
Abstract
Nonalcoholic steatohepatitis (NASH) is a key step in the progression of nonalcoholic fatty liver (NAFL) to cirrhosis. However, the molecular mechanisms of the NAFL-to-NASH transition are largely unknown. Here, we identify methyltransferase like 3 (METTL3) as a key negative regulator of NASH pathogenesis. Hepatocyte-specific deletion of Mettl3 drives NAFL-to-NASH progression by increasing CD36-mediated hepatic free fatty acid uptake and CCL2-induced inflammation, which is due to increased chromatin accessibility in the promoter region of Cd36 and Ccl2. Antibody blockade of CD36 and CCL2 ameliorates NASH progression in hepatic Mettl3 knockout mice. Hepatic overexpression of Mettl3 protects against NASH progression by inhibiting the expression of CD36 and CCL2. Mechanistically, METTL3 directly binds to the promoters of the Cd36 and Ccl2 genes and recruits HDAC1/2 to induce deacetylation of H3K9 and H3K27 in their promoters, thus suppressing Cd36 and Ccl2 transcription. Furthermore, METTL3 is translocated from the nucleus to the cytosol in NASH, which is associated with CDK9-mediated phosphorylation of METTL3. Our data reveal a mechanism by which METTL3 negatively regulates hepatic Cd36 and Ccl2 gene transcription via a histone modification pathway for protection against NASH progression.
Similar content being viewed by others
Introduction
Nonalcoholic fatty liver disease (NAFLD), which ranges from nonalcoholic fatty liver (NAFL) to nonalcoholic steatohepatitis (NASH), is one of the most common chronic liver diseases in both developed and developing countries owing to an increased rate of obesity1,2. Approximately 25% of patients with NAFL develop NASH3, which is characterized by hepatic steatosis, liver injury, chronic inflammation, and liver fibrosis and is a key step in the development of cirrhosis and hepatocellular carcinoma (HCC)3,4. Several mediators have been shown to regulate NASH initiation and progression, such as lipotoxicity, oxidative stress, mitochondrial dysfunction, cell death, and immune cell activation5,6. However, the molecular events that determine whether patients with NAFL develop NASH remain unknown.
A “two-hit” theory has been proposed to explain NASH pathogenesis7. The first hit, hepatic steatosis, consists of lipid accumulation in the liver due to increased free fatty acid uptake or de novo lipogenesis8,9, which sensitizes the liver to the second hit of oxidative stress, inflammation, and injury7,10,11. Approximately 59% of hepatic triglycerides in humans are derived from serum non-esterified fatty acids9, which indicates the importance of free fatty-acid uptake in the pathogenesis of NAFL and NASH. Free fatty acid uptake is increased in NAFL and NASH due to increased expression of CD3612. Knockout of Cd36 has been shown to protect against diet-induced steatosis and NASH13. The second hit may be inflammation, which drives the progression of NAFL-to-NASH10. Chemokines such as CCL2 and its receptor CCR2 are abnormally upregulated during NASH progression14,15, and inhibition of CCL2 and CCR2 has been shown to be a therapeutic approach for the treatment of NASH16,17. It is possible that the molecular drivers that coordinate steatosis and inflammation mediate the NAFL-to-NASH transition. However, these molecular drivers have not yet been identified.
Methyltransferase like 3 (METTL3) is a key RNA methyltransferase that catalyzes mRNA m6A modifications18. METTL14 and WTAP both regulate METTL318,19,20. METTL3-mediated m6A modification has been shown to participate in many biological processes, such as neurogenesis21,22, spermatogenesis23, circadian rhythms24, stem cell pluripotency25,26, postnatal development of interscapular brown adipose tissue in mice27, and islet β-cell function28, by regulating mRNA stability, mRNA splicing, and translational efficiency. In addition, METTL14 regulates neurogenesis through the modulation of histone modifications29. Recently, METTL3 has also been shown to regulate cancer progression by affecting the expression of multiple genes30,31,32,33,34. However, whether METTL3 coordinates steatosis and inflammation to mediate the NAFL-to-NASH transition is largely unknown.
Here, we have demonstrated that METTL3 is a key repressor of the NAFL-to-NASH transition. Hepatocyte-specific deletion of Mettl3 drives the progression of NAFL-to NASH in HFD-fed mice by promoting CD36-mediated hepatic free fatty acid uptake and CCL2-induced inflammation. Mettl3-HKO mice also promote MCD-induced NASH, whereas hepatocyte-specific overexpression of Mettl3 protects against MCD-induced NASH. Mechanistically, METTL3 directly binds to the promoters of the Cd36 and Ccl2 genes and recruits HDAC1/2, which causes deacetylation of H3K9 and H3K27 in their promoters; this, in turn, suppresses the transcription of Cd36 and Ccl2. Furthermore, nuclear METTL3 is decreased in NASH, which is likely owing to CDK9-mediated phosphorylation of METTL3. These data reveal a mechanism by which METTL3 represses the transcription of Cd36 and Ccl2 via histone modification. This study also suggests that METTL3 is a negative regulator of NASH pathogenesis and may serve as a drug target for the treatment of NASH.
Results
Nuclear METTL3 is decreased in NASH livers
db/db (leptin receptor deficiency) mice exhibit severe NAFL but not NASH, whereas a NASH or MCD diet is able to induce NASH in mice. To identify potential regulators that are responsible for the NAFL-to-NASH transition, we assessed two previously published RNA-seq data sets deposited in the Gene Expression Omnibus (GEO) (GEO DataSets: GSE43314 and GSE119340) from WT VS db/db mouse livers and NC VS NASH mouse livers35,36,37. Some genes showed opposite patterns (db/db VS WT fold change >3, and NASH VS NC fold change < 1) in these two data sets, and thus, they may play an important role in the NAFL-to-NASH transition. Among these genes, we noted that Mettl3, an RNA methyltransferase-encoding gene, was dramatically upregulated (fold change = 3.945455) in db/db mouse livers but was not upregulated (fold change = 0.88228) in NASH mouse livers, which indicates that METTL3 may regulate NASH progression.
To confirm the RNA-seq data, we performed qPCR assays. Mettl3 mRNA levels were increased in livers from both db/db and HFD-fed mice but were not increased in the livers of MCD-fed mice (Supplementary Fig. 1a–c). METTL3 was primarily located in the nucleus18,38. To test whether METTL3 displays different subcellular locations in NAFL and NASH, we measured the METTL3 protein levels in the nuclei, cytosol, and total cell lysates from the livers of db/db, HFD-fed, and MCD-fed mice by immunoblotting. As shown in Fig. 1a and Supplementary Fig. 1d, METTL3 protein levels in the nuclei, cytosol, and total cell lysates from the livers of db/db mice were largely increased. Similarly, HFD feeding significantly increased METTL3 protein levels in the nuclei, cytosol, and total cell lysates by more than twofold (Fig. 1b and Supplementary Fig. 1e). This observation is consistent with a previous study39. However, METTL3 protein levels in the nuclei of MCD-fed mouse livers were dramatically decreased by 80% but were significantly increased by 10.9-fold in the cytosol (Fig. 1c and Supplementary Fig. 1f). Consistently, in human patients with NASH, we observed similar phenotypes (Fig. 1d and Supplementary Fig. 1g). METTL3 protein levels in the nuclei of NASH livers were dramatically decreased by 71.2% but were significantly increased by 2.84-fold in the cytosol (Fig. 1d and Supplementary Fig. 1g). These data indicate that downregulation of nuclear METTL3 protein levels may contribute to the NAFL-to-NASH transition.

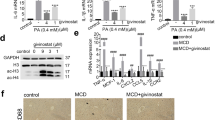
a–c METTL3 protein levels in nuclei, cytosol, and total cell lysate from the livers of db/db (11 weeks old) mice and HFD (for 8 weeks)- or MCD (for 3 weeks)-fed mice were measured by immunoblotting. The quantification was shown in Supplementary Fig. 1d–f. d Immunoblotting of METTL3 protein levels in nuclei, cytosol, and total cell lysate from human NASH and normal liver tissues. The quantification was shown in Supplementary Fig. 1g. e Primary hepatocytes were isolated and treated with TNFα (20 ng/ml) for 2 h. METTL3 protein levels in nuclei, cytosol, and total cell lysate were measured by immunoblotting. The quantification was shown in Supplementary Fig. 1i. f Primary hepatocytes were infected with Ad-Flag-METTL3 adenovirus, and then treated with or without TNFα (20 ng/ml) for 2 h. The total cell lysate was immunoprecipitated with anti-phosphoserine antibody and immunoblotted with anti-Flag antibody. g Myc-CDK9 expression vector was co-transfected with or without Flag-METTL3 expression vector in HEK293T cells. Total cell lysates were immunoprecipitated with Flag beads and then immunoblotted with anti-Myc or anti-Flag antibodies. h In vitro kinase assay. i Primary hepatocytes were treated with vehicle, TNFα (20 ng/ml), and TNFα (20 ng/ml) plus BAY-1143572 (2 μM) for 2 h. In TNFα plus BAY-1143572 group, primary hepatocytes were pretreated with BAY-1143572 (2 μM) for 2 h. METTL3 protein levels in nuclei, cytosol, and total cell lysate were measured by immunoblotting. CDK9, p-CDK9, Lamin B1, and Tubulin levels were also measured by immunoblotting. The quantification was shown in Supplementary Fig. 1m–n. The samples were derived from the same experiment and the blots were processed in parallel. n was the number of biologically independent mice or cell samples. The cell culture experiments were repeated three times independently with similar results. Source data are provided as a Source Data file.
