus. In the Moraxella-dominated cluster, between 48.2 and 75.2% of the ASV relative abundance could be attributed to this genus. When combined with Haemophilus this proportion dropped slightly from an average of 58.1 to 41.5% and here Haemophilus accounted for 13.2 to 47.9%. A small grouping of samples with Delftia/Ochrobactrum as the dominant genera were also found. The final cluster consisted of two distinct profiles, where in one Streptococcus accounted for 63.8% and the second with Streptococcus/Haemophilus at 25.9% and 52.6%, respectively.

Observed richness and Simpsons 1-D (a measure of diversity) are shown in panels a and b respectively. Only a significant difference (**) was observed for nasal samples for those aged 5–17 when compared against 50–65 (p = 0.0023). Beta diversity is shown using Double Principle Co-ordinates Analysis (DPCoA) (panel c). Here, clusters are observed for nasal samples (left) with grouping based on age group. No clusters were observed for oral samples (right).

The dendrogram (top) shows clustering of samples with the below bar chart showing the relative abundance of the eight most commonly observed Genera. Six clear clusters were observed including a Corynebacterium dominant profile (n = 28; 35.4%), a Corynebacterium/Dolosigranulum profile (n = 16; 20.3%), a Moraxella profile (n = 10; 12.7%) a Moraxella/Haemophilus profile (n = 10; 12.7%), a Delftia/Orchobactum profile (n = 4; 5.1%) and a final group of two samples characterised by high Streptococcus, in one case with Haemophilus. Culture results for S. aureus (dark blue) and S. pneumoniae (pink) are shown as coloured circles at the tips of the dendrogram. There is no clear distribution of individuals who were culture positive for either bacterium.
Co-occurrence of ASVs are shown in Fig. 6. Evidence of co-occurrence between ASVs of Haemophilus with Moraxella and Streptococcus can be seen, in particular the latter. There appears both negative and positive associations between Corynebacterium and Haemophilus. Dolosigranulum is linked to ASVs of both Moraxella and Corynebacterium. The Ochrobactrum-Delftia ASVs appear within a collection of nodes with other environmental taxa—further suggesting profiles dominated by these taxa (Fig. 5) are the result of contamination.

Node size represents the Log2 abundance of each ASV and are labelled by genus. Edge colours denote the direction of association with positive (green) and negative (red). Edge size shows strength of co-occurrence with a weighting value >2.
Double PCoA ordination and Jaccard distance-based network analyses were used to examine nasal microbiomes in the context of self-reported symptoms of respiratory illness (Supplementary Fig. 5). It was not possible to delineate samples based on symptomology given that there were significant differences between location. We further examined log2-fold changes in genera abundances using DESeq2 but found no significant taxa associated with individuals that reported URT illness within the previous one month or three months.
No correlation between microbiome composition, in terms of dominant genus, and culture outcomes for S. aureus or S. pneumoniae, as the two most frequently cultured pathobionts, were seen (Fig. 5). To determine if the abundance of either genus was related to culture positivity, the relative abundance of each was compared between samples from which either S. aureus or S. pneumoniae was isolated (Supplementary Fig. 6). The relative abundance of Staphylococcal ASVs was not significantly different between groups; however; those from whom S. pneumoniae were cultured did have a significantly higher relative abundance of Streptococci ASVs (p = 0.016); although this may be due to the presence of two profiles characterised by a significant dominance of Streptococcus. To explore the differences in microbiomes between the younger and older age groups, differentially abundant ASVs were identified using DESeq2 (Supplementary Fig. 7). The 5–17-year age group had ASVs classified as Haemophilus (including H. influenzae), Moraxella and Streptococcus. In contrast, Propionibacterium, Peptoniphilus and Corynebacterium ASVs were significantly increased in adults.
Streptococcus, Neisseria and Haemophilus dominated the oral microbiota (Supplementary Fig. 8). Lower levels of Prevotella, Rothia, Porphyromonas, Veillonella and Aggregatibacter were also among the eight most observed genera. Streptococcal-dominated profiles were characterised by 43.7% (±15.7%) relative abundance of ASVs belonging to this genus. Those with a Streptococcus/Haemophilus profile still had 30.7% (±7.6%) Streptococci ASV relative abundance but with 15.0% (±10%) Haemophilus. The final profile, where clear dominance of one particular genus was observed, were those where Neisseria accounted for 30.6% (±12.5) of the relative abundance of all ASVs.
Discussion
As a site that can be colonised by commensals, as well as opportunistic pathogens such as the pneumococcus21, S. aureus22 and H. influenzae23, understanding the microbiology of the upper respiratory tract is key in tackling respiratory disease. The Anna Karenina principle of microbiomes, that “all healthy microbiomes are similar; each dysbiotic microbiome is dysbiotic in its own way” is a useful maxim for examining microbiomes in the context of disease24. However, from previous studies of communities that live unique, traditional lifestyles a clear challenge to the paradigm of health mirroring health has been shown3,8. Here we examined the respiratory microbiomes of Orang Asli communities living traditional lifestyles in Terengganu, a rural state in the North-east of Peninsular Malaysia. These communities are known to face specific health challenges to that of the broader population of Malaysia, a country where respiratory disease is still a significant burden25. Our study showed a high prevalence of S. aureus carriage but interestingly not the presence of Staphylococci-dominated nasal microbiomes. Instead, Corynebacterium, with Dolosigranulum and Moraxella dominated as commensal flora. We also showed the potential impact that PCV introduction in Malaysia may have on circulating pneumococcal serotypes.
