
Abstract
Dietary restriction (DR) extends animal lifespan, but imposes fitness costs. This phenomenon depends on dietary essential amino acids (EAAs) and TOR signalling, which exert systemic effects. However, the roles of specific tissues and cell-autonomous transcriptional regulators in diverse aspects of the DR phenotype are unknown. Manipulating relevant transcription factors (TFs) specifically in lifespan-limiting tissues may separate the lifespan benefits of DR from the early-life fitness costs. Here, we systematically analyse transcription across organs of Drosophila subjected to DR or low TOR and predict regulatory TFs. We predict and validate roles for the evolutionarily conserved GATA family of TFs, and identify conservation of this signal in mice. Importantly, restricting knockdown of the GATA TF srp to specific fly tissues recapitulated the benefits but not the costs of DR. Together, our data indicate that the GATA TFs mediate effects of dietary amino acids on lifespan, and that by manipulating them in specific tissues it is possible to reap the fitness benefits of EAAs, decoupled from a cost to longevity.
Similar content being viewed by others

Introduction
How can we counter ageing? Answering this question is a major goal, as ever-increasing human lifespans outpace advances in gerontology at great social, personal and financial cost.1 Dietary restriction (DR), a mild reduction in nutrient intake without malnutrition, has the evolutionarily conserved capacity to improve lifelong health, but at a cost of reduced biological fitness and vigour in youth.2 Despite having been discovered over 80 years ago,3 the molecular mechanisms underpinning DR longevity remain elusive. Elucidating these mechanisms could help isolate the benefits of DR from the corollary costs.
The lifespan benefits of DR can be recapitulated by adjusting the relative abundance of nutrients, without restricting the amount of food consumed.4,5,6 In Drosophila, the ratio of dietary sugar to yeast modulates lifespan, which is explained by essential amino acids (EAAs) from the yeast.7 Importantly, this mechanism is conserved in mice.8,9 EAAs up-regulate Target of Rapamycin (TOR) signalling,10 and recent evidence indicates that the phenotype of EAA-restricted Drosophila is recapitulated by pharmacologically suppressing TOR.11,12 Understanding of how TOR curtails lifespan is incomplete, although maintenance of proteome quality likely plays a role.13,14,15 TOR also affects transcription,16,17,18 but to date this output has been relatively poorly studied.
In Drosophila, transcriptomic responses to DR have been characterised at the cellular and organismal levels.18 Characterising tissue-specific transcription may prove key to understanding the nature of the trade-off between lifespan and fitness imposed by DR. EAAs are unusual nutrients since, by definition, they can be neither synthesised nor stored. Therefore, their dilution in the diet likely changes EAA levels uniformly across tissues. However, this altered signalling may be lifespan-limiting in only a subset of the affected tissues. In this case, longevity may be achieved by reprogramming a DR-like signalling state specifically in those tissues, whilst sparing other tissues of this manipulation would promote their optimal function. Such a manipulation would offer the benefits of an EAA-replete diet, decoupled from its pernicious lifespan-shortening effects. To date, there is evidence from Drosophila that benefits of DR are mediated at least in part by the gut19 but not the ovary,20 suggesting that costs and benefits of dietary change can indeed be partitioned amongst tissues. We have therefore focused on detailing systematically the tissue-specific transcriptional changes induced by DR. We also assess the extent to which DR-dependent transcriptional changes can be explained by TOR signalling, by suppressing its activity in an EAA-replete context, and measuring the degree to which the DR state resembles the low-TOR state. We then predict which transcription factors (TFs) mediate the observed transcriptional changes. We predict and experimentally validate a role for GATA family of TFs in DR longevity. Importantly, we confirm that costs and benefits of DR can be partitioned amongst tissues by targeted TF manipulation.
Results
This study required precise dietary manipulations. We have developed a semi-defined Drosophila diet, which is optimal for early-life egg laying, in which 50% of dietary EAAs are provided as a supplement to yeast-based medium.11 Lifespan can be extended by omission of the EAA supplement (i.e., DR), but at a cost of reduced egg laying. Thus, specific EAA dilution provides a precise model to understand how dietary variation influences lifespan, and the associated fecundity cost. Capitalising on this tool and orthology between Drosophila and vertebrate organs, we characterised EAAs’ effects on transcriptomes in the brain, fat body (the analogue of the vertebrate liver and adipose), gut, ovary, and thorax (which largely comprises muscle). This enabled us to establish systematically the transcriptional effects of EAA dilution.
Tissue-specific signatures of DR
EAAs are required ubiquitously by Drosophila tissues, leading us to expect that DR would orchestrate some global changes to gene expression across tissues. We tested for such changes at a transcriptome-wide scale. For each gene in each tissue, a measure of average fold-change in expression upon DR was calculated, and these values were clustered. Contrary to expectation, this analysis did not discriminate any clear clusters (Fig. 1). This result indicated that each tissue under study exhibited a broadly independent transcriptomic response to DR.

