
Abstract
Acoustic communication, broadly distributed along the vertebrate phylogeny, plays a fundamental role in parental care, mate attraction and various other behaviours. Despite its importance, comparatively less is known about the evolutionary roots of acoustic communication. Phylogenetic comparative analyses can provide insights into the deep time evolutionary origin of acoustic communication, but they are often plagued by missing data from key species. Here we present evidence for 53 species of four major clades (turtles, tuatara, caecilian and lungfish) in the form of vocal recordings and contextual behavioural information accompanying sound production. This and a broad literature-based dataset evidence acoustic abilities in several groups previously considered non-vocal. Critically, phylogenetic analyses encompassing 1800 species of choanate vertebrates reconstructs acoustic communication as a homologous trait, and suggests that it is at least as old as the last common ancestor of all choanate vertebrates, that lived approx. 407 million years before present.
Similar content being viewed by others

Introduction
Despite the unquestionable importance of acoustic communication among vertebrates, our knowledge regarding its origin remains sparse. The current consensus based on available evidence favours a convergent origin of acoustic communication among vertebrates: studies on acoustic sensory abilities show that the morphology in the hearing apparatus and its sensitivity vary considerably among vertebrates1,2,3. This, in addition to observed differences in vocal tract morphology, suggests that acoustic communication likely evolved multiple times, emerging independently among diverse clades3. Phylogenetic analyses used to reconstruct the ancestral state of acoustic communication along the tree nodes, whilst suggestive of multiple origins4, are arguably complicated by missing data from key taxa.
An alternative hypothesis is that acoustic communication has a common and ancient evolutionary origin. In support of this, vertebrate hearing epithelia and cerebral promotor circuits that control vocal behaviours are considered to be homologous and operate in the same hindbrain compartment, respectively5,6,7,8,9. Furthermore, in spite of the variety of sound production mechanisms, all Choanata (Dipnoi (lungfishes) + Tetrapoda) lineages have lungs as the physical source of their calling behaviours.
Among vertebrates, clades that can be easily recognised to produce complex sounds (i.e. frogs, crocodilians, birds and mammals) have been studied extensively (e.g. ref. 10,11,12). However, some vertebrate clades, in contrast, have been assumed to be non-vocal based on limited or sparse data. As a consequence, the absence of concrete evidence for vocal production is sometimes treated as evidence of non-vocal tendencies (e.g. ref. 4). Central to a robust reconstruction of acoustic communication is a systematic documentation of these key, neglected groups.
Here, we investigate the evolutionary origins of acoustic communication in choanate vertebrates combining critical data with phylogenetic trait reconstruction methods using a comprehensive dataset. We assess the acoustic communication abilities in species of diverse vertebrate groups, including Lepidosauria (tuataras, lizards and snakes), non-anuran Amphibia (salamanders and caecilians), Chelonians (turtles) and lungfishes (Dipnoi) that are key to mapping vocal communication in the vertebrate tree of life. Using this dataset combined with data of well-known acoustic clades (e.g. mammals, birds and frogs), we test if the evolutionary origin of acoustic communication is shared among choanate vertebrates. We suggest a single origin of acoustic communication in the last common ancestor of all Choanata over 400 million years before present (mybp).
Results
Origins of acoustic communication
We found widespread evidence for acoustic behaviour among all choanate vertebrates. Our recordings include 53 species that belong to groups often thought to be non-vocal and commonly neglected in vocal communication research (Supplementary data 1). Of these, 50 species are turtles—representing over 54% of all genera and more than 14% of all extant species13. We also recorded tuataras (Sphenodon punctatus), one species of caecilian (Typhlonectes compressicauda), and the South American lungfish (Lepidosiren paradoxa). All recorded species were found to possess a varied acoustic repertoire comprising a number of different sounds (see Figs. 1, 2 and supplementary Data 2 to listen to sounds, and supplementary Data 3 for sound descriptions).

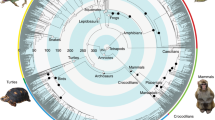
Tree includes 1800 choanatian species assigned either with the character presence or absence of acoustic communication. Pie charts at ancestral nodes indicate likelihood of each character state. Colours in the spectrograms represent sound intensity, with warm colours representing high intensity and cold colours (i.e. blue) representing low intensity or absence of sounds. Character states for each species can be accessed in Supplementary Data 4.