Although pneumococcal carriage can vary globally between 10 and 90%26,27,28, the highest carriage prevalence is usually seen in young children, particularly those <5 years28. Here, over half of the <5 year olds were colonised which is suggestive of high levels of carriage in this age group, although we note a very limited sample size (n = 7). Examining the next age group, 5–17, we observed a higher carriage than seen elsewhere in older children29. However, given the relatively young age of this cohort at ~9 years old, the higher carriage observed (50%; 95% CI, 37.3 and 62.7%) perhaps supports our proposition of high paediatric carriage. Of interest is that this level of carriage is similar to the 60.9% observed in Aboriginal children over 5 years old—a demographic known to experience high rates of invasive pneumococcal disease (IPD)30. As expected, the carriage in adults was low at 8.3% (95% CI, 0–17.4%). For context this is similar to the NP carriage prevalence of 11.1% (95% CI, 9.8 and 12.6%) seen in the USA31, slightly higher than recent surveys in the UK, which found a 2.8% (95%CI: 1.2–5.5) prevalence in this age group29 but significantly lower than that seen in Gambian adults where pneumococcal carriage exceeds 50% in all age groups32. As for serotypes, while there is growing epidemiological data for the region33, there is limited data for Malaysia. A study of 245 cases of paediatric IPD between 2014 and 2017 showed that serotypes 14, 6B, 19A, 6A and 19F were the most common, accounting for ~75% of disease34. This is concordant with this study where serotypes 14 and 6B were also the most common in carriage with 6A and 19F being identified. Of the non-VT serotypes isolated only 34, 35F and 18A did not feature in those identified in the IPD study. The lack of 19A is intriguing but we suspect a consequence of the relatively minimal sampling. Both PCV10 and PCV13 are available in Malaysia however only in private practice and therefore the presence of VT serotypes is not surprising. Given the presence of vaccine types here that have all but been eliminated from carriage in countries where public health initiatives have introduced these or similar vaccines these data do suggest introduction of PCV into the national immunisation programme as planned may prove efficacious.
Only one of the children less than 5 years old was culture positive for H. influenzae but given the low numbers in this age group this must be interpreted with caution. While certainly higher than in the UK where only 6.5% H. influenzae carriage was seen in older children35, the 25% (95% CI, 14.0 and 40.0%) found here is similar to that observed in other developing countries such as Kenya36, but lower than the 52.8% seen in Aboriginal children aged between 5 and 15 years old30. Whether this increased carriage translates into disease risk is difficult to determine; there is no data on the burden of otitis media among Orang Asli, although the country wide incidence in children <12 year olds is low at 2.3% for the Asia-Pacific region37. It is likely that there is not a burden of H. influenzae serotype B given the high coverage (98% in 2019) of the Hib vaccine in Malaysia38, although the lack of accurate health records for these communities renders this an assumption.
The prevalence of M. catarrhalis at 6.7% (95% CI, 0.35 and 13.0%) was very similar to that of our previous UK study where 5.7% of the same age group were colonised35, a level also observed in other western countries39. This is a stark contrast to the 67% in Aboriginal children of a similar age30; however, this low level of carriage (6%) has been seen in Warao Amerindians in Venezuela40—arguably a similar demographic to the one being presented here.
S. aureus carriage is known to vary markedly, by age but also by geographic location41. Although the lowest prevalence observed here was in <5 year olds (28.6%; 95% CI: 0–62.0), which is surprising as one would expect the highest carriage would be seen in this age group, it could be either a feature of the low recruitment or that the average age of this group was 2.5 years and carriage is known decline between 1 and 1022. As for the carriage in adults, a previous study in Malaysia of 384 adults (students with an average age of 25) found a carriage prevalence of 23.4%42, which is much lower than the 47.2% in our population of Orang Asli. A similarly high level of adult carriage (41.7% and 57.8% at two separate sampling timepoints) was observed in Wayampi Amerindians who, in terms of remote living, are similar to the Orang Asli43. Our own research has previously shown adult carriage at 24.4% in a UK population35 and thus the differences we observed are unlikely to be methodological. More likely it is a consequence of greater co-habitation (the average household size of our communities being ~5 people) and poorer sanitation infrastructure, both of which have been highlighted as reasons why S. aureus carriage has declined in other parts of the world22. It also is interesting that in our study females had significantly greater odds of carrying S. aureus which is in contrast to the typical burden in males44.
Of the ESKAPE pathogens isolated and tested only S. aureus exhibited any substantial level of resistance although the prevalence of resistance to Penicillin was in keeping with the most recent National Antibiotic Resistance Surveillance Report from 2018 where 80.6% of 34,564 isolates were also resistant45; importantly all isolates from this study were sensitive to methicillin. While there appears to be limited resistance it is important to note that this represents the only data on the antimicrobial resistance (AMR) in these communities and is therefore merely a starting point for future studies. This is particularly important given that AMR burden in indigenous populations has been seen to increase elsewhere46.