Tissue-specific transcriptomic signatures of DR. The heatmap shows transcriptome-wide log2 fold-changes in gene expression induced by DR in each tissue, relative to the EAA-replete control condition. Dendrograms show hierarchical clustering by Euclidian distance
To test the tissue-specificity of DR’s effect at a more granular scale, we analysed differential gene expression independently in each tissue. Differentially expressed (DE) genes were detected in all tissues except the ovary (Table 1). Since DR reduces fecundity11 and ovary size (unpublished observation), we speculate that DR down-regulates ovariole development, but without altering the transcriptome of each ovariole, with the result that no change was evident at the level of the whole ovary. Evaluating the overlap in sets of DE genes amongst tissues revealed an overwhelmingly tissue-specific effect of DR. No single gene was DE in all tissues. Only five genes (Cyp9B2, tobi, CG10910, smp-30, CG14661) were differentially expressed in more than two tissues, and pairwise intersections of DE genes comprised only a small proportion of overall transcriptional change (Supplementary Fig. 1, Supplementary data). Thus, DR had highly tissue-specific effects at the level of gene expression.
Whilst the transcripts affected by DR were highly tissue-specific, a possibility remained that these genes were involved in the same processes, which could lead to equivalent physiological effects of DR amongst tissues. To address this possibility, Gene Ontology (GO) category enrichment was analysed for each tissue’s DE genes. GO analysis revealed that the functional effects of DR were also tissue-specific (Supplementary data). Thus, DR regulates tissue-specific genes, associated with tissue-specific functions.
Overall, our transcriptome analysis indicated that the regulatory effects of DR cannot be understood at the level of the whole organism: instead, gene regulation and function of specific tissues must be studied.
The DR regulon is enriched in the TOR regulon
Although the transcriptional signatures of DR were highly tissue-specific, these changes were all due to the same upstream stimulus (i.e., EAA dilution). This commonality left open the possibility that similar cellular signalling pathways mediate DR’s effects across tissues. The TOR pathway was a strong target for such a pathway, given its evolutionarily conserved role in signalling EAA availability. In our experimental paradigm, adding rapamycin to the EAA-replete (control) medium promotes phenotypic effects similar to those of DR,11 indicating that rapamycin acts downstream of EAAs to promote DR-like signalling. Therefore, we asked whether the transcriptional effects of supplementing the EAA-replete diet with rapamycin were akin to those of DR. These samples were collected in the same experiment, facilitating their direct and quantitative comparison.
To address congruence of DR and rapamycin’s transcriptomic effects, we isolated genes that were DE following rapamycin feeding within each tissue, and examined overlaps within tissues between sets of DR-regulated and rapamycin-regulated genes. Overlaps were examined separately for up-regulated and down-regulated genes, by applying hypergeometric tests. This comparison could not be applied in the ovary, since DR did not cause differential expression there. Rapamycin feeding affected gene expression in all tissues (Supplementary Files). Furthermore, significant overlaps were detected with DR’s transcriptional targets for all tissues, although we note that no genes were up-regulated by both treatments in the brain, and in the fat body the overlap for up-regulated genes was only marginally statistically significant (Table 2). As a second, broad-scale line of testing, within-tissue correlations in changes to gene expression relative to the EAA-replete control were evaluated, both at the level of the whole transcriptome and just DE genes. This analysis revealed correlated effects of DR and rapamycin feeding on DE genes (Fig. 2). Thus, rapamycin feeding not only shared transcriptional targets with DR, but the changes in expression of those targets are quantitatively alike. This pattern was also evident transcriptome-wide, for all tissues but the fat body (Fig. 2). Altogether, these three different analyses show that, for at least one measure per tissue, the transcriptional effects of dietary EAA dilution are consistent with those of reduced TOR signalling, suggesting that TOR signals downstream of DR to regulate gene expression.

Transcriptional effects of DR are consistent with reduced TOR signalling. Panels show log2 fold-changes in expression (lFC) relative to the EAA-replete control in the DR and rapamycin-fed conditions for each tissue assessed (a - brain; b - fat body, c - gut, d - ovary and, e - thorax). Slopes of lines show correlation coefficients (Kendall’s Tau) for all genes in the transcriptome (green), or differentially expressed genes only (pink). All correlations were statistically significant (p < 0.05)
Whilst DR and rapamycin had overlapping sets of transcriptional targets, this overlap was not complete. To ask whether the functions associated with reduced TOR were consistent with those of DR, we tested the sets of rapamycin-regulated genes for GO enrichment, and compared to the equivalent analysis of DR-regulated genes. Similar GO categories were enriched in the genes regulated by DR and rapamycin (Supplementary Files): For example, in the brain, both DR and rapamycin regulated genes involved in lipid metabolism and cold acclimation, whereas in the fat body both treatments regulated genes involved in antimicrobial defence and vitellogenesis. Thus, rapamycin feeding and DR are predicted to be associated with similar physiological changes.