Tree includes every turtle genus assigned either with the character presence or absence of acoustic communication. Pie charts at ancestral nodes indicate the likelihood of each character state. Colours in the spectrograms represent sound intensity, with warm colours representing high intensity and cold colours (i.e. blue) representing low intensity or absence of sounds. Character states for each species can be accessed in Supplementary data 5.
A critical review of the extensive literature focusing on groups often considered to lack acoustic communication resulted in a total of 106 species, including 54 turtles, 14 lepidosaurs, 29 salamanders, four caecilians, four frogs and one lungfish having been reported to engage in vocal communication (Supplementary Data 1).
The African lungfish (Protopterus annectens) has been reported to produce sounds14 and to being able to perceive sounds both in the water and air15. Among the ten families of caecilians, we found evidence of acoustic communication in representatives of four of them (Dermophiidae, Grandisoniidae, Ichthyophiidae and Siphonopidae)16,17,18,19. In salamanders, eight out of 10 families have representatives known to produce vocalisations, with evidence being absent only in Hynobiidae and Rhyacotritonidae18,19,20,21,22. We also found evidence for acoustic communication in all species of frogs of the genera Ascaphus23,24 and Leiopelma24,25,26. Among Lepidosauria, examples of acoustic communication are found in most groups of Gekkota27,28, and in tuataras (ref. 29; present study). Among turtles, we found evidence of acoustic communication in representatives from all families, with some species producing over 15 different types of calls used in various situations, including parental care30,31. These findings confirm that the ability to produce vocalisations is distributed across such groups.
Ancestral-state reconstructions
The ancestral-state reconstruction for Choanata recovered the presence of acoustic communication as an unambiguous homologous trait, being present in the common ancestor of all choanate vertebrates (407 mybp), and in the majority of the tree nodes (e.g. tetrapods, amniotes, reptiles; Fig. 1). An ancestral-state reconstruction using a tree containing only turtle genera resulted in the presence of acoustic communication in every ancestor node, except for the Geoemydidae family (Fig. 2). This is likely an artefact of the effect of missing data given the limited representation of species of this clade in our dataset, many of which are endangered and hard to access. Nevertheless, the presence of this trait is unambiguously ancestral among turtles.
Discussion
Data across the turtle tree of life together with other critical taxa in combination with available evidence across all major tetrapod clades totalling in excess of 1800 species confirms a common origin of sound production and acoustic communication among choanate vertebrates, dating from the Palaeozoic (at least 407mybp). These findings support the hypothesis that innovations in the sound production apparatus among choanates were acquired after the first, common appearance of acoustic communication within this group.
The interpretation of acoustic behaviour as a non-homologous trait proposed in previous research3,4 was driven largely by a lack of information on key groups of animals. That is, analyses of ancestral-state reconstruction are complicated by missing data which can subsequently be treated as evidence of absence. Nevertheless, the recent growth of evidence for acoustic communication among certain tetrapod groups, commonly considered to be non-vocal, such as aquatic turtles (e.g. ref. 31,32, and the data provided by us in this paper), are key in revealing the common ancestry of such behaviour. In fact, including evidence from only 14 species (12 turtles, tuatara and lungfish) to the analysis proposed by Chen & Wiens4 was enough to recover opposite results, that were reinforced by the inclusion of data from our critical study of the literature. The sensitivity of ancestral state reconstruction analyses to the character state of key lineages makes a deeper investigation of poorly studied groups imperative.
Knowledge of the natural history of organisms is fundamental in surveys of the macroevolution of certain features. The intensive documentation of vocal communication in turtles in our study is an example. Recordings, observations and subsequent analyses in a phylogenetic framework suggest the homology of vocal communication across turtles and in the last common ancestor of the clade. This result strengthens our broader conclusions on the origins of acoustic communication among choanate vertebrates. Specifically, by evidencing that acoustic communication is widespread and homologous among all turtle genera, we ensure that analysis with a much less comprehensive sample of this group is trustworthy and not subject to extensive interpretation changes by switching the state character of only a few species.
With the inclusion of several taxa of comparatively understudied groups to the analyses, we show that the use of sounds in communication is not only a homologous but also conserved behaviour, widely distributed among choanate vertebrates. The wide variety of mechanisms of sound production—with some of the most distinguished examples being the bird syrinx33,34, the trombone-like crest of Parasaurolophus dinosaurs35,36, and the sound apparatus of bats and dolphins, capable of producing ultrasounds37—also deserves consideration, in order to reveal the anatomical and physical transformations that must have been involved around a common neurobiological framework of sound production.