URT microbiome research has shown that the composition and/or dominance of particular bacterial taxa can be indicative of nasal community state types (CSTs). Previously, in a study of adults in the USA, seven CSTs were identified that were characterised by S. aureus (CST1), or other Staphylococci (CST3), a mix of Proteobacteria including Escherichia and Enterobacteriaceae sp. (CST2), Corynebacterium either with Propionibacterium acnes (CST4) or without (CST5), Moraxella (CST6) and Dolosigranulum (CST7)47. It is here that we see the most startling contrasts to out Orang Asli population. Although we also identified seven clusters based on what is/are the dominant taxa we did not see anything resembling CST1 or 3 (Staphylococci), and no CST2. Although we observed a high carriage prevalence of S. aureus, it is likely that this discordance with CSTs is, in part, an artefact of the ease with which this bacterium is cultured from nasal samples. Nevertheless, the lack of a dominant molecular signature is intriguing. Here we hypothesise that the high prevalence of profiles similar to CST5, Corynebacterium dominated, along with another that is defined by high abundance of both Corynebacterium and Dolosigranulum, a composition that has been found in the nasopharynx48, explains this absence and is based on previous work showing that Corynebacterium reduces S. aureus carriage49. A similar interaction may also explain the absence of Streptococci50—although we recognise in both cases that our hypothesis requires further testing due to the identification of both S. aureus and pneumococcal carriage among our participants. A Moraxella dominated profile similar to CST6, is also present in the Orang Asli, a profile which has also been observed in healthy adults in a European setting51, as well as in both Kenyan52 and Fijian children53 (where it is more common in iTaukei, the indigenous Fijians). We also saw a Moraxella-Haemophilus profile which hasn’t been described previously. The Streptococcus profiles are both from older children (6 and 10 years old) and this is not uncommon as a profile among this age group52,54. Given the low numbers it is impossible to determine whether we would have found more Streptococcus/Haemophilus profiles as been described elsewhere52. The significant proportion of individuals who harbour profiles that are being considered to be potentially protective either in terms of acute respiratory infections55 or chronic disease such as asthma56,57,58, is intriguing. Similar profiles were observed in Fijian children with either Dolosigranulum or Haemophilus/Corynebacterium dominant profiles53 although there are important contrasts, remembering that these were observed in older children and adults, we observed Corynebacterium and Corynebacterium/Dolosigranulum profiles in the Orang Asli. Lastly, while Delftia is not uncommon as an oral bacterium59, given the common finding of Delftia and Ochrobactrum as contaminants in microbiome research60 we interpret these four samples with extreme caution. The presence of these profiles is in spite of our efforts to control for such contaminants through the collection and sequencing of control swabs. Epidemiologically, all four samples come from adult females, but two each from Kampung Berua and Kampung Sungai Pergam. The samples are below the 25% quantile for ASV number (n = 3831). It is possible that these reflect true profiles, but it is impossible to rule out process error during sample handling, extraction and/or sequencing.
The microbiota recovered from the oral cavity was substantially more diverse than the nasal samples, as expected59. The top three genera were unremarkable in terms of what are considered to be most commonly observed, mirroring exactly that found in a study of 447 datasets deposited as part of large-scale microbiome projects61. A previous study of Temiar, Orang Asli in Perak, found a significant difference in oral microbiota compositions between males and females20; however, a PERMANOVA analysis of the data presented here showed no such distinction (p = 0.41).
While this piece of research was highly informative as a first glimpse into the respiratory microbiology of Orang Asli communities, it could nevertheless have been strengthened in a number of ways. First, it was only possible to visit two sites on a single occasion, and to conduct a pragmatic survey of the communities. As a consequence, the study lacks follow-up and longitudinal data. While we were very successful at recruiting older children and adults, future studies will have to identify how to recruit greater numbers of younger children in particular. In addition, those >65 years old were poorly represented as well. While we have included self-reported symptoms of respiratory infection, we have consciously avoided overinterpretation of these data. Health records are nonexistent for these communities, and there are suggestions that these communities suffer from a lack of adequate provision and/or are reluctant to seek out healthcare62, and thus it is impossible to determine their validity. It is intriguing that participants from Site 1 reported significantly greater incidences of respiratory infection, both in the preceding 1 and 3 months, and suggests community-specific health burdens. That this does not correlate to the greater incidence of pneumococcal serotypes associated with disease, and is not associated with any notable difference in microbiome composition in individuals from this particular site opens this as a question for future study. In terms of pathobiont carriage, serotyping of more than one pneumococcal isolate from each individual would have enabled a determination of the prevalence of multiple serotype carriage. Serotyping of H. influenzae would also have been informative, although future work on exploring the genomics of all isolates will address this in more detail. This additional insight could also be expanded to include all the pathogens/pathobionts isolated. Moreover, here we have relied on culture to determine carriage, which can be less sensitive than molecular methods as we have and others have shown previously35,63. As with all microbiome research, longitudinal sampling taking into account an individual’s temporal variability, the seasonality and medical history would be desirable. Finally, the lack of data on bacterial load leaves open the question, particularly with respect to nasal profiles, of whether this may be a factor in the differences we observed in clusters.
Important findings from this study include the prevalence of pneumococcal serotypes that would be covered by pneumococcal conjugate vaccines, supporting their introduction as part of a national immunisation programme. The high prevalence of S. aureus carriage is noteworthy and warrants further study. In terms of the airway microbiomes the dominance of Corynebacterium, additionally with Dolosigranulum and Moraxella in nasal profiles is particularly intriguing and future work should explore these commensals in the context of burden and susceptibility to both acute and chronic respiratory conditions in these communities.
Methods
Ethics
Ethical approval for this study was provided by Universiti Sultan Zainal Abidin (UniSZA) Ethics Committee: approval no. UniSZA/C/1/UHREC/628–1(85) dated 27 June 2016, the Department of Orang Asli Affairs and Development (JAKOA): approval no. JAKOA/PP.30.052Jld11 (42), and by the University of Southampton Faculty of Medicine Ethics Committee (Submission ID: 20831). Informed consent was taken for all participants, with parents/guardians providing consent for those <17 years old. Participation in the study involved reading (or being read) and understanding the Malay-translated participant information sheet, and the completion of a consent form and questionnaire. These documents were translated into Malay by A.S.M.H and A.R from the Faculty of Applied Social Sciences, Universiti Sultan Zainal Abidin who are familiar with the Orang Asli and have on-going social sciences-based research with them, and by M.A.R and C.C.Y from the Faculty of Medicine, Universiti Sultan Zainal Abidin, who are proficient in Malay and who ensured that the appropriate research and clinical context was preserved in the translated documents. The information sheet includes guidelines about the confidentiality and anonymity of the data obtained. This process was facilitated by native-speaking research assistants from the Faculty of Applied Social Sciences, Universiti Sultan Zainal Abidin, who underwent prior training on how to administer the questionnaires for the purpose of this study.