Predicting lifespan-regulatory transcription factors
The major goal of our study was to leverage transcriptional data to predict regulators of tissue-autonomous responses to DR. Transcription factors (TFs) were of particular interest, because of their capacity to coordinate multiple transcriptional targets to effect a given physiological programme. We searched the genes that were DE in response to DR for over-representation of motifs known to bind TFs, using a tool that has previously been used to successfully predict TFs that regulate fly phenotype.21,22,23 This analysis revealed a strong enrichment of motifs to bind GATA TFs in all tissues (Supplementary Files). The enrichment was both ubiquitous and highly statistically significant. This association was consistent with prior knowledge of the evolutionarily conserved biology of these TFs: GATA factors play known roles in signalling amino acid availability via TOR in evolutionarily diverse eukaryotes (e.g., yeast24 and mosquitos25). Furthermore, GATA TFs have known roles in signalling networks governing longevity in C. elegans.26,27,28 Thus, the motif enrichment analysis provided a logical result, associating DR-regulated transcripts to TFs with roles in longevity and signalling EAA availability downstream of TOR.
Having established an association between DR and the GATA binding element, we asked whether that signal could be attributed to TOR. We repeated the motif analysis, focusing on the transcriptional targets of rapamycin. Indeed, for all tissues, genes that were DE following rapamycin feeding were also highly enriched in the GATA element. Thus, our results suggested a circuit between circulating EAAs, cellular TOR signalling and the GATA element.
Our results indicated highly tissue-specific transcriptional effects of DR, all of which were associated with TOR/GATA signalling. How could a ubiquitous EAA-TOR-GATA TF circuit achieve tissue-specific transcriptional effects? One possibility is that the identity of the GATA TF TOR signals through depends on the tissue in question. The Drosophila genome encodes five GATA factors, and plotting their expression in our transcriptome data revealed that, indeed, each was expressed with its own pattern of tissue specificity (Supplementary Fig. 2). These unique expression profiles may combine with tissue-specific factors to translate systemic signalling into a local gene expression language. Altogether, our results outline a candidate mechanism in which GATA factors act downstream of TOR in diverse tissues to coordinate transcription.
GATA TFs are evolutionarily conserved in vertebrates including mammals, playing important tissue-specific roles in development and disease.29 DR and rapamycin promote lifespan of rodents, as well as flies, and the transcriptional effects of both interventions have been established in murine liver by microarray. We asked whether any evidence of GATA signalling could be recovered from these data. Supporting our Drosophila data, three of the six GATA TFs encoded by the mouse genome were differentially expressed, either in response to DR or a combined treatment of DR and rapamycin.30 Specifically, these microarray data suggested that, relative to a condition of adlibitum feeding, GATAd1, GATA1 and GATA6 were differentially expressed when mice were fed rapamycin and DR food, and that DR alone was sufficient to explain this effect for GATAd1. Thus, the association between DR, TOR signalling and GATA TFs appears to be conserved in a vertebrate.
Functional roles for GATA factors in the DR phenotype
Our results predicted that the tissue-specific activity of GATA factors regulated the effect of DR. We set out to test this prediction for longevity and egg laying, focussing on two GATA TFs in particular: srp and GATAe. srp was of interest because an ortholog of srp in Aedes mosquitos, AaGATAa, links EAAs to oogenesis via TOR and regulation of yolk protein precursors.25 This appears to be an evolutionarily conserved function since, in Drosophila, it has known roles in regulating oogenesis via yolk proteins in the fat body,31 although this function has not yet been connected to EAAs. Given this information and the effect of DR to reduce egg laying, srp seemed a likely mediator of responses to DR. The gut is another organ with major roles in the lifespan effect of DR,19 so GATAe was selected for testing because of its roles in the maintenance of intestinal stem cells,22 and the association between their proliferation late in life and longevity.19 We tested whether GATAe and srp in the gut and fat body, respectively, mediated the effect of DR on lifespan and egg laying.
To manipulate GATAe and srp we expressed RNAi using the well-characterised TiGS driver for the gut, and S1106 for the fat body, which are activated by feeding an inducer, RU486. With these tools, we asked whether expressing RNAi against GATAe in the gut or against Srp in the fat body altered the phenotypic effect of DR. To test these responses, we statistically modelled survival using Cox proportional hazards (CPH) analysis, using the main effects of DR and RU486, and a DR:RU486 interaction term as predictive variables. In this paradigm a significant interaction indicates that the effect of DR is contingent on eating RU486, providing a sensitive analysis without a need for pairwise comparisons between experimental conditions.
In the absence of RU486, DR extended lifespan of TiGS; UAS-GATAe[RNAi] flies (Fig. 3a, Table 3). However, knocking down GATAe altered this effect dramatically. Intestinal GATAe[RNAi] accelerated the onset of mortality independent of diet but, remarkably, reversed the effect of DR on lifespan: EAAs extended the lifespan of flies expressing intestinal GATAe[RNAi]. Whilst DR extended median lifespan by 6% in the absence of RU486, it shortened lifespan by 6% in the presence of RU486 (Table 4). CPH analysis detected a significant DR:RU486 interaction, confirming that intestinal GATAe[RNAi] significantly altered DR’s effect on lifespan. Thus, genetic intervention can reverse the sign of DR’s effect on lifespan.