The larynx has an important role in tetrapod acoustic production, being the main site of vocal production in most lineages38. Although some of the acoustic structures used by choanate vertebrates do not share their embryological origins with the larynx—for example, the bird syrinx develops from different tissues33,34,39—they all share the use of air circulation in the production of sounds, powered by the lungs (with the exception of Plethodontid salamanders that lost the latter). Additionally, all choanate vertebrates are able to produce laryngeal sounds, including birds, while hissing40.
Furthermore, all vertebrates share the location of motoneurons associated with vocalisations in the caudal hindbrain5,41. Vocal-acoustic and pectoral-gestural signalling also share evolutionary and developmental neural origins5,9,42, which implies a common vocal-sonic central pattern generator in the vertebrate brain5,9,41,43.
Many salamanders, caecilians, lizards and snakes produce complex, modulated sounds44,45, that were not considered acoustic communication in the previous analysis due to its usage being usually applied to inter-specific communication4—e.g. defensive behaviour44,46—or allegedly produced by accident47. However, it cannot be excluded that these sounds could have had a common evolutionary origin to those used for intraspecific communication. The same rationale applies to simpler sounds. Hissing and sniffing sounds produced by most vertebrates, especially amniotes48—and nearly all reptiles28—might also contain more information than what we account for. e.g. non-vocal sounds encode individual signals in birds40 and some colubrid snakes mimic the hiss of vipers49.
Considering the fact that communication is usually multimodal, and the loss of acoustic abilities can quickly happen due to redundancy with other channels such as visual and chemical ones in many taxa50, analyses based on ancestral state reconstruction can be biased due to character interpretation and loss of track caused by recent changes in character state in the tip species. The widespread usage of hissing and sniffing sounds among vertebrates might be further evidence that acoustic communication is a shared character in this group and started before the diversification of sounds and their various usages by different clades. Because we chose more conservative methods that include only sounds that play a role in communication with conspecifics, and excludes inter-specific communication (e.g. defence hisses by snakes and other species), future studies might broaden this scope by including such calls, since hisses, in particular, appear to be common across tetrapods, and are typically defence vocalisations.
If all sounds have the same evolutionary origin, vocalisations produced by clades commonly conceived to have had a secondary development of acoustic communication in previous works, such as non-anuran amphibians and Gekkota, mainly Gekkonidae and Pygopodidae4,28, would be homologous to the ones produced by other vertebrates.
The recovery of a single origin of acoustic communication among vertebrates in our analysis reinforces the need to investigate hissing and sniffing sounds, here not classified as acoustic communication. Investigations on the potential primary homology of these sounds, taking a neurobiological, physiological, and anatomical approach, would be important to shed light on this discussion. Comparative studies on the diversification of calls and vocal apparatus, including different vertebrate groups, are also needed to clarify the question of homology.
Vocalisations are also a widespread behaviour among actinopterygian fishes. In this case, however, it may have evolved recurrently and independently over 30 times during their evolutionary history5,9,51,52,53. Although many are the mechanism of sound production among actinopterygians, swim bladder vibrations seem to be the most widespread and ancient of them53. Considering that the homology of lungs among vertebrates is still debated, with strong evidence for a common origin between the lung and the swim bladder54,55,56, a shared origin of acoustic behaviour between choanates and actinopterygians cannot be ruled-out. In this case, expanding our analysis to include actinopterygians may reveal the origins of acoustic communication to be even older than 407mybp57.
Whilst inferring a common origin of acoustic communication among actinopterygians and choanates may be complicated at present due to lack of data, additional alternative evidence points in the same direction—perhaps a deep homology58 of shared brain mechanisms5,9,41,42,43. Furthermore, the gill arches used by fish as a breathing system evolved into the tetrapod hyoid and laryngeal apparatus, used in many mechanisms that include breathing, feeding and sound production59. Such connections might be modulated by the same or similar brain channels, that might suggest evolutionary continuity9.
A challenge to test the hypothesis of a common origin of acoustic communication among some actinopterygian lineages and choanates will be understanding how the morphological transformations involved in the transition to choanates affected the mechanisms of sound production. Palaeontological data and phylogenetic reconstructions are the most common approaches applied to shed light on the evolutionary transformation of traits. However, to date, palaeontological data that are key to a robust resolution of the origins of acoustic communication among actinopterygians and sarcopterygians are missing48,60. This raises a number of additional questions, but of particular importance: is the acoustic communication of choanates an innovation homologous to the acoustic communication based on the swim bladder observed in fishes? If so, did acoustic communication first appear among actinopterygians or in the event that precedes them—such as in the transition from ‘protochordates’ (~550 mybp, 5)? Integrative comparative studies of embryological development, physiology of vocal apparatus and of vocal neural architecture across brain regions combined with gene expression among taxa will be helpful to trace the evolution of acoustic behaviour among vertebrates. The common ancestral origin of acoustic communication provides further justification for the use of choanate animals as models in the study of the origins of human language and speech.