To further test how METTL3 is translocated from the nucleus to the cytosol in NASH, primary hepatocytes were isolated and treated with palmitic acid (PA) or TNFα, as it is known that PA administration can mimic fatty liver disease and that TNFα can induce liver inflammation40,41. As shown in Supplementary Fig. 1h, PA did not induce the translocation of METTL3, whereas TNFα was able to increase cytosolic METTL3 accumulation and decrease nuclear METTL3 levels (Fig. 1e and Supplementary Fig. 1i). Phosphorylation has been shown to regulate nuclear localization42,43. We observed that TNFα was able to induce the serine phosphorylation of METTL3 (Fig. 1f), which may contribute to the inhibition of METTL3 nuclear localization. Next, we investigated which protein kinase was responsible for the mediation of TNFα-induced phosphorylation of METTL3. Protein interaction data in the neXtProt human protein knowledgebase and IntAct database showed that two protein kinases, CDK9 and PRKDC, might interact with METTL344,45. Co-immunoprecipitation (Co-IP) experiments performed in HEK293T cells showed that CDK9 interacted with METTL3 (Fig. 1g), and an in vitro kinase assay showed that CDK9 was able to phosphorylate METTL3 (Fig. 1h). We observed that TNFα also induced the phosphorylation of CDK9 (Supplementary Fig. 1j and Fig. 1i), and p-CDK9 levels were increased in NASH livers (Supplementary Fig. 1k–l), which further indicates that CDK9 may be involved in the nuclear/cytosolic translocation of METTL3 in NASH. Next, we asked whether inhibition of CDK9 can block the TNFα-induced cytosolic accumulation of METTL3. As shown in Fig. 1i and Supplementary Fig. 1m–n, BAY-1143572, a CDK9 inhibitor, blocked both TNFα-induced p-CDK9 and the nuclear/cytosolic translocation of METTL3. These data indicate that TNFα/CDK9-mediated phosphorylation of METTL3 may contribute to the reduction of nuclear METTL3 levels in NASH livers.
Hepatic deletion of Mettl3 exacerbates both HFD and MCD -induced NASH
To determine whether METTL3 regulates the NAFL-to-NASH transition, we generated hepatocyte-specific Mettl3 knockout (HKO) mice by crossing Mettl3flox/flox mice with Alb-Cre transgenic mice. The genotype of the Mettl3-HKO mice was Mettl3flox/flox Alb-Cre+/−. As expected, METTL3 protein levels were dramatically decreased by 93.2% in the livers of Mettl3-HKO mice (Fig. 2a). We did not observe any differences in body weights (Supplementary Fig. 2a), serum triacylglycerol (TAG) levels (Supplementary Fig. 2b), liver weights (Supplementary Fig. 2c), or liver TAG levels (Supplementary Fig. 2d) between Mettl3flox/flox and Alb-Cre mice. Mettl3flox/flox mice also displayed similar MCD-induced NASH as Alb-Cre mice, as revealed by similar body weight (Supplementary Fig. 2e), serum TAG levels (Supplementary Fig. 2f), liver weights (Supplementary Fig. 2g), and liver TAG levels (Supplementary Fig. 2h). Therefore, we used Mettl3flox/flox mice as the control for Mettl3-HKO mice in the following experiments. When normal chow was provided, the liver weights and TAG levels were comparable between Mettl3-HKO and Mettl3flox/flox mice at 11 weeks old (Fig. 2b, c). However, after they were fed an HFD for 12 weeks, Mettl3-HKO mice displayed more severe NAFL, as revealed by higher serum ALT activities (Fig. 2d), higher liver weights (Fig. 2e, f), more hepatic lipid droplets (Fig. 2f), and higher liver TAG levels (Fig. 2g) compared with Mettl3flox/flox mice. To further test whether METTL3 is involved in NASH progression, Mettl3-HKO and Mettl3flox/flox mice were fed an MCD for 3 weeks. Consistently, Mettl3-HKO mice also exhibited higher serum ALT activities (Fig. 2h), higher liver weights (Fig. 2i, j), more hepatic lipid droplets (Fig. 2j), and higher liver TAG levels (Fig. 2k), which indicates that Mettl3-HKO mice are more sensitive to MCD-induced NASH.

a METTL3 protein levels in livers of Mettl3flox/flox and Mettl3-HKO mice at 8 weeks old (n = 3 for each group; P = 0.0059). The samples were derived from the same experiment and the blots were processed in parallel. b Liver weights of Mettl3flox/flox and Mettl3-HKO mice at 11 weeks old (n = 9 for each group). c Liver TAG levels in Mettl3flox/flox and Mettl3-HKO mice at 11 weeks old (Mettl3flox/flox, n = 9; Mettl3-HKO, n = 8). d Serum ALT activity in Mettl3flox/flox and Mettl3-HKO mice fed an HFD for 12 weeks (Mettl3flox/flox, n = 14; Mettl3-HKO, n = 13; P = 0.0332). e Liver weights of Mettl3flox/flox and Mettl3-HKO mice fed an HFD for 12 weeks (Mettl3flox/flox, n = 8; Mettl3-HKO, n = 10; P = 0.0135). f Representative pictures and H&E staining of livers from Mettl3flox/flox and Mettl3-HKO mice fed an HFD for 12 weeks. g Liver TAG levels in Mettl3flox/flox and Mettl3-HKO mice fed an HFD for 12 weeks (Mettl3flox/flox, n = 9; Mettl3-HKO, n = 11; P = 0.0007). h Serum ALT activity in Mettl3flox/flox and Mettl3-HKO mice fed an MCD for 3 weeks (Mettl3flox/flox, n = 9; Mettl3-HKO, n = 10; P = 0.0046). i Liver weights of Mettl3flox/flox and Mettl3-HKO mice fed an MCD for 3 weeks (Mettl3flox/flox, n = 8; Mettl3-HKO, n = 7; P = 0.0127). j Representative pictures and H&E staining of livers from Mettl3flox/flox and Mettl3-HKO mice fed an MCD for 3 weeks. k Liver TAG levels in Mettl3flox/flox and Mettl3-HKO mice fed an MCD for 3 weeks (Mettl3flox/flox, n = 9; Mettl3-HKO, n = 10; P = 0.0006). l Sirius Red staining of liver sections from Mettl3flox/flox and Mettl3-HKO mice fed an HFD for 12 weeks (n = 6 for each group; P = 0.0002). m F4/80 immunostaining of liver sections from Mettl3flox/flox and Mettl3-HKO mice fed an HFD for 12 weeks (n = 4 for each group; P = 0.0439). n RT-qPCR analysis of mRNA levels in the livers of Mettl3flox/flox and Mettl3-HKO mice fed an HFD for 12 weeks (Mettl3flox/flox, n = 10–15; Mettl3-HKO, n = 12–14; Collagen1A1, P = 0.0061; αSMA, P = 0.0163; Mmp9, P = 0.0884; Tgfb1, P = 0.0001). o Sirius Red staining of liver sections from Mettl3flox/flox and Mettl3-HKO mice fed an MCD for 3 weeks (Mettl3flox/flox, n = 5; Mettl3-HKO, n = 6; P = 0.00004). p F4/80 immunostaining of liver sections from Mettl3flox/flox and Mettl3-HKO mice fed an MCD for 3 weeks (n = 5 for each group; P = 0.0446). q RT-qPCR analysis of mRNA levels in the livers of Mettl3flox/flox and Mettl3-HKO mice fed an MCD for 3 weeks (Mettl3flox/flox, n = 10–11; Mettl3-HKO, n = 11–12; Collagen1A1, P = 0.0004; αSMA, P = 0.0015; Mmp9, P = 0.0062; Tgfb1, P = 0.0151). n was the number of biologically independent mice. Data represent the mean ± SEM. Significance was determined by unpaired two-tailed Student’s t test analysis. *P < 0.05. **P < 0.01. Source data are provided as a Source Data file.