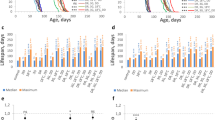
Tissue-restricted knockdown of GATAe and srp interact with dietary EAAs to determine egg laying and lifespan. Survival curves of flies fed EAA-replete or DR media, with/without expression of RNAi against GATAe in the midgut (a, TiGS, UAS-GATAe-RNAi), against srp in the fat body (b, S1106, UAS-Srp-RNAi) Complementary egg laying indices are given in panels c–d (c, TiGS, UAS-GATAe-RNAi; d, S1106, UAS-Srp-RNAi). Control medium was EAA-replete. Survival curves show proportion surviving over time. Box plots show medians, first and third quartiles, and whiskers extend 1.5 × interquartile range, with points showing individual data
Knocking down Srp also supported a role for GATA TFs in DR longevity. DR extended the lifespan of S1106/UAS-Srp[RNAi] flies in the absence of RU486 (Fig. 3b, Table 3) However, the effect of DR on lifespan was contingent on Srp knockdown, revealed by a significant interaction of RU486 and DR in CPH analysis. Fat body Srp[RNAi] extended median lifespan by 6% on EAA-replete media, but by only 2% on DR media (Table 4). Thus, expressing this RNAi seemed to extend lifespan only when flies were fed the EAA-replete medium, insulating them against the lifespan-shortening effect of the diet. Thus, Srp appears to be required in the fat body for the lifespan-shortening effect of dietary EAAs. Consistent with the study of intestinal GATAe, this result suggests a role for GATA TFs in DR longevity.
An ideal anti-ageing intervention should be effective without biological costs in early life. Reduced egg laying is a well-documented fitness cost of DR in flies. Having shown that the benefits of DR were contingent on GATA TFs, we tested whether they also mediated this cost. We counted eggs laid in early life (day 8 of adulthood) by the same flies that were assayed for survival, and analysed egg laying by fitting the same terms as were fitted for survival in an ANOVA model. Consistent with expectation, in the absence of RU486, DR reduced egg laying for both S1106/UAS-Srp[RNAi] and TiGS; UAS-GATAe[RNAi] flies. However, neither Srp[RNAi] nor GATAe[RNAi] reduced egg laying, nor interacted with the effect of DR on egg laying (Fig. 3c,d, Table 5). Thus, the roles of these GATA TFs in the gut and fat body on DR longevity appears to be independent of DR’s fecundity effects.
The effect of GATAe and Srp on lifespan but not reproduction indicated that DR’s influence on these life history traits are not obligately coupled by signalling. We hypothesised that this could be explained either by the effect of DR on egg laying and lifespan being mediated by the same mechanism in distinct tissues, and, additionally or alternatively, by distinct mechanisms in the same tissues. Srp provided a system to test these models, since our results showed that its expression in the fat body is required for the full effect of DR on longevity but not fecundity; because it is expressed in tissues other than the fat body (Supplementary Fig. 2); and because it is a known regulator of signals that promote oogenesis.31 Therefore, we ubiquitously expressed the same UAS-Srp[RNAi] construct as previously, under the control of the Daughterless-GeneSwitch (DaGS) driver. We expected that this systemic manipulation would interact with DR to determine lifespan as previously, but with an additional fecundity effect. Such an effect would indicate a role in reproduction in tissues other than the fat body. Consistent with previous results, ubiquitous Srp[RNAi] expression altered the effect of DR on lifespan, evidenced by a significant diet:RU486 interaction term in CPH survival analysis (Fig. 4a, Tables 3–4). Thus, the effect of knocking down Srp in the fat body had the same qualitative effect as ubiquitous knockdown. We then examined the effect of ubiquitous Srp[RNAi] expression on egg laying: An interaction between ubiquitous Srp[RNAi] and DR would suggest a direct role for Srp in mediating diet’s fecundity effects, whilst additive effects would indicate parallel roles. Strikingly, ubiquitous Srp[RNAi] expression reduced fecundity by an order of magnitude (Fig. 4b). This effect confirmed a role for Srp in egg laying and indicated a strong biological cost of this intervention, in contrast to the fat body-restricted knockdown. However, there was no interaction between the effect of DR and the effect of Srp[RNAi] (Table 5), consistent with Srp and dietary EAAs affecting fecundity via parallel mechanisms. Thus, the effect on longevity of knocking down Srp appears to be spatially discrete from its fecundity effect, if limited to the fat body. Systemically, its fecundity effect appears to be via a diet-independent mechanism. Thus, whilst all three of the GATA knockdowns in this study interacted with the effect of diet to determine lifespan, none of them interacted with diet to determine fecundity.