Study sites and participants
Two Orang Asli villages were visited in August 2017—Kampung Sungai Pergam in Kemaman district on the 8th August, and Kampung Berua in Hulu Terengganu district on the 9th August. Both sites are located in the state of Terengganu, which lies in the north-east of Peninsular Malaysia. Maps showing village locations were created using the R packages tmap64 and ggmap65. Participant recruitment was with consent, across all ages with no exclusion criteria. Questionnaires were used to capture participant metadata which included gender, age, number of dwelling co-occupants and occupation in addition to health-related questions including current or recent (within the last month and last three months) respiratory symptoms, antibiotic use and vaccination status.
Swab collection
A total of four swabs were taken from each participant on the day of visit to the study site. A nasal swab (anterior nares) and a nasopharyngeal swab, using rayon tipped transport swabs containing Amies media with charcoal (Medical Wire and Equipment, Corsham, UK), were used for bacterial pathobiont isolation. A further nasal swab, taken from the unswabbed nostril, and an oral (whole mouth) swab, supplied by uBiome Inc. (San Francisco, USA), were taken for microbiome analysis. For each study site visited four swabs (two each of nasal and oral) were opened as per the swabbing protocol but then placed, without use, into the uBiome Inc. transport containers for processing alongside used swabs as negative controls.
Bacteriology
Isolation of the common respiratory pathobionts S. pneumoniae, H. influenzae, M. catarrhalis, N. meningitidis, S. aureus, K. pneumoniae and P. aeruginosa was done by culture. Swabs were plated onto CBA (Columbia blood agar with horse blood), CHOC (Columbia blood agar with chocolated horse blood), CNA (Columbia Blood Agar with Colisitin and Naladixic Acid), BACH (Columbia Agar with Chocolated Horse Blood and Bacitracin), GC (Lysed GC Selective Agar) and, for P. aeruginosa, CFC (Pseudomonas CFC Selective agar) (all Oxoid, UK). Swabs were then vortexed in skim milk, tryptone, glucose and glycerol (STGG)66, and 10 μL of the suspension was then plated onto the appropriate media. Plates were then incubated for 24–48 h at 37 °C in 5% CO2. Primary identification of all bacterial species was by colonial morphology. Carriage for each pathobiont was then determined for age groups known to exhibit differences in prevalence: young children (aged <5 years), older children (>5 years to 17) and adults (18 to 65 years old).
Serotyping of S. pneumoniae
Pneumococcal isolates were serotyped by latex agglutination on a slide using a Neufeld S. pneumoniae antisera kit following manufacturer’s guidelines (Statens Serum Institute, Copenhagen, Denmark). Isolates were cultured from frozen STGG vials onto Columbia Blood agar (CBA) and incubated overnight at 37 °C in 5% CO2. A single colony was then purity plated onto a second CBA plate and again incubated overnight. Growth from this plate was inoculated into 3 ml of Todd-Hewitt Broth (THB) and incubated overnight at 37 °C in 5% CO2. Following o/n culture, cells were pelleted and resuspended in 100 µl THB. On a clean slide, 3 µl of the suspension was mixed with 3 µl of the pooled sera and examined for signs of agglutination. The chessboard for the identification of pneumococcal groups/types indicated if the isolate was a type (e.g. serotype 8) in which case no further serotyping was necessary, or a group (e.g. serogroup 15) in which case factor sera were used to further identify the serotype.
Antimicrobial resistance testing
All bacteria were phenotypically tested for antibiotic resistance using antibiotic discs and/or minimum inhibitory concentration (MIC) strips, in accordance with EUCAST. Firstly, 10 μL (a suspension of cells in liquid STGG) of each isolate was plated onto CBA (Oxoid, UK) or CHOC agar (Oxoid, UK). M. catarrhalis, S. pneumoniae, S. aureus and K. pneumoniae isolates were plated on CBA, while H. influenzae and N. meningitidis isolates were plated onto CHOC agar. Plates were incubated for 24 h at 37 °C in 5% CO2. Pure colonies were added to 1 ml of saline to get an inoculum of 0.5 McFarland. For M. catarrhalis, S. pneumoniae, H. influenzae and N. meningitidis, a sterile swab was used to spread this inoculum over Mueller–Hinton agar + 5% defibrinated horse blood and 20 mg−1 β-NAD plates (MHF, Oxoid, UK). For S. aureus and K. pneumoniae a sterile swab was used to spread the inoculum over Mueller–Hinton agar plates (MH, Oxoid, UK). Antibiotic discs (Oxoid, UK) (four per plate) or MIC strips (E-tests; Oxoid, UK) (one per plate) were added and plates were incubated at 37 °C in 5% CO2 for 18 hours (±2 hours).