Human patients with NASH have a high risk of developing liver fibrosis5. To address whether the increased steatohepatitis in Mettl3-HKO mice also promotes liver fibrosis and immune cell infiltration, we measured pathological collagen deposition and immune cell infiltration using Sirius Red staining and F4/80 immunostaining, respectively. Liver sections from Mettl3-HKO mice contained significantly larger Sirius Red-positive areas than Mettl3flox/flox mice under both HFD- and MCD-feeding conditions (Fig. 2l, o). Liver sections from Mettl3-HKO mice contained significantly more F4/80-positive areas than Mettl3flox/flox mice under both HFD- and MCD-feeding conditions (Fig. 2m, p). Consistently, the expression of fibrosis markers (collagen IA1 and aSMA) and profibrogenic factor (Tgfb1) was significantly increased in Mettl3-HKO mice fed either an HFD (Fig. 2n) or MCD (Fig. 2q). These data indicate that hepatic deletion of Mettl3 accelerates the progression from NASH to liver fibrosis.
Hepatic deletion of Mettl3 accelerates free fatty-acid uptake by increasing CD36 expression
Hepatic lipid accumulation contributes to NASH progression5,11. Liver steatosis results from an imbalance among free fatty acid uptake, lipogenesis, fatty acid β-oxidation, and very low-density lipoprotein (VLDL) secretion8,46. To determine which process is responsible for hepatic lipid accumulation in Mettl3-HKO mice, the expression of genes related to free fatty acid uptake (Fatp2, Fatp5, and Cd36), fatty acid β-oxidation (Cpt1α, Mcad, and Ppara), lipogenesis (Fasn, Scd1, Srebp1, Chrebp, Pparg, mtGPAT1, and Dgat1) and VLDL secretion (ApoB and Mttp) was measured by real-time quantitative PCR (RT-qPCR). As shown in Fig. 3a, Cd36 mRNA levels were dramatically increased whereas those of Fasn, Srebp1, Chrebp, Fatp2, and Fatp5 were decreased, which indicates that CD36-mediated free fatty acid uptake was increased whereas lipogenesis was decreased in the livers of Mettl3-HKO mice. Mcad and ApoB mRNA levels were slightly increased whereas Cpt1a, Ppara, and Mttp mRNA levels were not altered in the livers of Mettl3-HKO mice. These data suggest that fatty acid β-oxidation, lipogenesis, and VLDL secretion do not contribute to the increased liver steatosis seen in Mettl3-HKO mice and suggest that increased CD36-mediated hepatic free fatty-acid uptake leads to more severe NALF and NASH in Mettl3-HKO mice. Consistent with the increased mRNA levels, CD36 protein levels were also significantly increased in the livers of Mettl3-HKO mice (Fig. 3b). To further test whether free fatty acid uptake is increased in Mettl3-HKO mice, we measured free fatty-acid uptake both in vivo and in vitro. As shown in Fig. 3c, acute injection of the fluorescent palmitate analog BODIPY FL C16 into Mettl3-HKO and Mettl3flox/flox mice resulted in a 1.7-fold increase in free fatty acid uptake by the liver in Mettl3-HKO mice. To further verify whether hepatic METTL3 regulates free fatty-acid uptake in a cell-autonomous manner, primary hepatocytes were isolated from Mettl3-HKO and Mettl3flox/flox mice, and BODIPY FL C16 uptake experiments were performed. As shown in Fig. 3d, free fatty-acid uptake was significantly increased by 2.1-fold according to the measurement of intracellular fluorescence of BODIPY FL C16. These data demonstrate that increased CD36-mediated free fatty acid uptake contributes to the increased liver steatosis seen in Mettl3-HKO mice.

a Relative mRNA levels in livers of Mettl3flox/flox and Mettl3-HKO mice at 8 weeks old were determined by RT-qPCR (Mettl3flox/flox, n = 12–13; Mettl3-HKO, n = 11; Fatp2, P = 0.0171; Fatp5, P = 0.0029; Cd36, P < 0.00001; Cpt1α, P = 0.4018; Mcad, P = 0.0025; Pparα, P = 0.2124; Fasn, P = 0.0393; Scd1, P = 0.5119; Srebp1, P = 0.02599; Chrebp, P = 0.00298; Pparg, P = 0.6043; mtGPAT1, P = 0.358; Dgat1, P = 0.1546; ApoB, P = 0.0453; Mttp, P = 0.0594). b CD36 protein levels in Mettl3flox/flox and Mettl3-HKO mice at 8 weeks age were measured by immunoblotting. CD36 protein levels were quantified by ImageJ and normalized to Tubulin (n = 3 for each group; P = 0.0014). The samples were derived from the same experiment and the blots were processed in parallel. c Representative BODIPY FL C16 fluorescence image and relative lipid uptake levels in Mettl3flox/flox and Mettl3-HKO mice at 8 weeks old (Mettl3flox/flox, n = 7; Mettl3-HKO, n = 6; P = 0.02). d Representative BODIPY FL C16 fluorescence image and relative lipid uptake levels in primary hepatocytes isolated from Mettl3flox/flox and Mettl3-HKO mice at 8 weeks old (n = 5 for each group; P = 0.00001). n was the number of biologically independent mice or cell samples. The cell culture experiments were repeated for three times independently with similar results. Data represent the mean ± SEM. Significance was determined by unpaired two-tailed Student’s t test analysis. *P < 0.05. **P < 0.01. Source data are provided as a Source Data file.
Liver-specific knockout of Mettl3 enhances liver injury
Increased hepatic steatosis is not sufficient to induce NASH5. For this reason, an HFD alone cannot induce NASH. Surprisingly, HFD feeding was sufficient to induce NASH in Mettl3-HKO mice. Liver injury and inflammation have been shown to be key regulators of the NAFL-to-NASH transition10. Serum ALT activities were significantly increased in Mettl3-HKO mice fed either an HFD or MCD, which suggests that hepatic deletion of Mettl3 accelerates diet-induced liver injury and inflammation. We measured hepatocyte apoptosis using terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) assays in both HFD- and MCD-fed mice. The number of TUNEL-positive cells was significantly increased in both HFD- and MCD- fed Mettl3-HKO mice (Fig. 4a, b). In addition, cleaved caspase3 and its activity were much higher in Mettl3-HKO mice (Fig. 4c, d), which indicates that Mettl3-HKO mice displayed more severe liver injury after they were fed an HFD or MCD.

a The TUNEL-positive cells in Mettl3flox/flox and Mettl3-HKO mice fed an HFD for 12 weeks (n = 5 for each group; P = 0.0474). b The TUNEL-positive cells in Mettl3flox/flox and Mettl3-HKO mice fed an MCD for 3 weeks (n = 5 for each group; P = 0.0027). c Cleaved caspase3 levels were measured by immunoblotting in the livers of Mettl3flox/flox and Mettl3-HKO mice fed either an HFD or MCD. Cleaved caspase3 levels were quantified by ImageJ and normalized to Tubulin (n = 3 for each group; HFD, Mettl3flox/flox versus Mettl3-HKO, P = 0.0197; MCD, Mettl3flox/flox versus Mettl3-HKO, P = 0.0013). The samples were derived from the same experiment and the blots were processed in parallel. d Caspase3 activity in the livers of Mettl3flox/flox and Mettl3-HKO mice fed either an HFD or MCD was measured by using a Caspase3 Assay Kit (ab39383, Abcam) following the instruction (Mettl3flox/flox, n = 6 for HFD, n = 5 for MCD; Mettl3-HKO, n = 5; HFD, Mettl3flox/flox versus Mettl3-HKO, P = 0.0034; MCD, Mettl3flox/flox versus Mettl3-HKO, P = 0.0433). n was the number of biologically independent mice. Data represent the mean ± SEM. Significance was determined by unpaired two-tailed Student’s t test analysis. *P < 0.05. **P < 0.01. Source data are provided as a Source Data file.