Methods
No general ethics approval was required for this study as it was conducted with animals that were already captive. Nevertheless, approval was granted by specific institutions that required analysis from a committee (includes Chester Zoo, CEQUA and Turtle Island).
Sound recordings and analysis
For underwater sound recordings, we used the OceanBase (developed by the Laboratory of Acoustic and Environment—University of Sao Paulo, in partnership with Bunin tech®), an acoustic recorder specifically designed for underwater noise monitoring. It has a sensitivity of −157 ± 2 dB rel 1 V/uPa ± 2 dB and a frequency band of 5 Hz–90 kHz. In-air recordings were conducted using a Tascam® recorder DR-100MKIII with a sensitivity of −115.5 ± 0.5 dB rel 13 mV/uPa ± 4 dB and a frequency band of 5 Hz–96 kHz.
Recordings were made in captivity using plastic pools to ensure that all sounds were produced by the animals being recorded. Each species was recorded for at least 24 h, capturing both day and night activity. We aimed for recording males and females in different life stages whenever specimens were available. We also recorded ambient sound without the presence of any animals in order to account for possible noise/interference.
Sounds recorded were analyzed using Raven Pro 1.661 and Praat62. Sounds were measured using six acoustic parameters: Number of bouts, fundamental frequency (Hz), minimum frequency (Hz), peak frequency (Hz), duration (s), and sound type (simple or complex). These parameters were only used as a first description of the repertoires, and were not subjected to any analysis in the present study.
Acoustic communication data
We recorded 53 species, that include 50 turtle species, one caecilian, one lungfish and tuataras, of which all communicated acoustically (Figs. 1, 2, Supplementary Data 1–3 and Supplementary Note 1). Apart from the vocalisations we recorded, most of the acoustic communication data used in this work originates from the dataset published by Chen & Wiens4, that includes 1799 tetrapod species (supplementary Data 4). In addition, we searched for information on acoustic communication among groups that are often considered to be silent (i.e. Testudines, Lepidosauria, Gymnophiona, Caudata and some anuran species). We gathered information from peer-reviewed articles, books and personal communication with researches that work directly with the referred groups (Supplementary Data 1). Our search was conducted using Google Scholar and Web of Science between March and November 2021 using the keywords “acoustic communication”, “call”, “vocal communication”, “vocalisation”, “song”, and “sound”, in association with the species’ name and superior taxonomic ranks. The search was conducted following PRISMA63 guidelines (Supplementary file 1).
Sound communication entails not only that the animal is producing a sound, but also that it has communicative significance. To avoid mistakes in determining its significance, and to ensure we are comparing homologous types of acoustic communication, we favoured the hypothesis that the presence of a complex repertoire (presence of a number of different sounds and/or harmonic calls) entails communicative meaning and considered only sounds produced by the respiratory tract (excluding scale scratching and tail rattling, for example). We also decided to exclude lungless salamanders of the Plethodontidae family, as they might have a different, non-homologous method of sound production20.
Additionally, in order to ensure character homology, our analysis includes only Sarcopterygian lineages (namely Choanata: Tetrapoda + Dipnoi; ref. 64), as we hypothesised the presence of lungs as a major driver for acoustic behaviour in this clade. Sound production systems in other vertebrate clades are diverse and we have not yet enough evidence to infer its homology.
Sounds produced during defensive behaviour such as hissing and sniffing in lizards or bellowing in snakes were not considered to be intraspecific acoustic communication and, therefore, were not included. Although these behaviours might have a common origin to the sounds here considered acoustic communication, we lack evidence to support this claim and opted for a more conservative approach. Only sounds that were considered by literature reports to be used by the studied species in intraspecific communication were considered.
Compiled information regarding amphibians and reptiles that, in discordance with common beliefs, are capable of producing sounds, were compiled (Supplementary Data 1).