M. catarrhalis were tested with amoxicillin–clavulanic acid (2–1 µg), cefotaxime (5 µg), ceftriaxone (30 µg), erythromycin (15 µg), tetracycline (30 µg), chloramphenicol (30 µg), ciprofloxacin (5 µg) and meropenem (10 µg) antibiotic discs. S. pneumoniae were tested with oxacillin (1 µg), erythromycin (15 µg), tetracycline (30 µg) and chloramphenicol (30 µg) antibiotic discs. H. influenzae were tested with benzylpenicillin (1 µg), tetracycline (30 µg), chloramphenicol (30 µg) and ciprofloxacin (5 µg) antibiotic discs as well as erythromycin (15 µg) MIC strips. S. aureus were tested with benzylpenicillin (1 µg), erythromycin (15 µg), cefoxitin (30 µg), tetracycline (30 µg), chloramphenicol (30 µg) and ciprofloxacin (5 µg) antibiotic discs. K. pneumoniae were tested with amoxicillin–clavulanic acid (20–10 µg), cefotaxime (5 µg), ciprofloxacin (5 µg), meropenem (10 µg) and ceftazidime (10 µg) antibiotic discs. N. meningitidis were tested with amoxicillin, benzylpenicillin (1 µg), cefotaxime (5 µg), ceftriaxone (30 µg), chloramphenicol (30 µg), ciprofloxacin (5 µg) and meropenem (10 µg) MIC strips.
16S rRNA sequencing
The V4 region was amplified using primers 515 F (5′-GTGCCAGCMGCCGCGGTAA-3′) and 806 R (5′-GGACTACHVGGGTWTCTAAT-3′) to generate an amplicon of 460 bp67 at uBiome Inc. (San Francisco, USA). Samples were individually barcoded and sequenced on a NextSeq 500 (Illumina, San Diego, USA) to generate 2 × 150 bp paired-end reads. Reads were supplied having undergone primer and low-quality end trimming. Maximum read lengths for forward and reverse were capped at 125 and 124 bp, respectively.
Microbiome analysis
Initial data handling was done using QIIME 2 2018.868. Only forward reads were processed owing to additional low-quality scores (<Q30) of reverse reads and that merging of read pairs resulted in significant loss of data. Raw sequence data were denoised with DADA269. Amplicon sequence variants (ASVs) were aligned with mafft70 (via q2‐alignment) and a phylogeny constructed using fasttree271. Taxonomic classification was done using the q2‐feature‐classifier72 classify‐sklearn naïve Bayes taxonomy classifier using the Greengenes 13_8 (99%) reference sequences73. The feature table, taxonomy, phylogenetic tree and sample metadata were then combined into a Phyloseq object using qiime2R74 via qza_to_phyloseq. All further analysis was done in R v3.6.075 in RStudio and figures were produced using the package ggplot276. Phyloseq v1.29.077 was used following a published workflow78. Potential contaminants were identified by prevalence in the blank swab controls and removed using the R package ‘decontam’79. Ten ASVs were removed as a result. Only one was identified beyond “Bacteria” and belonged to the Genus Meiothermus. In addition, ASVs with Phylum classifications of ‘NA’ or ‘uncharacterised’ and any Phyla with a total of fewer than five ASVs whose provenance suggested low level environmental contamination were removed using the subset_taxa() function. Finally ASVs with Kingdom classifications other than ‘Bacteria’, Phylum classifications of ‘Cyanobacteria’, Family classifications of ‘mitochondria’ and Class classifications of ‘Chloroplast’ were also removed. Samples with <1000 ASVs were excluded and taxa present in less than 5% of samples were removed using the prune_taxa() phyloseq function. Alpha diversity was calculated using the estimate_richness(). The R function stat_compare_means() was used to compare age groups using the nonparametric Wilcoxon test. Beta diversity was determined using Double Principle Co-ordinates Analysis and plot_ordination(). Hierarchical clustering was done using Bray–Curtis Dissimilarity calculated using vegist() from the R package vegan80. Differentially abundant ASVs were identified using the DESeq281 package using an adjusted p-value cut-off of 0.05 and a Log2 fold change of 1.5. Networks were generated using the R implementation of SPIEC-EASI (SpiecEasi)82 using the meinshausen-buhlmann’s neighbourhood selection method with results visualised using ggnet().
Reporting summary
Further information on research design is available in the Nature Research Reporting Summary linked to this article.
Data availability
Sequence files and associated metadata have been deposited in the European Nucleotide Archive (ENA) in project PRJEB38610 under experiment accessions ERX4147052 to ERX4147288.
Code availability
Code is available at: https://github.com/davidwcleary/orang_asli_16S/blob/master/OrangAsli.Rmd
References
Rogers, G. B., Ward, J., Brown, A. & Wesselingh, S. L. Inclusivity and equity in human microbiome research. Lancet 393, 728–729 (2019).
Schnorr, S. L. et al. Gut microbiome of the Hadza hunter-gatherers. Nat. Commun. 5, 3654 (2014).
Clemente, J. C. et al. The microbiome of uncontacted Amerindians. Sci. Adv. 3, e1500183 (2015).
Obregon-Tito, A. J. et al. Subsistence strategies in traditional societies distinguish gut microbiomes. Nat. Commun. 6, 6505 (2015).
Filippo, C. et al. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl Acad. Sci. USA https://doi.org/10.1073/pnas.1005963107 (2010).
Yatsunenko, T. et al. Human gut microbiome viewed across age and geography. Nature 486, 222–227 (2012).
Sankaranarayanan, K. et al. Gut microbiome diversity among cheyenne and arapaho individuals from Western Oklahoma. Curr. Biol. 25, 3161–3169 (2015).
Nasidze, I. et al. High Diversity of the saliva microbiome in Batwa Pygmies. PLoS ONE 6, e23352 (2011).
Contreras, M. et al. The bacterial microbiota in the oral mucosa of rural Amerindians. Microbiology 156, 3282–3287 (2010).
Rivera-Olivero, I. et al. Pneumococcal carriage among indigenous Warao children in Venezuela: serotypes, susceptibility patterns, and molecular epidemiology. Clin. Infect. Dis. 45, 1427–1434 (2007).