Hepatic deletion of Mettl3 increases liver inflammation by enhancing CCL2 expression
To comprehensively compare the gene expression profiles in the livers of Mettl3-HKO and Mettl3flox/flox mice, we performed RNA-sequencing (RNA-seq) analysis. As shown in Fig. 5a, a total of 551 genes were upregulated, and 474 genes were downregulated. Gene Ontology (GO) analysis showed that genes related to defense response, immune system process, immune response, and innate immune response were significantly increased, whereas those associated with negative regulation of gluconeogenesis, locomotor rhythm, negative regulation of lipid storage, and digestion were downregulated (Fig. 5b). Cd36 was one of the upregulated genes, which was confirmed above by RT-qPCR and immunoblotting (Fig. 3a, b). RT-qPCR analysis further confirmed other upregulated genes. As shown in Fig. 5c–e, Ccl2, Cx3cr1, Ccr2, Ccl3, Cxcl10, and Tnfa mRNA levels were increased in Mettl3-HKO mice, but the expression levels of other inflammatory genes such as Infg, Il1b, and Il6 were unchanged. To further determine whether hepatic METTL3 regulates the expression of inflammatory genes in a cell-autonomous manner, primary hepatocytes were isolated from Mettl3-HKO and Mettl3flox/flox mice, and gene expression was measured by RT-qPCR. As shown in Fig. 5f, Ccl2 was the most upregulated gene in primary hepatocytes isolated from Mettl3-HKO mice. Furthermore, CCL2 protein levels were also significantly increased in the livers of Mettl3-HKO mice fed the three types of diets (normal chow, HFD, and MCD) (Fig. 5g). These data suggest that hepatic upregulation of CCL2 in Mettl3-HKO mice contributes to more severe NASH.

a RNA-seq analysis was performed in the livers of Mettl3flox/flox and Mettl3-HKO mice at 8 weeks old. The differentially expressed genes (DEGs) (HKO VS ff) including 474 downregulated genes and 551 upregulated genes were illustrated in a volcanoplot (|log2foldchange|>1.5 and pval < 0.05). b Top GO biological process terms enriched in downregulated and upregulated genes. c Real-time qPCR analysis of mRNA levels in the livers of Mettl3flox/flox and Mettl3-HKO mice at 8 weeks old (Mettl3flox/flox, n = 12, 13; Mettl3-HKO, n = 11; Infg, P = 0.4526; Il1b, P = 0.1869; Il6, P = 0.2102; Tnfα, P = 0.0056; Ccl2, P = 0.0024; Cxcr1, P = 0.0085; Ccr2, P = 0.5215; Ccl3, P = 0.0461; Cxcl10, P = 0.0001). d RT-qPCR analysis of mRNA levels in the livers of Mettl3flox/flox and Mettl3-HKO mice fed an HFD for 12 weeks (Mettl3flox/flox, n = 13–16; Mettl3-HKO, n = 14; Infg, P = 0.0237; Il1b, P = 0.0977; Il6, P = 0.0353; Tnfα, P = 0.0042; Ccl2, P = 0.0008; Cxcr1, P < 0.0001; Ccr2, P = 0.0008; Ccl3, P = 0.0777; Cxcl10, P < 0.0001). e RT-qPCR analysis of mRNA levels in the livers of Mettl3flox/flox and Mettl3-HKO mice fed an MCD for 3 weeks (Mettl3flox/flox, n = 11, 12; Mettl3-HKO, n = 11, 12; Infg, P = 0.7685; Il1b, P = 0.3087; Il6, P = 0.6141; Tnfα, P = 0.0033; Ccl2, P = 0.0003; Cxcr1, P = 0.0753; Ccr2, P = 0.0306; Ccl3, P = 0.2095; Cxcl10, P = 0.0381). f Real-time qPCR analysis of mRNA levels in primary hepatocytes isolated from Mettl3flox/flox and Mettl3-HKO mice at 8 weeks old (n = 6 for each group; Infg, P = 0.7077; Il1b, P = 0.1698; Il6, P = 0.6845; Tnfα, P = 0.6021; Ccl2, P < 0.0001; Cxcr1, P = 0.6532; Ccr2, P = 0.7745; Ccl3, P = 0.484; Cxcl10, P < 0.0001). g CCL2 protein levels were measured by immunoblotting in the livers of Mettl3flox/flox and Mettl3-HKO mice fed NC, HFD and MCD, respectively. CCL2 protein levels were then quantified by ImageJ and normalized to Tubulin (n = 3 for each group; NC, P = 0.0096; HFD, P = 0.0026; MCD, P = 0.0117). The samples were derived from the same experiment and the blots were processed in parallel. n was the number of biologically independent mice. Data represent the mean ± SEM. Significance was determined by unpaired two-tailed Student’s t test analysis. *P < 0.05. **P < 0.01. Source data are provided as a Source Data file.
Inhibition of CD36 and CCL2 ameliorates NASH progression in Mettl3-HKO mice
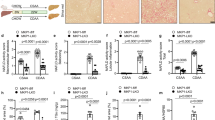
To further confirm that CD36 and CCL2 contribute to NASH progression in Mettl3-HKO mice, Mettl3-HKO mice were fed an MCD diet and treated with a combination of anti-CD36 and anti-CCL2-neutralizing antibodies. Interestingly, inhibition of CD36 and CCL2 ameliorated NASH progression in Mettl3-HKO mice, as revealed by lower serum ALT activity (Fig. 6a), lower liver weights (Fig. 6b), normal morphology of the liver (Fig. 6c), lower liver TAG levels (Fig. 6d), fewer TUNEL-positive cells (Fig. 6e), and lower caspase3 cleavage (Fig. 6f). These data demonstrate that antibody blockade of CD36 and CCL2 prevents the NASH progression in Mettl3-HKO mice and further supports the theory that elevated CD36 and CCL2 contribute to NASH progression in Mettl3-HKO mice.

Mettl3-HKO mice at 8 weeks old were treated with a combination of anti-CD36 and anti-CCL2 antibodies (4 μg/antibody/mouse, i.v.) or an equal amount of control antibodies (IgA and IgG2b) on day 1, 4, 7, and 8 after MCD feeding. Mice were killed on day 9 after MCD feeding. a Serum ALT activity (n = 6 for each group; p = 0.0204). b Liver weights (n = 6 for each group; P = 0.0325). c Representative liver pictures. d Liver TAG levels (n = 6 for each group; P = 0.0108). e TUNEL-positive cells (n = 6 for each group; P = 0.0059). f The cleavage of caspase3 was measured by immunoblotting, quantified by ImageJ, and normalized to Tubulin (n = 4 for each group; P = 0.0043). The samples were derived from the same experiment and the blots were processed in parallel. n was the number of biologically independent mice. Data represent the mean ± SEM. Significance was determined by unpaired two-tailed Student’s t test analysis. *P < 0.05. **P < 0.01. Source data are provided as a Source Data file.
Hepatic overexpression of Mettl3 ameliorates MCD-induced NASH
Next, we asked whether hepatic overexpression of Mettl3 can ameliorate MCD-induced NASH. We generated STOP-Mettl3 mice, in which a STOP-Flag-Mettl3 cassette was inserted in the Rosa26 allele using the CRISPR-Cas9 technique (Fig. 7a and Supplementary Fig. 3a). Hepatocyte-specific Mettl3-overexpressing (HOE) mice were generated by crossing STOP-Mettl3+/− mice with Alb-Cre transgenic mice. The genotype of the Mettl3-HOE mice was STOP-Mettl3+/− Alb-Cre+/−, whereas the genotype of the control mice was STOP-Mettl3+/− (Supplementary Fig. 3b). As expected, Flag-METTL3 levels were dramatically increased specifically in the livers but not in other tissues (WAT, Skeletal muscle, BAT, Heart, Spleen, Kidney, and Brain) of Mettl3-HOE mice (Fig. 7b). When the mice were fed normal chow, the serum TAG levels, liver weights, and liver TAG levels were comparable between Mettl3-HOE and their control littermates (Supplementary Fig. 3c–e). We then challenged these mice with an MCD for 2 weeks to determine whether Mettl3-HOE mice are resistant to MCD-induced NASH. Interestingly, Mettl3-HOE mice displayed resistance to MCD-induced NASH, as revealed by lower serum ALT activity (Fig. 7c), fewer hepatic lipid droplets (Fig. 7d), and lower liver TAG levels (Fig. 7e). Mettl3-HOE mice also showed a significant decrease in TUNEL-positive cells (Fig. 7f), which indicates that the hepatocyte-specific overexpression of Mettl3 could ameliorate MCD-induced liver injury. Moreover, liver sections from Mettl3-HOE mice contained fewer Sirius Red-positive areas than their control littermates (Fig. 7g). Consistently, the expression of fibrosis markers (collagen IA1 and Mmp9) and profibrogenic factor (Tgfb1) was significantly decreased in Mettl3-HOE mice (Fig. 7h). These data indicate that hepatic overexpression of Mettl3 ameliorates MCD-induced NASH and liver fibrosis.