Systemic srp knockdown recapitulates benefits of DR at enhanced cost. Survival curves of flies fed EAA-replete or DR media, with/without expression of RNAi against srp under control of daughterless GeneSwitch (a), with complementary egg laying indices (b). Control medium was EAA-replete. Survival curves show proportion surviving over time. Box plots show medians, first and third quartiles, and whiskers extend 1.5 × interquartile range, with points showing individual data
Discussion
DR improves lifelong health in a range of organisms, from yeast to primates. A growing body of evidence shows the particular importance of dietary nutrient balance for ageing, particularly lowered protein:carbohydrate ratio,32 which implicates amino-acid sensitive TOR signalling. A likely role of TOR was recently underlined in Drosophila, by the demonstration that active TOR is required for EAA enrichment to shorten lifespan.11 In the present study we have confirmed that the transcriptional effects of DR are consistent with reduced TOR signalling. Moreover, our results now identify a third level of regulation, implicating the GATA family of TFs as mediators of dietary EAAs’ lifespan effects. The evolutionarily conserved roles of this family of TFs in signalling EAA availability, TOR signalling, oogenesis and tissue-specific gene regulation, in addition to their regulation by DR and rapamycin in murine liver, make them potential mechanisms of longevity assurance worthy of further investigation. Roles of GATA TFs in worm longevity have been established.26,27,28 Mice are also long-lived when fed a restricted diet or rapamycin, and the finding that GATA TFs are differentially regulated in mouse liver,30 reveals evolutionary conservation and warrants study of their role in mouse longevity.
The major motivation to understand how DR affects physiology is to identify means to recapitulate its benefits in later life without associated biological costs, not to mention costs of an ascetic lifestyle. Our results show that GATA TFs interact with diet to determine lifespan, but not egg laying–the major fitness cost of DR in Drosophila. Furthermore, whilst ubiquitous expression of Srp[RNAi] did affect egg laying, there was no interaction with diet: in fact, none of the GATA TF knockdowns altered the effect of diet on egg laying. Furthermore, restricting the Srp[RNAi] to the fat body made the flies long-lived without any effect on fecundity. Thus, GATA TFs are candidates to relax tradeoffs between life-history traits relevant to ageing, particularly when targeted specifically to lifespan-relevant tissues, allowing the benefits of feeding to repletion whilst sparing costs to lifespan.
An important outcome of our analysis is the highly independent transcriptional response of each tissue to DR. We hypothesised that transcriptional programmes under tissue-specific control would be sufficient for longevity benefits of DR, by disconnecting lifespan-limiting pathologies and processes from the homoeostatic regulation of other tissues. Importantly, the experiments involving srp knock-down demonstrate that benefits of DR can be reaped by targeting transcription factors in specific organs, making flies constitutively long-lived even when eating an EAA-replete diet. Whilst these results could possibly result from stronger expression of the DaGS driver than the S1106 driver, this seems an unlikely explanation given that both constructs imparted a similar lifespan phenotype, but vastly different egg laying phenotypes. Curiously, GATAe knockdown in the gut changed the sign of the effect of DR on lifespan, suggesting either a different optimal dietary balance in these flies, or that elevated EAAs rescued the pathological effects of gut-specific GATAe knockdown. These experiments implicate GATA factors as mechanistic links between diet, tissue-specific gene regulation and lifespan.
The putative connections between lifespan, diet and GATA TFs are entirely consistent with the evolutionarily conserved functions of these proteins. The GATA TFs are an ancient family, with well-characterised roles in development and nutrient signalling. In multicellular differentiated organisms, GATA TFs are required in the development of multiple tissue types, which in Drosophila includes the heart,33 fat body34 and gut.35 However, their functions in adulthood are poorly characterised. GATAe plays a role in midgut homoeostasis in adult Drosophila, but it is unclear where it integrates into broader midgut signalling networks.36,37 One of the better-described roles for GATA factors in adult animals is in nutrient regulation of oogenesis in mosquitos.25,38,39,40 In Aedes aegypti, oogenesis requires a blood meal, which contains the mosquito’s only source of protein. Before feeding, egg production is suppressed partly by a GATA TF repressing expression of the major yolk precursor protein, Vg, in the fat body. After feeding, TOR enhances expression of the transcriptional activator AaGATAa, which de-represses Vg expression. This circuit appears to be at least partially conserved, since srp regulates yolk protein expression in Drosophila,31 and our results show that its ubiquitous knockdown strongly attenuates egg laying, although the absence of an interaction with diet indicates a mechanism independent of EAAs. Evidence from yeast suggests that the role of GATA factors in regulating nitrogen metabolism is basal in Eukaryotes. In Saccharomyces cerevisiae, selective amino acid catabolism is controlled by a circuit known as Nitrogen Catabolite Repression: when nitrogen availability supports only poor growth, TOR-dependent nuclear localisation of a GATA TF activates expression of genes involved in the transport and metabolism of less-preferred nitrogen sources.24 Together, these data highlight conserved connections between protein nutrition, growth, reproduction, TOR and transcriptional regulation by GATA TFs; and we have now connected GATA TFs to longevity by DR. These roles are entirely consistent with evolutionary theories that ageing is a consequence of deleterious pleiotropy with mechanisms under selection in early life.41
This study reveals tissue-specific patterns of transcriptional regulation in response to the longevity-promoting restriction of EAAs. The transcriptional effects of EAAs are associated with TOR signalling and motifs to bind GATA TFs, and the tissue-specific activity of GATA factors appears to dictate the effect of diet on phenotype. Importantly, these experiments also suggest that the costs and benefits of dietary variation may be mediated by different tissues, and therefore that benefits may be reaped without fitness tradeoffs by tissue-specific genetic interventions. The evolutionary conservation of GATA factors, of their connection to regulating amino acid metabolism, and of the capacity of DR to mediate lifespan extension, suggests that GATA factors may be relevant to DR’s anti-ageing effect in a broad range of animals.