Sun, W. et al. Association between early bacterial carriage and otitis media in Aboriginal and non-Aboriginal children in a semi-arid area of Western Australia: a cohort study. BMC Infect. Dis. 12, 366 (2012).
Lozano, R. et al. Global and regional mortality from 235 causes of death for 20 age groups in 1990 and 2010: a systematic analysis for the Global Burden of Disease Study 2010. Lancet 380, 2095–2128 (2012).
Man, W. H., de Steenhuijsen Piters, W. A. A. & Bogaert, D. The microbiota of the respiratory tract: gatekeeper to respiratory health. Nat. Rev. Microbiol. 15, 259–270 (2017).
Kwambana-Adams, B. et al. Rapid replacement by non-vaccine pneumococcal serotypes may mitigate the impact of the pneumococcal conjugate vaccine on nasopharyngeal bacterial ecology. Sci. Rep. 7, 8127 (2017).
SyedHussain, T., Krishnasamy, D. S. & Hassan, A. A. G. Distribution and demography of the Orang Asli in Malaysia. Int. J. Humanities Soc. Sci. Invent. 6, 40–45 (2017).
Masron, T., Ismail, M. F. & Orang, N. Asli in Peninsular Malaysia: population, spatial distribution and socio-economic condition. J. Ritsumeikan Soc. Sci. Humanit 6, 75–115 (2013).
Mohd Asri, M. N. Advancing the orang Asli through Malaysia clusters of excellence policy. J. Int. Comp. Edu. https://doi.org/10.14425/00.45.76 (2012).
Al-Mekhlafi, H. M. et al. Prevalence and risk factors of Strongyloides stercoralis infection among Orang Asli schoolchildren: new insights into the epidemiology, transmission and diagnosis of strongyloidiasis in Malaysia. Parasitology 146, 1602–1614 (2019).
Al-Mekhlafi, H. M. et al. Protein-energy malnutrition and soil-transmitted helminthiases among Orang Asli children in Selangor, Malaysia. Asia Pac. J. Clin. Nutr. 14, 188–194 (2005).
Yeo, L.-F., Aghakhanian, F. F., Tan, J. S. Y., Gan, H. M. & Phipps, M. E. Health and saliva microbiomes of a semi-urbanized indigenous tribe in Peninsular Malaysia. F1000Res. 8, 175–175 (2019).
Bogaert, D., de Groot, R. & Hermans, P. W. M. Streptococcus pneumoniae colonisation: the key to pneumococcal disease. Lancet Infect. Dis. 4, 144–154 (2004).
Wertheim, H. F. L. et al. The role of nasal carriage in Staphylococcus aureus infections. Lancet Infect. Dis. 5, 751–762 (2005).
Slack, M. P. E. A review of the role of Haemophilus influenzae in community-acquired pneumonia. Pneumonia. https://doi.org/10.15172/pneu.2015.6/520 (2015).
Zaneveld, J. R., McMinds, R. & Vega Thurber, R. Stress and stability: applying the Anna Karenina principle to animal microbiomes. Nat. Microbiol. 2, 17121 (2017).
Malaysia DoS (Department of Statistics Malaysia). www.dosm.gov.my (2018).
Hill, P. C. et al. Nasopharyngeal carriage of Streptococcus pneumoniae in gambian infants: a longitudinal study. Clin. Infect. Dis. 46, 807–814 (2008).
Regev-Yochay, G. et al. Nasopharyngeal carriage of Streptococcus pneumoniae by adults and children in community and family settings. Clin. Infect. Dis. 38, 632–639 (2004).
Bogaert, D. et al. Colonisation by Streptococcus pneumoniae and Staphylococcus aureus in healthy children. Lancet. https://doi.org/10.1016/s0140-6736(04)16357-5 (2004).
Southern, J. et al. Pneumococcal carriage in children and their household contacts six years after introduction of the 13-valent pneumococcal conjugate vaccine in England. PLoS ONE 13, e0195799 (2018).
Mackenzie, G. A., Leach, A. J., Carapetis, J. R., Fisher, J. & Morris, P. S. Epidemiology of nasopharyngeal carriage of respiratory bacterial pathogens in children and adults: cross-sectional surveys in a population with high rates of pneumococcal disease. BMC Infect. Dis. 10, 304 (2010).
Watt, J. P. et al. Nasopharyngeal versus oropharyngeal sampling for detection of Pneumococcal Carriage in adults. J. Clin. Microbiol. 42, 4974–4976 (2004).
Hill, P. C. et al. Nasopharyngeal carriage of Streptococcus pneumoniae in gambian villagers. Clin. Infect. Dis. 43, 673–679 (2006).
Jauneikaite, E., Jefferies, J. M., Hibberd, M. L. & Clarke, S. C. Prevalence of Streptococcus pneumoniae serotypes causing invasive and non-invasive disease in South East Asia: a review. Vaccine 30, 3503–3514 (2012).
Arushothy, R. et al. Pneumococcal serotype distribution and antibiotic susceptibility in Malaysia: a four-year study (2014–2017) on invasive paediatric isolates. Int. J. Infect. Dis. 80, 129–133 (2019).
Coughtrie, A. L. et al. Ecology and diversity in upper respiratory tract microbial population structures from a cross-sectional community swabbing study. J. Med. Microbiol. 67, 1096–1108 (2018).
Abdullahi, O., Nyiro, J., Lewa, P., Slack, M. & Scott, J. A. G. The descriptive epidemiology of Streptococcus pneumoniae and Haemophilus influenzae nasopharyngeal carriage in children and adults in Kilifi district, Kenya. Pediatr. Infect. Dis. J. 27, 59–64 (2008).