Phylogenies
Two different phylogenies were used in this work. In order to analyze the origins of acoustic communication among choanate vertebrates, we used the tree from ref. 57, modified by ref. 4. Besides including representatives of all clades of tetrapods in the family level in a proportional sampling, it matches the available information on acoustic communication for 1799 species4. We made a minor modification to the tree by including Dipnoi as the extant outgroup to Tetrapoda (Choanata). We used the lungfish (Lepidosiren paradoxa) as the sister taxon to all tetrapods and inserted a branch length of 407mybp, based on Eoactinistia foreyi, the oldest coelacanthimorph, from the Devonian57,65.
Although the position of turtles among reptiles is still debated66,67,68, we decided to position it as the sister taxa to archosaurians, as most commonly recovered by recent molecular work69,70.
In the second analysis, we used the turtle phylogeny proposed by Pereira et al.71. This is not the most recent phylogeny available72, but it is the one with the largest overlap with our dataset. In any case, the relationship among genera is the same in both trees. We used the function drop.tip from the Ape package73 in R platform74 to exclude terminals. A tree containing each living turtle genus was created and used to analyze the distribution of sound production among turtles.
Ancestral-state reconstruction
We based our analysis for choanate vertebrates on the dataset compiled by ref. 4. We reassigned character states based on the information gathered in our literature search and our own recordings: 0 for the absence of acoustic communication (which is, in many cases, no more than the absence of information) and 1 for presence. The same analysis was used for the turtle genera tree. Character states assigned to each species can be found in Supplementary Tables 5, 6, respectively.
Considering the great amount of missing data regarding turtle vocal behaviours, we inferred the presence of acoustic communication to a genus whenever at least one of its representatives is known to do so. The evolution of acoustic communication was inferred for each ancestral node across-tree using maximum-likelihood reconstruction, and the equal-rates model (ER).
Reporting summary
Further information on research design is available in the Nature Research Reporting Summary linked to this article.
Data availability
The authors declare that the data supporting the findings of this study are available within the paper and/or its supplementary information files: Supplementary Data 1 contains the list of species and corresponding sources obtained in the literature search. Sound repertoires of the species recorded in the present work can be found in Figs. 1, 2 and in detail in Supplementary Data 2 or in an online interactive presentation (shorturl.at/cwMU2). Supplementary Data 3 contains the description of the repertoires and the conditions in which each species was recorded. Supplementary Note 1 includes the PRISMA workflow with the methods used in the literature search, together with the resulting list of references. The literature search was conducted through the platforms Web of Science (https://clarivate.com/webofsciencegroup/solutions/web-of-science/) and Google scholar (https://scholar.google.com/). Supplementary Data 4, 5 contain the character coding information used in the choanate and the turtle analyses, respectively. Code and input files can be found in Supplementary Code 1.
Code availability
This study used available R packages rather than custom coding. Nevertheless, we provide code and input files in the supplementary material. All code used for this work is provided in Supplementary Code 1.
References
Wever, E. G (ed.). in The Reptile Ear (Princeton Univ. Press, 1978).
Wever, E. G. The Amphibian Ear (Princeton Univ. Press, 1985).
Russell, A. P. & Bauer, A. M. Vocalization by extant nonavian reptiles: a synthetic overview of phonation and the vocal apparatus. Anat. Rec. 304, 1478–1528 (2021).
Chen, Z. & Wiens, J. J. The origins of acoustic communication in vertebrates. Nat. Commun. 11, 1–8 (2020).
Bass, A. H., Gilland, E. H. & Baker, R. Evolutionary origins for social vocalization in a vertebrate hindbrain-spinal compartment. Science 321, 417–421 (2008).
Barth, F. G., Giampieri-Deutsch, P. & Klein, H. D. Sensory Perception: Mind and Matter (Springer, 2012).
Nothwang, H. G. Evolution of mammalian sound localization circuits: a developmental perspective. Prog. Neurobiol. 141, 1–24 (2016).
Pfaff, C., Schultz, J. A. & Schellhorn, R. The vertebrate middle and inner ear: a short overview. J. Morphol. 280, 1098–1105 (2019).
Barkan, C. L. & Zornik, E. Inspiring song: the role of respiratory circuitry in the evolution of vertebrate vocal behavior. Dev. Neurobiol. 80, 31–41 (2020).
Gerhardt, H. C. The evolution of vocalization in frogs and toads. Ann. Rev. Ecol. Syst. 25, 293-324 (1994).
Naguib, M., Diehl, J., van Oers, K. & Snijders, L. Repeatability of signaling traits in the avian dawn chorus. Front. Zool. 16, 1–11 (2019).
Derryberry, E. P. et al. Ecological drivers of song evolution in birds: disentangling the effects of habitat and morphology. Ecol. Evol. 8, 1890–1905 (2018).