a A schematic representation of the Rosa26-STOP-METTL3 targeting vector and generation of liver-specific overexpression of Mettl3 mice. b Tissue extracts from control and Mettl3-HOE mice were immunoblotted with antibodies to Flag and β-actin, respectively. Lane 1 Control liver; lane 2 HOE liver; lane 3 HOE White adipose tissue; lane 4 HOE Skeletal muscle; lane 5 HOE Brown adipose tissue; lane 6 HOE Heart; lane 7 HOE Spleen; lane 8 HOE Kidney, lane 9 HOE Brain. c Serum ALT activities (control, n = 10; Mettl3-HOE, n = 7; P = 0.0024). d H&E staining of liver from control and Mettl3-HOE mice fed an MCD for 2 weeks. e Liver TAG levels (control, n = 10; Mettl3-HOE, n = 7; P = 0.0243). f TUNEL-positive cells (n = 6 for each group; P = 0.00003). g Sirius Red staining of livers from control and Mettl3-HOE mice fed an MCD for 2 weeks (n = 5 for each group). h Relative mRNA levels (control, n = 7–8; Mettl3-HOE, n = 7–8; collagen1A1, P = 0.0365; αSMA, P = 0.2169; Mmp9, P = 0.0011; Tgfb1, P = 0.0072). i The relative lipid uptake in control and Mettl3-HOE mice at 8 weeks old (control, n = 9; Mettl3-HOE, n = 11; P < 0.0001). j, k Cd36 and Ccl2 mRNA levels (control, n = 10; Mettl3-HOE, n = 7; Cd36, P = 0.0121; Ccl2, P = 0.0178). l CD36 and CCL2 protein levels were measured by immunoblotting, quantified by ImageJ and normalized to Tubulin (n = 3 for each group; CD36, P = 0.0101; CCL2, P = 0.0427). n was the number of biologically independent mice. The samples were derived from the same experiment and the blots were processed in parallel. Data represent the mean ± SEM. Significance was determined by unpaired two-tailed Student’s t test analysis. *P < 0.05. **P < 0.01. Source data are provided as a Source Data file.
We then tested whether the protective effects of hepatic METTL3 overexpression against MCD-induced NASH are due to the reduction in CD36-mediated free fatty acid uptake and CCL2-associated liver inflammation. Free fatty acid uptake and the expression of Cd36 and Ccl2 were measured in Mettl3-HOE and control mice. Mettl3-HOE mice showed a significant reduction in hepatic free fatty acid uptake (Fig. 7i), which was due to a decrease in CD36 expression (Fig. 7j, l). We did not observe significant differences in the expression of genes related to TAG synthesis, fatty acid β-oxidation, or VLDL secretion in Mettl3-HOE mice (Supplementary Fig. 3f), which suggests that hepatic overexpression of Mettl3 ameliorates MCD-induced liver steatosis primarily by decreasing CD36-mediated free fatty acid uptake into the liver. In addition, CCL2 expression was significantly decreased in Mettl3-HOE mice (Fig. 7k, l). These data suggest that reductions in both CD36-mediated hepatic free fatty acid uptake and CCL2-associated liver inflammation ameliorate MCD-induced NASH in Mettl3-HOE mice.
METTL3 regulates Cd36 and Ccl2 transcription
We next sought to investigate the mechanism by which METTL3 affects the expression of Cd36 and Ccl2. METTL3-mediated m6A modification has been shown to regulate mRNA processing. To determine whether METTL3-mediated m6A modification regulates the expression of Cd36 and Ccl2, m6A RNA immunoprecipitation sequencing (m6ARIP-seq) analysis was performed in the livers of Mettl3-HKO and Mettl3flox/flox mice. Consistent with published m6ARIP-seq results, the m6A peaks identified in the livers of Mettl3flox/flox mice were enriched at the stop codon and 3′-UTR and were characterized by the presence of the canonical GGACU motif (Supplementary Fig. 4a, b). However, the m6A peaks in the livers of Mettl3-HKO mice were dispersed at the transcription start site (TSS), 5′-UTR, and stop codon, and no enriched motif was found (Supplementary Fig. 4a). In the livers of Mettl3flox/flox mice, we identified 6556 genes with m6A peaks (Supplementary Table 4), and 3489 genes exhibited decreased m6A levels in the livers of Mettl3-HKO mice (Supplementary Fig. 4c). GO analysis showed that the genes with downregulated m6A peaks were related to metabolic process, RNA metabolic process, and cellular macromolecule biosynthetic process (Supplementary Fig. 5). KEGG pathway analysis showed that the genes with downregulated m6A peaks were associated with ribosome, metabolic pathways, HIF-1-signaling pathway, pathways in cancer, Epstein-Barr virus infection, Alzheimer’s disease, ubiquitin-mediated proteolysis, glycolysis/gluconeogenesis, protein processing in the endoplasmic reticulum, prolactin signaling pathway, and NAFLD (Supplementary Fig. 6). Among genes related to NAFLD, both m6A peaks and mRNA levels of Srebp1 (also named Srebf1) were significantly decreased (Supplementary Fig. 6, Supplementary Table 4 and Fig. 3a), which further indicates that lipogenesis likely contributes little to fatty liver observed in Mettl3-HKO mice. However, we did not detect significant m6A peaks in either Cd36 or Ccl2 transcript in the livers of Mettl3flox/flox and Mettl3-HKO mice, which indicates that METTL3 regulates the expression of Cd36 or Ccl2, but that it is unlikely due to an m6A modification in their transcripts.
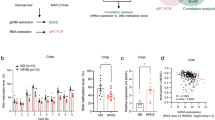
Nuclear run-on RT-qPCR showed that hepatic deletion of Mettl3 significantly increased Cd36 and Ccl2 nascent mRNA levels (Fig. 8a). These data indicate that METTL3 regulates Cd36 and Ccl2 expression at the transcriptional level independent of m6A modifications. To further test this hypothesis, we mapped open chromatin in the livers of Mettl3-HKO mice using Assay for Transposase-Accessible Chromatin with high-throughput sequencing (ATAC-seq). As expected, we observed strong enrichment of open chromatin in the gene promoters and the ATAC-seq peaks near the TSS (Supplementary Fig. 7), which are associated with the activation of gene transcription. ATAC-seq peaks were significantly increased in the promoters of Cd36 and Ccl2 in the livers of Mettl3-HKO mice (Fig. 8b, c), which indicates that deletion of Mettl3 promotes Cd36 and Ccl2 transcription. RNA-seq data also showed that both Cd36 and Ccl2 transcripts were dramatically increased in the livers of Mettl3-HKO mice (Fig. 8b, c). Furthermore, METTL3 directly bound to the Cd36 and Ccl2 promoters but did not bind to the Actb promoter in the livers of Mettl3-HOE mice, as detected by chromatin immunoprecipitation (ChIP) (Fig. 8d). In addition, promoter luciferase assays showed that METTL3 decreased both Cd36 and Ccl2 promoter-luciferase activities in a dose-dependent manner (Fig. 8e). The METTL3 enzyme-dead mutation (D395A) also reduced both Cd36 and Ccl2 promoter-luciferase activities in a dose-dependent manner (Fig. 8f). Consistently, both METTL3 and its mutation D395A significantly decreased the mRNA and protein levels of CD36 and CCL2 (Fig. 8g, h). METTL3 mutation (D395A) also dramatically decreased the expression of Cd36 and Ccl2 in isolated hepatocytes from Mettl3-HKO mice (Supplementary Fig. 8). These data demonstrate that METTL3 serves as a transcriptional repressor of both the Cd36 and Ccl2 genes independent of its methyltransferase enzyme activity.

a Nascent Cd36 and Ccl2 mRNA levels in primary hepatocytes isolated from Mettl3flox/flox and Mettl3-HKO mice (n = 5 for each group; Cd36, P = 0.0261; Ccl2, P = 0.0439). b, c ATAC-seq peaks in the promoters of Cd36 and Ccl2 genes and RNA-seq peaks of Cd36 and Ccl2 transcripts in the livers of Mettl3flox/flox and Mettl3-HKO at 8 weeks old. d Binding of Flag-METTL3 to the promoters of Cd36 and Ccl2 were assessed in the livers of control and Mettl3-HOE mice at 8 weeks old by ChIP. The liver samples were immunoprecipitated with Flag beads. Immunoprecipitated DNA was extracted for PCR analysis. Representative image and statistic analysis were shown (n = 3 for each group; Cd36, P < 0.0001; Ccl2, P = 0.0007; Actb, P = 0.666). The samples were derived from the same experiment and the gels were processed in parallel. e, f Cd36 or Ccl2 luciferase reporter plasmids were co-transfected with METTL3, D395A, or empty expression vector by polyethyleneimine (Sigma) into HEK293T cells. 48 h later, HEK293T cells were lysed in reporter lysis buffer, and luciferase activity was measured and normalized to β-Gal activity (n = 6 for each group; Cd36, P < 0.0001; Ccl2, P < 0.0001). g, h Primary hepatocytes were infected with Ad-βGal, Ad-METTL3 and Ad-METTL3 (D395A) adenovirus overnight, Cd36 and Ccl2 mRNA levels were measured by qPCR (n = 6 for each group; for Cd36, βGal versus METTL3, P = 0.0008, METTL3 versus D395A, P = 0.0006; for Ccl2, βGal versus METTL3, P = 0.0044, METTL3 versus D395A, P = 0.0042). CD36 and CCL2 protein levels were measured by immunoblotting, quantified by ImageJ and normalized to Tubulin (n = 3 for each group; for CD36, βGal versus METTL3, P = 0.0111, METTL3 versus D395A, P = 0.0134; for CCL2, βGal versus METTL3, P = 0.0012, METTL3 versus D395A, P = 0.0006). The samples were derived from the same experiment and the blots were processed in parallel. n was the number of biologically independent mice or cell samples. The cell culture experiments were repeated three times independently with similar results. Data represent the mean ± SEM. Significance was determined by unpaired two-tailed Student’s t test analysis. *P < 0.05. **P < 0.01. Source data are provided as a Source Data file.