Mahadevan, M. et al. A review of the burden of disease due to otitis media in the Asia-Pacific. Int. J. Pediatr. Otorhinolaryngol. 76, 623–635 (2012).
WHO. WHO UNICEF estimates time series for Malaysia (MYS). https://apps.who.int/immunization_monitoring/globalsummary/estimates?c=MYS (2020).
Kovács, E. et al. Co-carriage of Staphylococcus aureus, Streptococcus pneumoniae, Haemophilus influenzae and Moraxella catarrhalis among three different age categories of children in Hungary. PLoS ONE 15, e0229021 (2020).
Verhagen, L. M. et al. Nasopharyngeal carriage of respiratory pathogens in Warao Amerindians: significant relationship with stunting. Tropical Med. Int. Health 22, 407–414 (2017).
Sollid, J. U. E., Furberg, A. S., Hanssen, A. M. & Johannessen, M. Staphylococcus aureus: determinants of human carriage. Infect. Genet. Evol. 21, 531–541 (2014).
Choi, C. S. et al. Nasal carriage of Staphylococcus aureus among healthy adults. J. Microbiol. Immunol. Infect. 39, 458–464 (2006).
Ruimy, R. et al. Are host genetics the predominant determinant of persistent nasal Staphylococcus aureus carriage in humans? J. Infect. Dis. 202, 924–934 (2010).
Humphreys, H., Fitzpatick, F. & Harvey, B. J. Gender differences in rates of carriage and bloodstream infection caused by Methicillin-Resistant Staphylococcus aureus: are they real, do they matter and why? Clin. Infect. Dis. 61, 1708–1714 (2015).
Hashim, R. National Antibiotic Resistance Surveillance Report 2018 (Antibiotic Resistance Surveillance Reference Laboratory, Institute for Medical Research 2018).
Tong, S. Y. C., Varrone, L., Chatfield, M. D., Beaman, M. & Giffard, P. M. Progressive increase in community-associated methicillin-resistant Staphylococcus aureus in Indigenous populations in northern Australia from 1993 to 2012. Epidemiol. Infect. 143, 1519–1523 (2015).
Liu, C. M. et al. Staphylococcus aureus and the ecology of the nasal microbiome. Sci. Adv. https://doi.org/10.1126/sciadv.1400216 (2015).
Cremers, A. J. et al. The adult nasopharyngeal microbiome as a determinant of pneumococcal acquisition. Microbiome 2, 1–10 (2014).
Yan, M. et al. Nasal microenvironments and interspecific interactions influence nasal microbiota complexity and S. aureus carriage. Cell Host Microbe 14, 631–640 (2013).
Bomar, L., Brugger, S. D., Yost, B. H., Davies, S. S. & Lemon, K. P. Corynebacterium accolens Releases Antipneumococcal Free Fatty Acids from Human Nostril and Skin Surface Triacylglycerols. mBio. https://doi.org/10.1128/mBio.01725-15 (2016).
De Boeck, I. et al. Comparing the healthy nose and nasopharynx microbiota reveals continuity as well as niche-specificity. Front. Microbiol. 8, 2372–2372 (2017).
Feazel, L. M. et al. Effects of vaccination with 10-vlent Pneumococcal non-typeable Haemophilus influenza protein D conjugate vaccine (PHiD-CV) on the nasopharyngeal microbiome of Kenyan Toddlers. PLoS ONE 10, e0128064–e0128064 (2015).
Boelsen, L. K. et al. The association between pneumococcal vaccination, ethnicity, and the nasopharyngeal microbiota of children in Fiji. Microbiome 7, 106–106 (2019).
Biesbroek, G. et al. Early respiratory microbiota composition determines bacterial succession patterns and respiratory health in children. Am. J. Respir. Crit. Care Med. https://doi.org/10.1164/rccm.201407-1240OC (2014).
Laufer, A. S. et al. Microbial communities of the upper respiratory tract and otitis media in children. mBio. https://doi.org/10.1128/mBio.00245-10 (2011).
Teo Shu, M. et al. The infant nasopharyngeal microbiome impacts severity of lower respiratory infection and risk of asthma development. Cell Host Microbe. https://doi.org/10.1016/j.chom.2015.03.008 (2015).
Durack, J., Boushey, H. A. & Lynch, S. V. Airway microbiota and the implications of dysbiosis in asthma. Curr. Allergy Asthma Rep. 16, 52 (2016).
Ege, M. J. et al. Exposure to environmental microorganisms and childhood asthma. N. Engl. J. Med. https://doi.org/10.1056/NEJMoa1007302 (2011).
Verma, D., Garg, P. K. & Dubey, A. K. Insights into the human oral microbiome. Arch. Microbiol. 200, 525–540 (2018).
Salter, S. J. et al. Reagent and laboratory contamination can critically impact sequence-based microbiome analyses. BMC Biol. https://doi.org/10.1186/s12915-014-0087-z (2014).
Wang, J., Li, D., Wang, J. & Zhang, Z. Human oral microbiome characterization and its association with environmental microbiome revealed by the Earth Microbiome Project. bioRxiv. https://doi.org/10.1101/732123 (2019).
Wong, Y. S., Allotey, P. & Reidpath, D. D. Why we run when the doctor comes: Orang Asli responses to health systems in transition in Malaysia. Crit. Public Health 29, 192–204 (2019).
Olwagen, C. P., Adrian, P. V. & Madhi, S. A. Comparison of traditional culture and molecular qPCR for detection of simultaneous carriage of multiple pneumococcal serotypes in African children. Sci. Rep. 7, 4628 (2017).
Tennekes, M. tmap: thematic Maps in R. J. Stat. Softw. https://doi.org/10.18637/jss.v084.i06 (2018).