Thomson, S. A. Turtles of the world: annotated checklist and atlas of taxonomy, synonymy, distribution, and conservation status. Phyllomedusa: J. Herpetol. 20, 225–228 (2021).
M’Donnell, R. Observations on the habits and anatomy of the Lepidosiren annectans. Nat. Hist. Rev. 7, 93–112 (1860).
Christensen, C. B., Christensen-Dalsgaard, J. & Madsen, P. T. Hearing of the African lungfish (Protopterus annectens) suggests underwater pressure detection and rudimentary aerial hearing in early tetrapods. J. Exp. Biol. 218, 381–387 (2015).
Largen, M. J., Morris, P. A. & Yalden, D. W. Observations on the caecilian Geotrypetes grandisonae Taylor (Amphibia Gymnophiona) from Ethiopia: Pubblicazioni del centro di studio per la faunistica ed ecologia tropicali del CNR: LX. Monitore Zoologico Italiano. Supplemento 4, 185–205 (1972).
Thurow, G. R. & Gould, H. J. Sound production in a caecilian. Herpetologica 33, 234–237 (1977).
Duellman, W. E. & Trueb, L. Biology of Amphibians (JHU Press, 1994).
Colafrancesco, K. C., & Gridi-Papp, M. in Vertebrate Sound Production and Acoustic Communication (eds Popper,A. N., Fay, R. R., Suthers, R. A., Tecumseh Fitch, W.) Ch. 3 (Springer, 2016).
Maslin, T. P. The production of sound in caudate Amphibia. Ser. Biol. 1, 29–45 (1950).
Hubáček, J., Šugerková, M. & Gvoždík, L. Underwater sound production varies within not between species in sympatric newts. PeerJ 7, e6649 (2019).
Bedi, A., Bedi, V., Nag, S. & Suyesh, R. Wild observations of the reproductive behaviour and first evidence of vocalization in Crocodile newt Tylototriton himalayanus (Caudata: Salamandridae) from the Himalayan biodiversity hotspot in Eastern India. Salamandra 57, 65–74 (2021).
Stephenson, B. & Verrell, P. Courtship and mating of the tailed frog (Ascaphus truei). J. Zool. 259, 15–22 (2003).
Santos, M. T. T. et al. Complex acoustic signals in Crossodactylodes (Leptodactylidae, Paratelmatobiinae): a frog genus historically regarded as voiceless. Bioacoustics 30, 1–16 (2021).
Bell, B. D. Observations on the ecology and reproduction of the New Zealand Leiopelmid frogs. Herpetologica 34, 340–354 (1978).
Green, D. M. Antipredator behaviour and skin glands in the New Zealand native frogs, genus Leiopelma. N.Z. J. Zool. 15, 39–45 (1988).
Manley, G. A. Evolution of structure and function of the hearing organ of lizards. J. Neurobiol. 53, 202–211 (2002).
Capshaw, G., Willis, K. L., Han, D., & Bierman, H. S. in Neuroendocrine Regulation of Animal Vocalization. (eds Rosenfeld, C. S. & Hoffmann, F.) Ch. 7 (Academic Press, 2021).
Wojtusiak, R. J. & Majlert, Z. Bioacoustics of the voice of the tuatara, Sphenodon punctatus punctatus. N.Z. J. Sci. 16, 305–313 (1973).
Ferrara, C. R., Vogt, R. C. & Sousa-Lima, R. S. Turtle vocalizations as the first evidence of posthatching parental care in chelonians. J. Comp. Psychol. 127, 24 (2013).
Ferrara, C. R., Vogt, R. C., Giles, J. C. & Kuchling, G. in Biocommunication of Animals (ed. Witzany, G) Ch. 15, 261–274 (Springer, 2014).
Lacroix, C., Davy, C. M. & Rollinson, N. Hatchling vocalizations and beneficial social interactions in subterranean nests of a widespread reptile. Anim. Behav. 187, 233–244 (2022).
Goller, F. & Larsen, O. N. A new mechanism of sound generation in songbirds. Proc. Natl Acad. Sci. USA 94, 14787 (1997).
Kingsley, E. P. et al. Identity and novelty in the avian syrinx. Proc. Natl Acad. Sci. USA 115, 10209–10217 (2018).
Weishampel, D. B. Acoustic analyses of potential vocalization in lambeosaurine dinosaurs (Reptilia: Ornithischia). Paleobiology 7, 252–261 (1981).