METTL3 interacts with HDAC1/2 and regulates H3K9ac and H3K27ac levels in the promoters of Cd36 and Ccl2
We next investigated how METTL3 functions as a transcriptional repressor. Protein interaction data in the neXtProt human protein knowledgebase and IntAct database showed that METTL3 might interact with HDAC1 and HDAC244,45. HDAC1 and HDAC2 are known to repress the expression of target genes by deacetylating histone H3K9 and H3K27. We found that METTL3 coimmunoprecipitated with either HDAC1 or HDAC2 in both HEK293T cells (Fig. 9a, b) and livers from Mettl3-HOE mice (Fig. 9c). These interactions were decreased in MCD-induced NASH (Fig. 9d). Immunofluorescence data showed that Flag-METTL3 was translocated from the nucleus to the cytosol in MCD-induced NASH (Supplementary Fig. 9). These data indicate that the decreased interaction between METTL3 and HDAC1/2 in NASH was likely due to the cytosolic translocation of METTL3. To determine whether METTL3 regulates the acetylation of histone H3K9 and H3K27 in the promoters of Cd36 and Ccl2, the acetylation of H3K9 and H3K27 in Mettl3-HOE, Mettl3-HKO, and corresponding control mice was measured by ChIP. As shown in Fig. 9e, f, both H3K9ac and H3K27ac levels in the promoters of Cd36 and Ccl2 were significantly decreased in the livers of Mettl3-HOE mice, which indicates that METTL3 was able to elicit deacetylation of H3K9 and H3K27 in the promoters of Cd36 and Ccl2. To confirm that the inhibition of Cd36 and Ccl2 expression is mediated by METTL3-elicited histone deacetylation, primary hepatocytes were infected with Ad-METTL3 and treated with or without trichostatin A (TSA), an HDACs inhibitor. As shown in Fig. 9g, h, TSA reversed the suppression of Cd36 and Ccl2 expression mediated by METTL3 overexpression. To further test this hypothesis, we used a more specific HDAC1/2 inhibitor (Romidepsin) to repeat this experiment. As shown in Supplementary Fig. 10a–b, Romidepsin completely reversed the suppression of Cd36 expression and partially rescued the inhibition of Ccl2 expression mediated by METTL3 overexpression. Furthermore, we measured the acetylation of H3K9 and H3K27 in the promoters of Cd36 and Ccl2 in the livers of Mettl3-HKO and Mettl3flox/flox mice. As shown in Fig. 9i, j, hepatic deletion of Mettl3 markedly increased both H3K9ac and H3K27ac levels in the Cd36 and Ccl2 promoters in the liver. To further determine whether METTL3 regulates HDAC1/2 activity, we measured the activities of HDAC1/2 in the livers of Mettl3-HKO and Mettl3flox/flox mice. As shown in Supplementary Fig. 11, the activities of HDAC1/2 were significantly decreased in Mettl3-HKO mice. These data demonstrate that METTL3 regulates Cd36 and Ccl2 transcription by modulating H3K9ac and H3K27ac in their promoters by the involvement of HDAC1/2.

a, b HA-HDAC1/2 expression vector was co-transfected with or without Flag-METTL3 expression vector in HEK293T cells. Total cell lysates were first incubated with DNase1 (200 U/ml) at 37 °C for 30 min. These lysates were immunoprecipitated with Flag or HA beads and then immunoblotted with anti-HA or anti-Flag antibodies. The samples were derived from the same experiment and the blots were processed in parallel. c Liver extracts from control and Mettl3-HOE mice at 8 weeks old were first incubated with DNase1 (200 U/ml) at 37 °C for 30 min. These lysates were immunoprecipitated with Flag beads and then immunoblotted with anti-HDAC1 or anti-HDAC2 antibodies. The samples were derived from the same experiment and the blots were processed in parallel. d Mettl3-HOE mice were fed an NC or MCD for 2 weeks. Liver extracts were first incubated with DNase1 (200 U/ml) at 37 °C for 30 min. These lysates were immunoprecipitated with Flag beads and then immunoblotted with anti-HDAC1 or anti-HDAC2 antibodies. The HDAC1/2 protein levels associated with Flag-METTL3 were quantified by ImageJ and normalized to input HDAC1/2 (n = 3 for each group; For HDAC1, NC versus MCD, P = 0.0096; For HDAC2, NC VS MCD, P = 0.0462). The samples were derived from the same experiment and the blots were processed in parallel. e, f H3K9ac and H3K27ac levels in the promoters of Ccl2, Cd36 and Actb genes in Mettl3-HOE and control mice fed an MCD for 2 weeks (n = 6 for each group; For H3K9ac, Ccl2, P = 0.0334, Cd36, P = 0.03, Actb, P = 0.8045; For H3K27ac, Ccl2, P = 0.0401, Cd36, P = 0.02585, Actb, P = 0.6527). g, h Primary hepatocytes were infected with Ad-βGal and Ad-METTL3 adenovirus. Ad-METTL3-infected hepatocytes were treated with or without TSA (2 μM) overnight. The relative Cd36 and Ccl2 mRNA levels were measured by qPCR (For Cd36, n = 6 for each group, βGal versus METTL3, P = 0.0016, METTL3 versus METTL3+TSA, P = 0.0009; For Ccl2, n = 4 for each group, βGal versus METTL3, P = 0.0054, METTL3 versus METTL3+TSA, P = 0.0167). CD36 and CCL2 protein levels were measured by immunoblotting, quantified by ImageJ and normalized to Tubulin (n = 5 for each group; For CD36, βGal versus METTL3, P = 0.0004, METTL3 versus METTL3+TSA, P = 0.0112; For CCL2, βGal versus METTL3, P < 0.0001, METTL3 versus METTL3+TSA, P = 0.0167). The samples were derived from the same experiment and the blots were processed in parallel. i, j H3K9ac and H3K27ac levels in the promoters of Ccl2, Cd36 and Actb genes in the livers of Mettl3-HKO and Mettl3flox/flox mice at 8 weeks old (n = 7 for each group; for H3K9ac, Ccl2, P = 0.0037, Cd36, P = 0.0113, Actb, P = 0.4987; For H3K27ac, Ccl2, P = 0.0097, Cd36, P = 0.0126, Actb, P = 0.4736). n was the number of biologically independent mice or cell samples. The cell culture experiments were repeated three times independently with similar results. Data represent the mean ± SEM. Significance was determined by unpaired two-tailed Student’s t test analysis. *P < 0.05. **P < 0.01. Source data are provided as a Source Data file.
Discussion
NASH is a major worldwide health problem and is characterized by hepatic steatosis, liver injury, and chronic inflammation. NASH is a key step whereby patients with simple NAFL develop cirrhosis and HCC3,5. However, only 25% of patients with NAFL develop NASH3. Thus, it is crucial to identify the positive and negative regulators that determine whether patients with simple NAFL develop NASH. In the present study, METTL3 was identified as a previously unrecognized suppressor of the NAFL-to-NASH transition. Hepatic deletion of Mettl3 promotes NASH progression under either HFD or MCD-feeding conditions due to increased hepatic free fatty acid uptake and inflammation, which are strongly associated with increased expression of Cd36 and Ccl2, respectively. Hepatic overexpression of Mettl3 protects against MCD-induced NASH by suppressing Cd36 and Ccl2 expression. Mechanistically, METTL3 directly binds to the promoters of Cd36 and Ccl2 genes and recruits HDAC1/2, which causes deacetylation of H3K9 and H3K27, thus suppressing Cd36 and Ccl2 transcription.