Kahle, D. W. H. & Wickham, H. ggmap: spatial visualization with ggplot2. R. J. 5, 144–161.
O’Brien, K. L. et al. Evaluation of a medium (STGG) for transport and optimal recovery of Streptococcus pneumoniae from nasopharyngeal secretions collected during field studies. J. Clin. Microbiol. 39, 1021–1024 (2001).
Almonacid, D. E. et al. 16S rRNA gene sequencing and healthy reference ranges for 28 clinically relevant microbial taxa from the human gut microbiome. PLoS ONE 12, e0176555 (2017).
Bolyen, E. et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 37, 852–857 (2019).
Callahan, B. J. et al. DADA2: high-resolution sample inference from Illumina amplicon data. Nat. Methods. https://doi.org/10.1038/nmeth.3869 (2016).
Katoh, K., Misawa, K., Kuma, K.-I. & Miyata, T. MAFFT: a novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 30, 3059–3066 (2002).
Price, M. N., Dehal, P. S. & Arkin, A. P. FastTree 2--approximately maximum-likelihood trees for large alignments. PLoS ONE. https://doi.org/10.1371/journal.pone.0009490 (2010).
Bokulich, N. A. et al. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome 6, 90 (2018).
McDonald, D. et al. An improved Greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 6, 610–618 (2012).
Bisanz, J. E. qiime2R: Importing QIIME2 artifacts and associated data into R sessions. https://github.com/jbisanz/qiime2R (2018).
Team, R. C. R: a language and environment for statistical computing. (R Foundation for Statistical Computing, 2014).
Wickham, H. ggplot2: Elegant Graphics for Data Analysis (Springer-Verlag, New York, 2016).
McMurdie, P. J. H. S. phyloseq: an R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE. https://doi.org/10.1371/journal.pone.0061217 (2013).
Callahan, B., Sankaran, K., Fukuyama, J., McMurdie, P. & Holmes, S. Bioconductor Workflow for Microbiome Data Analysis: from raw reads to community analyses [version 2; peer review: 3 approved]. F1000Res. https://doi.org/10.12688/f1000research.8986.2 (2016).
Davis, N. M., Proctor, D. M., Holmes, S. P., Relman, D. A. & Callahan, B. J. Simple statistical identification and removal of contaminant sequences in marker-gene and metagenomics data. Microbiome 6, 226 (2018).
Oksanen, J. et al. Vegan: Community Ecology Package. R package version 2. 5–7. https://CRAN.R-project.org/package=vegan (2020).
Love, M. I., Huber, W., Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. https://doi.org/10.1186/s13059-014-0550-8 (2014).
Kurtz, Z. D. et al. Sparse and compositionally robust inference of microbial ecological networks. PLoS Comput. Biol. 11, e1004226 (2015).
Acknowledgements
The laboratory microbiology work was funded by a grant from the Higher Education Funding Council for England (HEFCE) Newton Fund Official Development Assistance (ODA) fund. The microbiome sequencing was funded through a uBiome Inc. Microbiome Grants Initiative award. We would like to thank the Department of Orang Asli Affairs and Development (JAKOA), Additionally, the authors would especially like to thank the communities of Kampung Sungai Pergam and Kampung Berua for their support and participation in this study.
Author information
Authors and Affiliations
Contributions
D.W.C. and S.C.C. conceived the project with Y.C.C. D.E., R.A.A. and H.M. were the UoS contingent who undertook the sampling and helped with study planning and design. A.G.A., N.I.A.R., S.I., M.S.R., R.M.A., A.A.A., N.K.E., S.A., C.H.C. H.S. and R.A. helped with study design, coordinated sampling visits to Orang Asli settlements, liaised with those communities and assisted with sampling and questionnaires. A.G.A. did the microbiological culture and antibiotic testing with R.A.A. and D.E. D.W.C. carried out the data analysis and wrote the paper. All authors provided critical feedback and helped shape the manuscript.
Corresponding author
Ethics declarations
Competing interests
S.C.C. acts as principal investigator on studies conducted on behalf of University Hospital Southampton NHS Foundation Trust/University of Southampton that are sponsored by vaccine manufacturers but receives no personal payments from them. S.C.C. has participated in advisory boards for vaccine manufacturers but receives no personal payments for this work. S.C.C. has received financial assistance from vaccine manufacturers to attend conferences. D.W.C. was a post-doctoral researcher on projects funded by Pfizer and GSK between April 2014 and October 2017. All grants and honoraria are paid into accounts within the respective NHS Trusts or Universities, or to independent charities. All other authors have no conflicts of interest.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Cleary, D.W., Morris, D.E., Anderson, R.A. et al. The upper respiratory tract microbiome of indigenous Orang Asli in north-eastern Peninsular Malaysia. npj Biofilms Microbiomes 7, 1 (2021). https://doi.org/10.1038/s41522-020-00173-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41522-020-00173-5
This article is cited by
-
Genome-wide mapping of gene-microbe interactions in the murine lung microbiota based on quantitative microbial profiling
Animal Microbiome (2023)
-
Cooperative microbial interactions drive spatial segregation in porous environments
Nature Communications (2023)
-
Successional Changes of Microbial Communities and Host-Microbiota Interactions Contribute to Dietary Adaptation in Allodiploid Hybrid Fish
Microbial Ecology (2023)
-
Immunometabolism in biofilm infection: lessons from cancer
Molecular Medicine (2022)
-
Synergistic Effect of Sulfate-Reducing Bacteria and Cathodic Protection Potential on Hydrogen Permeation and Stress Corrosion Cracking of X100 Steel in the Maritime Mud Environment
Metallurgical and Materials Transactions A (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