Materials and methods
Diets
Diets were prepared according to ref. 11 The base (1SY) medium contained 100 g/l autolysed yeast (MP Biomedicals, OH, USA), 50 g/l sucrose (Tate & Lyle, London, UK), 15 g/l agar (Sigma-Aldrich, Dorset, UK), 30 ml/l nipagin (Chemlink Specialities, Dorset, UK), and 3 ml/l propionic acid (Sigma-Aldrich, Dorset, UK). The EAA-replete control medium comprised DR food with an EAA supplement dissolved in pH 4.5 water (final concentrations in fly media: L-arginine 0.43 g/l, L-histidine 0.21 g/l, L-isoleucine 0.34 g/l, L-leucine 0.48 g/l, L-lysine 0.52 g/l, L-methionine 0.1 g/l, L-phenylalanine 0.26 g/l, L-threonine 0.37 g/l, L-tryptophan 0.09 g/l, L-valine 0.4 g/l: all suppled by Sigma). EAA + rapamycin food consisted of EAA-replete medium with a rapamycin supplement (LC laboratories, MA, USA) dissolved in ethanol, to a final concentration of 200 µM in the diet. For RNAi experiments, RU486 (Sigma M8046) dissolved in ethanol was added to 1SY or EAA food to a final concentration of 200 µM.
Fly culture
Outbred wild-type Dahomey flies bearing the endosymbiont Wolbachia were cultured on a 12:12 light cycle at 25 °C and 60% humidity, on 1SY medium. For RNAi experiments, the TiGS, S1106 and DaGS drivers, and RNAi constructs (UAS-srp[RNAi]: Vienna Stock Center #35578; UAS-GATAeTRiP: Bloomington Stock Center #33748) were backcrossed into Dahomey flies bearing the w1118 mutation for at least six generations. UAS-GATAeTRiP was backcrossed into this background by genotyping individual flies by PCR of the vector. S1106 drives in the fat body and anterior midgut, however the effect of gut srp[RNAi] can be excluded owing to the absence of srp expression in the gut (Supplementary Files). All flies were maintained at large population sizes to maintain outbred genetic diversity. For all experiments, eggs were collected following an 18 h lay on grape juice agar, added to bottles of 1SY at a standardised density and cultured to adulthood. Newly emerged experimental females were mated on fresh food for 48 h, lightly CO2 anaesthetised, then allocated to experimental diets without males (10 females/vial). For egg laying and lifespan experiments, survival was scored and flies transferred to new media at least three times per week. Egg laying was scored on day 8 after eclosion, after 18 h egg laying. Phenotyping experiments were set up at different times, one per experimental genotype, thus multiple experiments validate the role of GATA TFs.
RNA sequencing
Tissues were dissected in ice-cold RNAlater solution, 6–10 h into the flies’ light cycle, and frozen at −80 °C. RNA was extracted using the QIAGEN total RNA isolation kit and quantified on an Agilent 2100 bioanalyser. Sequencing was performed by the high throughput genomics services center at the Huntsman Cancer Institute (University of Utah). Sample concentration and purity of RNA was measured on a NanoDrop spectrophotometer, and RNA integrity was assessed on an Agilent 2200 TapeStation. Illumina TruSeq libraries were prepared from this RNA with the Illumina TruSeq Stranded mRNA Sample Prep kit and sequenced on an Illumina HiSeq2000 101 v3 platform using paired-end sequencing.
Data analysis
Reads were aligned to D. melanogaster genome v6.19 using HISAT2 v2.1.0 and enumerated using featureCounts v1.6.0. Unmapped reads were discarded. Enumerated reads were analysed in R and bioconductor (3.3.1). Two gut samples from the DR condition were found to be internally inconsistent with other gut samples, and also with gut transcriptomes from external experiments. These samples were therefore excluded from further analysis. Fold-changes in expression were calculated by DESeq2 (1.18.1). Differential expression and fold-changes in expression were determined for each tissue by DESeq2, by fitting a model of the form
where y represented read counts of transcripti, replicate coded for biological replicate with a three-level factor, and diet coded for whether the flies were fed control medium, DR medium, or rapamycin-supplemented medium. Contrasts were then applied between the control versus DR condition, and control versus rapamycin-fed condition. P-values were corrected using independent hypothesis weighting, with base mean of normalised counts as a covariate. Differential expression was considered statistially significant when corrected P-values were ≤ 0.01. Motif enrichment amongst differentially expressed genes was analysed using i-Cis target.42 Heatmaps were clustered by Euclidian distance metrics and plotted using the heatmap.2 function from the gplots library, either on fold-changes calculated by DESeq2, or median of variance-stabilised GATA TF expression. Intersections between DE gene sets were visualised using the upset library.43
Fly survival and egg laying data were analysed in R (v3.1.1). Survival data were analysed using the coxph function from the survival library. Egg laying data were normalised to number of flies per vial and log-transformed, and analysed by fitting a linear model (ANOVA) using the lm function. Both survival and egg laying data were analysed with a model of the form
where y coded for survival or egg laying index, RU and EAAs for the presence of RU486 and EAAs, respectively, and the interaction of the two.