The loss of Mettl3 in the liver leads to more-pronounced steatosis, steatohepatitis, and collagen deposition, which suggests a robust fibrosis progression, whereas overexpression of Mettl3 in the liver induces the opposite effects. In agreement with these results, we showed that hepatic deletion of Mettl3 increases free fatty-acid uptake through increased Cd36 expression, whereas hepatic Mettl3 overexpression induces the opposite effects. It has been shown that abnormally increased expression of CD36 contributes to the development of steatosis and NASH12, whereas knockout of Cd36 has been shown to protect against MCD-induced NASH13. However, factors related to lipogenesis, fatty acid oxidation, and VLDL secretion were not changed by overexpression of Mettl3. Gene expression data suggest that fatty acid β-oxidation, lipogenesis, and VLDL secretion do not contribute to the increased liver steatosis seen in Mettl3-HKO mice. Thus, the loss of Mettl3 in the liver promotes hepatic steatosis mainly by increasing Cd36 expression.
Liver steatosis is considered a first but insufficient hit to induce NASH5. This is why an HFD alone cannot induce NASH. Surprisingly, an HFD is sufficient to induce NASH in Mettl3-HKO mice. Liver inflammation has been shown to be a key secondary hit for the NAFL-to-NASH transition. We confirmed the expression of cytokines and chemokines and found that CCL2 was strongly upregulated in Mettl3-HKO livers and hepatocytes. CCL2 and its receptor CCR2 are strongly associated with NASH14,15, and inhibition of CCL2 and CCR2 has been shown to be a therapeutic target for the treatment of NASH16,17. Hepatic overexpression of Mettl3 ameliorates MCD-induced NASH most likely by suppressing CCL2-induced inflammation and CD36-mediated free fatty acid uptake. Blockade of CD36 and CCL2 with neutralizing antibodies ameliorates the NASH progression in Mettl3-HKO mice, which further supports the theory that elevated CD36 and CCL2 contribute to the NASH progression in Mettl3-HKO mice.
Our focus was on the question of how METTL3 regulates Cd36 and Ccl2 expression. METTL3 is considered a key RNA methyltransferase that catalyzes RNA m6A modification and regulates most of the RNA processing steps18. m6ARIP-seq analysis identified 6,556 genes with m6A peaks in the livers of Mettl3flox/flox mice, and 3,489 genes showed decreased m6A levels in the livers of Mettl3-HKO mice, which indicates the importance of METTL3-mediated m6A modification in liver function. However, no significant m6A peaks were detected in either the Cd36 or the Ccl2 transcript in the livers of Mettl3flox/flox and Mettl3-HKO mice, which indicates that METTL3 regulates their expression through some means other than mRNA m6A modification. Instead, we observed that METTL3 binds directly to the promoters of Cd36 and Ccl2 and represses their promoter luciferase activities. METTL3 also interacts with HDAC1/2 in the liver. This interaction is likely disrupted during NASH progression. Hepatic deletion of Mettl3 decreases the activities of HDAC1/2, indicating that METTL3 regulates HDAC1/2 activity. Hepatic overexpression of Mettl3 decreases the levels of H3K9ac and H3K27ac, which are active epigenetic marks, in the promoters of Cd36 and Ccl2, leading to decreased transcription of Cd36 and Ccl2. Conversely, hepatic ablation of Mettl3 increases H3K9ac and H3K27ac levels in the promoters of Cd36 and Ccl2, which likely results in elevated chromatin accessibility and increased transcription of these genes. Furthermore, inhibition of HDAC1/2 by either TSA or Romidepsin blocks the suppression of Cd36 and Ccl2 transcription mediated by METTL3 overexpression. These data support a mechanism by which METTL3 suppresses free fatty acid uptake and inflammation at least in part by recruiting HDAC1/2 to the promoters of Cd36 and Ccl2 genes, where HDAC1/2 catalyzes the repressive deacetylation of H3K9 and H3K27. HDAC1/2 is not the only factor regulating H3K9ac/H3K27ac. Other HDACs and histone acetyltransferase (p300/CBP) may also contribute to the regulation of H3K9ac/H3K27ac. We cannot rule out the possibility that other HDACs and CBP/EP300 might be involved in this process, as Mettl14 deletion increases CBP/EP300 expression by increasing RNA stability due to decreased m6A modification29.
Several pieces of evidence support the conclusion that METTL3 can regulate the transcription of Cd36 and Ccl2 in the liver independent of its methyltransferase enzyme activity. Both METTL3 and METTL3 enzyme-dead mutation (D395A) reduces both Cd36 and Ccl2 promoter-luciferase activities in a dose-dependent manner. Consistently, both METTL3 and its mutation D395A significantly decrease the mRNA and protein levels of CD36 and CCL2. METTL3 mutation (D395A) also dramatically decreases the expression of Cd36 and Ccl2 in isolated hepatocytes from Mettl3-HKO mice. These data demonstrate that METTL3 serves as a transcriptional repressor of both the Cd36 and Ccl2 genes independent of its methyltransferase enzyme activity. Although METTL3-mediated m6A modification may not be involved in the regulation of Cd36 and Ccl2 expression in the pathogenesis of NASH, METTL3-mediated m6A modification may regulate the expression of other genes and other liver processes33, which requires further study. METTL14 and WTAP both interact with METTL318,19,20. Whether METTL14 and WTAP also negatively regulate NASH progression by inhibiting the expression of Cd36 and Ccl2 also needs future investigation.
Two recent studies show that METTL3 in mouse embryonic stem cells regulates chromatin accessibility depending on its methyltransferase enzyme activity47,48. One of the studies shows that Mettl3 knockout in mouse embryonic stem cells increases chromatin accessibility and activates transcription by increasing expression of the chromosome-associated regulatory RNAs, especially LINE1 elements, due to decreased m6A modification47. We checked the mRNA and m6A levels of LINE1 in the livers of Mettl3-HKO and Mettl3flox/flox mice from our RNA-seq and m6ARIP-seq data. RNA-seq data show that the fpkm value of LINE1(ENSMUSG00000087166) is o (zero) in the livers of Mettl3-HKO and Mettl3flox/flox mice, which indicates that LINE1 is not expressed in the liver, and unlikely regulates the chromatin state in the livers of Mettl3-HKO mice. These data also indicate that METTL3 can regulate chromatin accessibility through different mechanisms in a context-dependent and cell-type-specific manner.
Nuclear METTL3 protein levels are decreased in both human patients and mice with NASH, whereas nuclear METTL3 protein levels are increased in NAFL, which indicates that decreased nuclear METTL3 may contribute to the NAFL-to-NASH transition. Decreased nuclear METTL3 reduces HDAC1/2 activity in the promoters of Cd36 and Ccl2, which results in higher levels of H3K9ac and H3K27ac in their promoters and further increases their transcription. This has been observed in Mettl3-HKO mice. With respect to the nuclear/cytosolic translocation of METTL3 in NASH, TNFα/CDK9-mediated serine phosphorylation of METTL3 partially blocks the nuclear localization of METTL3, while inhibition of CDK9 increases the nuclear localization of METTL3. Although phosphorylation of some sites on METTL3 does not lead to nuclear/cytosolic translocation38, phosphorylation of other sites on METTL3 mediated by CDK9 may block its nuclear localization. We cannot rule out the possibility that other cytokines and kinases might also contribute to the cytosolic accumulation of METTL3 in NASH, as TNFa did not fully block its nuclear localization and a CDK9 inhibitor only partially blocked TNFa-induced cytosolic accumulation of METTL3.
In conclusion, we have demonstrated that METTL3 is an important repressor of NASH progression. Our data also reveal a mechanism by which METTL3 suppresses the transcription of Cd36 and Ccl2 via a histone modification pathway that includes the involvement of HDAC1/2. Decreased nuclear METTL3 may lead to impairment of the METTL3/HDAC1/2 axis, which further increases hepatic free fatty acid uptake and liver inflammation, contributing to NASH progression. Our results also indicate that the METTL3/HDAC1/2 axis may serve as a drug target for the treatment of NASH.