Farke, A. A., Chok, D. J., Herrero, A., Scolieri, B. & Werning, S. Ontogeny in the tube-crested dinosaur Parasaurolophus (Hadrosauridae) and heterochrony in hadrosaurids. PeerJ 1, e182 (2013).
Au, W. W. & Simmons, J. A. Echolocation in dolphins and bats. Phys. Today 60, 40 (2007).
Fitch, W. T. in Vertebrate Sound Production and Acoustic Communication (eds Suthers, R. A., Fitch, W. T., Popper, A. N. & Fay, R. R.) Ch. 10 (Springer, 2016).
Clarke, J. A. et al. Fossil evidence of the avian vocal organ from the Mesozoic. Nature https://doi.org/10.1038/nature19852 (2016).
Policht, R., Kowalczyk, A., Łukaszewicz, E. & Hart, V. Hissing of geese: caller identity encoded in a non-vocal acoustic signal. PeerJ 8, e10197 (2020).
Bass, A. H. Central pattern generator for vocalization: is there a vertebrate morphotype? Curr. Opin. Neurobiol. 28, 94–100 (2014).
Bass, A. H. & Chagnaud, B. P. Shared developmental and evolutionary origins for neural basis of vocal–acoustic and pectoral–gestural signaling. Proc. Natl Acad. Sci. USA 109, 10677–10684 (2012).
Feng, N. Y., Fergus, D. J. & Bass, A. H. Neural transcriptome reveals molecular mechanisms for temporal control of vocalization across multiple timescales. BMC Genomics 16, 1–25 (2015).
Young, B. A. A review of sound production and hearing in snakes, with a discussion of intraspecific acoustic communication in snakes. J. Pa. Acad. Sci. 71, 39–46 (1997).
Baeckens, S. Evolution of animal chemical communication: insights from non-model species and phylogenetic comparative methods. Belg. J. Zool. 149 (2019).
Fernandes, D. C. & Passos, D. C. The voices of an alleged mute: sound emissions in a Tropidurus lizard. Behaviour 1, 1–10 (2021).
Milton, T. H. Structure and Social Significance of Vocalization by Anolis Grahami Grahami (Sauria: Iguanidae). Doctoral dissertation, Virginia Polytechnic Institute and State University. (1974).
Senter, P. Voices of the past: a review of Paleozoic and Mesozoic animal sounds. Historical Biol. 20, 255–287 (2008).
Aubret, F. & Mangin, A. The snake hiss: potential acoustic mimicry in a viper–colubrid complex. Biol. J. Linn. Soc. 113, 1107–1114 (2014).
Kabir, M. S. & Thaker, M. Does the addition of a new signalling trait enhance receiver responses in diurnal geckos? Behav. Process. 184, 104322 (2021).
Albersheim-Carter, J. et al. Testing the evolutionary conservation of vocal motoneurons in vertebrates. Respir. Physiol. Neurobiol. 224, 2–10 (2016).
Ladich, F. & Winkler, H. Acoustic communication in terrestrial and aquatic vertebrates. J. Exp. Biol. 220, 2306–2317 (2017).
Rice, A. N. et al. Evolutionary patterns in sound production across fishes. Ichthyol. Herpetol. 110, 1–12 (2022).
Lambertz, M. The vestigial lung of the coelacanth and its implications for understanding pulmonary diversity among vertebrates: new perspectives and open questions. R. Soc. open Sci. 4, 171518 (2017).
Lambertz, M. & Perry, S. F. in Phylogeny, Anatomy and Physiology of Ancient Fishes. (eds Zaccone, G., Dabrowski, K., Hedrick, M. S., Fernandes, J. M., & Icardo, J. M.) Ch. 9 (CRC Press, 2015).
Longo, S., Riccio, M. & McCune, A. R. Homology of lungs and gas bladders: insights from arterial vasculature. J. Morphol. 274, 687–703 (2013).
Alfaro, M. E. et al. Nine exceptional radiations plus high turnover explain species diversity in jawed vertebrates. Proc. Nat. Acad. Sci. USA 106, 13410–13414 (2009).
Shubin, N., Tabin, C. & Carroll, S. Deep homology and the origins of evolutionary novelty. Nature 457, 818–823 (2009).
Zhou, C. F., Bhullar, B. A. S., Neander, A. I., Martin, T. & Luo, Z. X. New Jurassic mammaliaform sheds light on early evolution of mammal-like hyoid bones. Science 365, 276–279 (2019).