Code availability
All code is available on request.
Data availability statement
Raw RNAseq data have been deposited with ArrayExpress, Accession E-MTAB-6584. All processed RNAseq data used in Figs. 1 and 2 and phenotypic data used in Figs. 3 and 4 are available in Supplementary Files.
References
Niccoli, T. & Partridge, L. Ageing as a risk factor for disease. Curr. Biol. 22, R741–R752 (2012).
Piper, M., Partridge, L., Raubenheimer, D. & Simpson, S. J. Dietary restriction and aging: a unifying perspective. Cell. Metab. 14, 154–160 (2011).
McCay, C. M., Crowell, M. & Maynard, L. The effect of retarded growth upon the length of life span and upon the ultimate body size. J. Nutr. 10, 63–79 (1935).
Mair, W., Piper, M. D. & Partridge, L. Calories do not explain extension of life span by dietary restriction in Drosophila. PLoS Biol. 3, e223 (2005).
Lee, K. et al. Lifespan and reproduction in Drosophila: New insights from nutritional geometry. Proc. Natl Acad. Sci. USA 105, 2498–2503 (2008).
Skorupa, D. A., Dervisefendic, A., Zwiener, J. & Pletcher, S. D. Dietary composition specifies consumption, obesity, and lifespan in Drosophila melanogaster. Aging Cell. 7, 478–490 (2008).
Grandison, R. C., Piper, M. D. & Partridge, L. Amino-acid imbalance explains extension of lifespan by dietary restriction in Drosophila. Nature 462, 1061–1064 (2009).
Solon-Biet, S. et al. The ratio of macronutrients, not caloric intake, dictates cardiometabolic health, aging, and longevity in Ad Libitum-fed mice. Cell. Metab. 19, 418–430 (2015).
Solon-Biet, S. et al. Macronutrient balance, reproductive function, and lifespan in aging mice. Proc. Natl Acad. Sci. USA 112, 3481–3486 (2015).
Bar-Peled, L. & Sabatini, D. Regulation of mTORC1 by amino acids. Trends Cell. Biol. 24, 400–406 (2014).
Emran, S., Yang, M., He, X., Zandveld, J. & Piper, M. D. Target of rapamycin signalling mediates the lifespan-extending effects of dietary restriction by essential amino acid alteration. Aging 6, 390–398 (2014).
Simpson, S. J., Couteur, D. G. & Raubenheimer, D. Putting the balance back in diet. Cell 161, 18–23 (2015).
Taylor, R. C. & Dillin, A. XBP-1 is a cell-nonautonoous regulator of stress resistance and longevity. Cell 153, 1435–1447 (2013).
Katewa, S. & Kapahi, P. Role of TOR signaling in aging and related biological processes in Drosophila melanogaster. Exp. Gerontol. 46, 382–390 (2011).
Vilchez, D., Simic, M. S. & Dillin, A. Proteostasis and aging of stem cells. Trends Cell. Biol. 24, 161–171 (2014).
Bülow, M. H., Aebersold, R., Pankratz, M. J. & Jünger, M. A. The Drosophila FoxA ortholog fork head regulates growth and gene expression downstream of target of rapamycin. PLoS One 5, e15171 (2010).
Robida-Stubbs, S. et al. TOR signaling and rapamycin influence longevity by regulating SKN-1/Nrf and DAF-16/FoxO. Cell. Metab. 15, 713–724 (2011).
Tiebe, M. et al. REPTOR and REPTOR-BP regulate organismal metabolism and transcription downstream of TORC1. Dev. Cell. 33, 272–284 (2015).
Regan, J. C. et al. Sex difference in pathology of the ageing gut mediates the greater response of female lifespan to dietary restriction. eLife 5, e10956 (2016).
Mair, W., Sgrò, C. M., Johnson, A. P., Chapman, T. & Partridge, L. Lifespan extension by dietary restriction in female Drosophila melanogaster is not caused by a reduction in vitellogenesis or ovarian activity. Exp. Gerontol. 39, 1011–1019 (2004).
Potier, D. et al. Mapping Gene Regulatory Networks in Drosophila Eye Development by Large-Scale Transcriptome Perturbations and Motif Inference. Cell Rep. 9, 2290–2303 (2014).
Dutta, D. et al. Regional cell-specific transcriptome mapping reveals regulatory complexity in the adult Drosophila midgut. Cell Rep. 12, 346–358 (2015).
Dobson, A. J. et al. Nutritional programming of lifespan by FOXO inhibition on sugar-rich diets. Cell Rep. 18, 299–306 (2017).
Cooper, T. G. Transmitting the signal of excess nitrogen in Saccharomyces cerevisiae from the Tor proteins to the GATA factors: connecting the dots. FEMS Microbiol. Rev. 26, 223–238 (2002).
Attardo, G. M., Higgs, S., Klingler, K. A., Vanlandingham, D. L. & Raikhel, A. S. RNA interference-mediated knockdown of a GATA factor reveals a link to anautogeny in the mosquito Aedes aegypti. Proc. Natl Acad. Sci. USA 100, 13374–13379 (2003).
Zhang, P., Judy, M., Lee, S.-J. & Kenyon, C. Direct and indirect gene regulation by a life-extending FOXO protein in C. elegans: roles for GATA factors and lipid gene regulators. Cell. Metab. 17, 85–100 (2013).
Murphy, C. T. et al. Genes that act downstream of DAF-16 to influence the lifespan of Caenorhabditis elegans. Nature 424, 277–284 (2003).
Budovskaya, Y. V. et al. An elt-3/elt-5/elt-6 GATA transcription circuit guides aging in C. elegans. Cell 134, 1–13 (2008).
Lentjes, M. et al. The emerging role of GATA transcription factors in development and disease. Expert Rev. Mol. Med. 18, e3 (2016).
Fok, W. C. et al. Combined treatment of rapamycin and dietary restriction has a larger effect on the transcriptome and metabolome of liver. Aging Cell. 13, 311–319 (2014).
Lossky, M. & Wensink, P. Regulation of Drosophila yolk protein genes by an ovary-specific GATA factor. Mol. Cell. Biol. 15, 6943–6952 (1995).
Couteur, D. G. et al. The impact of low-protein high-carbohydrate diets on aging and lifespan. Cell. Mol. Life. Sci. 73, 1237–1252 (2016).
Sorrentino, R. P., Gajewski, K. M. & Schulz, R. A. GATA factors in Drosophila heart and blood cell development. Semin. Cell. Dev. Biol. 16, 107–116 (2005).
Sam, S., Leise, W. & Hoshizaki, D. The serpent gene is necessary for progression through the early stages of fat-body development. Mech. Dev. 60, 197–205 (1996).
Murakami, R., Okumura, T. & Uchiyama, H. GATA factors as key regulatory molecules in the development of Drosophila endoderm. Dev. Growth Differ. 47, 581–589 (2005).
Buchon, N. et al. Morphological and molecular characterization of adult midgut compartmentalization in Drosophila. Cell Rep. 3, 1725–1728 (2013).
Okumura, T., Takeda, K., Kuchiki, M. & Akaishi, M. GATAe regulates intestinal stem cell maintenance and differentiation in Drosophila adult midgut. Dev. Biol. 410, 24–25 (2015).
Hansen, I. A., Attardo, G. M., Park, J.-H., Peng, Q. & Raikhel, A. S. Target of rapamycin-mediated amino acid signaling in mosquito anautogeny. Proc. Natl Acad. Sci. USA 101, 10626–10631 (2004).
Hansen, I. A., Attardo, G. M., Roy, S. G. & Raikhel, A. S. Target of rapamycin-dependent activation of S6 kinase is a central step in the transduction of nutritional signals during egg development in a mosquito. J. Biol. Chem. 280, 20565–20572 (2005).
Park, J.-H., Attardo, G. M., Hansen, I. A. & Raikhel, A. S. GATA factor translation is the final downstream step in the amino acid/target-of-rapamycin-mediated vitellogenin gene expression in the anautogenous mosquito Aedes aegypti. J. Biol. Chem. 281, 11167–11176 (2006).
Williams, G. C. Pleiotropy, natural selection, and the evolution of senescence. Evolution 11, 398–411 (1957).
Herrmann, C., de Sande, B., Potier, D. & Aerts, S. i-cisTarget: an integrative genomics method for the prediction of regulatory features and cis-regulatory modules. Nucleic Acids Res. 40, e114–e114 (2012).
Lex, A., Gehlenborg, N. & Strobelt, H. UpSet: visualization of intersecting sets. IEEE Trans. Vis. Comp. Graph. 20, 1983–1992 (2014).
Acknowledgements
We thank VM Howick, C Slack, N Alic and AE Douglas for comments on drafts of the manuscript; and C Stubben of the Huntsman Cancer Institute for assistance with data management. This study was funded by grants to MP from the Biotechnology and Biological Sciences Research Council (BBI011544/1), the Royal Society (UF100158 and RG110303) and the Australian Research Council (FT150100237).
Author information
Authors and Affiliations
Contributions
Devised the study and acquired funding: M.D.W.P. Designed the study: A.J.D., M.Y. and M.D.W.P. Performed experiments: A.J.D., X.H., M.Y., Y.F. and E. Bolukbasi. Analysed data: A.J.D. and E. Blanc. Wrote the manuscript: A.J.D., M.D.W.P.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Dobson, A.J., He, X., Blanc, E. et al. Tissue-specific transcriptome profiling of Drosophila reveals roles for GATA transcription factors in longevity by dietary restriction. npj Aging Mech Dis 4, 5 (2018). https://doi.org/10.1038/s41514-018-0024-4
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41514-018-0024-4