Müller, J., Bickelmann, C. & Sobral, G. The evolution and fossil history of sensory perception in amniote vertebrates. Annu. Rev. Earth Planet. Sci. 46, 495–519 (2018).
K. Lisa Yang Center for Conservation Bioacoustics. Raven Pro: interactive sound analysis software (Version 1.6.1) [Computer software] (The Cornell Lab of Ornithology, 2019).
Boersma, P. Praat: doing phonetics by computer. http://www.praat.org/. (2006)
Moher, D., Altman, D. G., Liberati, A. & Tetzlaff, J. PRISMA statement. Epidemiology 22, 128 (2011).
Ax, P. Multicellular Animals. Order in Nature – System Made by Man Vol. III (Springer, 2003).
Johanson, Z., Long, J. A., Talent, J. A., Janvier, P. & Warren, J. W. Oldest coelacanth, from the Early Devonian of Australia. Biol. Lett. 2, 443–446 (2006).
Werneburg, I. & Sánchez-Villagra, M. R. Timing of organogenesis support basal position of turtles in the amniote tree of life. BMC Evolut. Biol. 9, 1–9 (2009).
Bever, G. S., Lyson, T. R., Field, D. J. & Bhullar, B. A. S. Evolutionary origin of the turtle skull. Nature 525, 239–242 (2015).
Joyce, W. G. The origin of turtles: a paleontological perspective. J. Exp. Zool. Part B: Mol. Dev. Evol. 324, 181–193 (2015).
Chiari, Y., Cahais, V., Galtier, N. & Delsuc, F. Phylogenomic analyses support the position of turtles as the sister group of birds and crocodiles (Archosauria). BMC Biol. 10, 1–15 (2012).
Field, D. J. et al. Toward consilience in reptile phylogeny: miRNAs support an archosaur, not lepidosaur, affinity for turtles. Evol. Dev. 16, 189–196 (2014).
Pereira, A. G., Sterli, J., Moreira, F. R. & Schrago, C. G. Multilocus phylogeny and statistical biogeography clarify the evolutionary history of major lineages of turtles. Mol. Phylogenet. Evol. 113, 59–66 (2017).
Thomson, R. C., Spinks, P. Q., & Shaffer, H. B. A global phylogeny of turtles reveals a burst of climate-associated diversification on continental margins. Proc. Natl Acad. Sci. USA 118, e2012215118 (2021).
Paradis, E., Claude, J. & Strimmer, K. APE: analyses of phylogenetics and evolution in R language. Bioinformatics 20, 289–290 (2004).
R Core Team. R: a language and environment for statistical computing (R Foundation for Statistical Computing, 2018).
Acknowledgements
Swiss Government Excellence Scholarship (ESKAS) supported G.J.-C. (Grant number: 2020.0190). This work was supported by SNF Grant No. 31003A‑169395 to M.R.S.-V. and by the Federal Commission for Scholarships for Foreign Students (FCS, Switzerland) to G.J.-C.
We thank Belize Foundation for Research and Environmental Education (BFREE) and Patrik Viana for Dermatemys and Typhlonectes photos, respectively; We thank Chester Zoo—Gerardo Garcia and the herpetology team, Turtle Island—Specially Maddy Wheatley and Lisa Marschnig, Centro de Estudos de Quelônios Amazônicos (CEQUA), Jansen Zuanon, Patrik Viana and Danilo Castanho for providing access to specimens. We also thank Dr. Torsten Scheyer and Lucas Sacheli Santos Parra for revising the manuscript and helping with the figures, respectively.
Author information
Authors and Affiliations
Contributions
All Authors gave substantial contributions to the present manuscript. G.J.-C., S.W.T. and M.R.S.-V. delineated the research; L.R.P. developed the recording equipment; G.J.-C., L.R.P., A.P.V., N.K., P.P., S.M., C.R.F., J.S. and S.E. collected and analyzed sound recordings; G.J.-C. performed the coding and statistical analyses; G.J.-C., S.E., M.R.S.-V. and S.W.T. contributed with text writing; and all authors contributed to the final revision.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks W. Tecumseh Fitch and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Jorgewich-Cohen, G., Townsend, S.W., Padovese, L.R. et al. Common evolutionary origin of acoustic communication in choanate vertebrates. Nat Commun 13, 6089 (2022). https://doi.org/10.1038/s41467-022-33741-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-022-33741-8
This article is cited by
-
Midbrain node for context-specific vocalisation in fish
Nature Communications (2024)
-
A new Frontier for Organismal Biology
Biosemiotics (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