Methods
Animal experiments
Animal experiments were carried out in strict accordance with the Guide for the Care and Use of Laboratory Animals (8th edition). Animal experiment protocols were approved by the Institutional Animal Care and Use Committee of Harbin Institute of Technology (HIT/IACUC). The approval number was IACUC-2018004. Mice were housed on a 12-h light/12-h dark cycle, temperature (24 ± 2 °C) and humidity (50% ± 10%) conditions. Mice were fed a normal chow diet with free access to water. For diet-induced obesity, mice were fed an HFD (MD12032, 45% fat, Medicience) for 12 weeks. For diet-induced NASH, mice were fed an MCD (MD12052, Medicience) for 2–3 weeks. Mettl3flox/flox mice, in which exon 2 and exon 3 of the Mettl3 gene were flanked by two loxp sites, were generated using the CRISPR-Cas9 technique as previously reported27. STOP-Mettl3 mice were generated using the CRISPR-Cas9 technique to insert a STOP-Flag-Mettl3 cassette into the Rosa26 allele with the help of Shanghai Biomodel Organism. Hepatocyte-specific Mettl3 knockout (Mettl3-HKO) mice were generated by crossing Mettl3flox/flox mice with Alb-Cre mice. Hepatocyte-specific Mettl3-overexpressing mice were generated by crossing STOP-Mettl3+/− mice with Alb-Cre mice. For inhibition of CD36 and CCL2 in vivo, 8-week-old Mettl3-HKO mice were treated with a combination of anti-CD36 (ab23680, Abcam) and anti-CCL2 (MAB479500, R&D Systems) antibodies (4 μg/antibody/mouse, i.v.) or control antibodies (IgA, bs-0774P, Bioss, and IgG2b, 65211-1-Ig, Proteintech) on days 1, 4, 7, and 8 after MCD feeding. Mice were sacrificed on day 9 after MCD feeding. Blood samples were collected from the orbital sinus. The serum alanine aminotransferase (ALT) activities and TAG levels were measured with an ALT or TAG reagent set, respectively49.
Human liver samples
Human liver samples were collected in the Third Affiliated Hospital of Sun Yat-sen University. The present study was approved by the Research Ethics Committee of the Third Affiliated Hospital of Sun Yat-sen University, and individual permission was obtained using standard informed consent procedures. The investigation conformed to the principles outlined in the Declaration of Helsinki regarding the use of human tissues. Detailed characteristics of patients with or without NASH are listed in Supplementary Table 1.
Free fatty-acid uptake assays
Mettl3flox/flox and Mettl3-HKO mice were fasted overnight and then injected intraperitoneally with 20 μM BODIPY FL C16 in 200 μl saline for 20 min. Livers were harvested and homogenized in RIPA buffer. Liver lysates were mixed with three volumes of Dole’s reagent (heptane: 2-propanol: 2N sulfuric acid; 10:40:1) and centrifuged at 18,000 × g for 10 min. The BODIPY FL C16 fluorescence in the top organic-phase supernatant was determined using a 488 nm excitation and 515 nm emission filter set (BioTek) and normalized to the protein concentration50.
For the in vitro free fatty-acid uptake assay, primary hepatocytes were isolated from Mettl3flox/flox and Mettl3-HKO mice. Hepatocytes were incubated with BODIPY FL C16 (1 μM) for 30 min in phosphate-buffered saline (PBS), washed with PBS, and fixed with 4% paraformaldehyde (PFA) for 10 min at RT. BODIPY FL C16 fluorescence was observed using a fluorescent microscope (Olympus) and quantified using ImageJ version 1.39 f (National Institutes of Health).
Primary hepatocyte culture and adenoviral infection
Primary hepatocytes were isolated from C57BL/6 WT, Mettl3flox/flox, and Mettl3-HKO mice by liver perfusion with type II collagenase (Worthington Biochem, Lakewood, NJ) and cultured at 37 °C and 5% CO2 in DMEM medium supplemented with 5% FBS. Primary hepatocytes from C57BL/6 WT mice were infected with an equal amount of βGal, Flag-METTL3, or Flag-METTL3(D395A) adenoviruses overnight.
Nuclear run-on RT-qPCR
Nascent Cd36 and Ccl2 mRNA levels were measured by nuclear run-on RT-qPCR51,52. In brief, primary hepatocytes were isolated from Mettl3flox/flox and Mettl3-HKO mice. Nuclei were extracted in NP-40 lysis buffer (10 mM Tris HCl pH 7.4, 10 mM NaCl, 3 mM MgCl2, and 0.5% NP-40), and then incubated with BrUTP and unlabeled ribonucleotides in a transcription reaction buffer supplemented with 100 U RNase OUT, 0.5 mM BrUTP, 1 mM ATP, 1 mM GTP, 1 mM CTP, and 0.5 mM UTP at 30 °C for 30 min. Nuclear RNA was extracted using the TriPure Isolation Reagent (Roche, Mannheim, Germany). These nuclear RNA samples were immunoprecipitated with anti-BrdU antibody (66241-1-Ig, Proteintech). Nascent Cd36 and Ccl2 mRNA levels were then measured by RT-qPCR and normalized to 36B4. Primers for real-time RT-qPCR were listed in Supplementary Table 2.
Luciferase assays
The mouse Cd36 promoter (from −2001 to −1) and Ccl2 promoter (from −2001 to −1) were cloned into pGL3 vectors, respectively. Transient transfection and luciferase assays were performed49,52. Briefly, HEK293T cells were seeded in 24-well plates 20 h before transfection. Cd36 or Ccl2 luciferase reporter plasmids were co-transfected with METTL3, D395A, or empty expression vector by polyethylenimine (Sigma) into HEK293T cells. 48 h later, HEK293T cells were lysed in reporter lysis buffer (Promega, Madison, WI), and luciferase activity was measured and normalized to β-galactosidase (β-Gal) activity. The reagents were listed in Supplementary Table 3.
Transient transfection, immunoprecipitation, and immunoblotting
HEK293T cells were seeded in 6-well plates 16 h before transfection. The indicated expression vectors (Myc-CDK9, HA-HDAC1, or HA-HDAC2: 1 μg) were co-transfected with or without the Flag-METTL3 expression vector (1 μg) in HEK293T cells. For the Co-IP of HDAC1/2 and METTL3, total cell lysates were first incubated with DNase1 (200 U/ml) at 37 °C for 30 min to digest genomic DNA. These lysates were then immunoprecipitated with Myc, Flag, or HA beads at 4 °C for 2 h. Immunoblotting was performed using the indicated antibodies. Antibody dilutions were as follows: METTL3 (96391, Cell Signaling Technology), 1:2500; Lamin B1 (12987-1-AP, Proteintech), 1:5000; Tubulin (sc-5286, Santa Cruz), 1:5000; β-actin (60008-1-Ig, Proteintech), 1:5000; CDK9 (11705-1-AP, Proteintech), 1:2500; p-CDK9 (2549, Cell Signaling Technology), 1:2500; CD36 (18836-1-AP, Proteintech), 1:5000; Flag (F1804, Sigma), 1:5000; Myc (16286-1-AP, Proteintech), 1:5000; HDAC1 (5356, Cell Signaling Technology), 1:2500; HDAC2 (5113, Cell Signaling Technology), 1:2500; and Caspase3 (9662, Cell Signaling Technology), 1:2500. Other reagents were listed in Supplementary Table 3.
In vitro kinase assay
Myc-CDK9 was immunopurified using anti-Myc magnetic Beads (B26301, Bimake). Myc-CDK9 was incubated with purified Flag-METTL3 (1.5 μg) in kinase buffer (20 mM HEPES, pH 7.5-7.6, 33 μM ATP, 10 mM MgCl2, 50 mM NaCl, 1 mM PMSF and Phosphatase Inhibitor Cocktail 1) at 30 °C for 30 min. Flag-METTL3 (1.5 μg) in the kinase buffer without Myc-CDK9 was served as a control. The reactions were stopped by adding SDS-PAGE loading buffer and boiling for 5 min. Proteins were immunoblotted with antibodies against phosphoserine, Flag, and Myc, respectively. Antibody dilutions were as follows: phosphoserine (AB1603, Sigma), 1:1000; Flag (F1804, Sigma), 1:5000; and Myc (16286-1-AP, Proteintech), 1:5000.
Real-time quantitative PCR
Total RNA was extracted using the TriPure Isolation Reagent (Roche, Mannheim, Germany), and first-strand cDNA was synthesized using random primers and M-MLV reverse transcriptase (Promega, Madison, WI)53. RT-qPCR was performed using a Roche LightCycler 480 real-time PCR system (Roche, Mannheim, Germany)54. The data were analyzed using LightCycler 480 Software (v1.5.1). The expression of individual genes was normalized to the expression of 36B4. Primers for real-time RT-qPCR were listed in Supplementary Table 2.
Chromatin immunoprecipitation assays
The livers were fixed by liver perfusion with 1% PFA. The nuclei were isolated from livers and subjected to sonication (M220, Focused-ultrasonicator; Covaris) to break genomic DNA into 500- to 1000-bp fragments using a chromatin shearing kit (520127, truChIP Chromatin Shearing Kit, Covaris). The samples were immunoprecipitated with anti-FLAG M2 magnetic beads (M8823, Millipore) or antibodies against H3K9ac or H3K27ac. DNA was extracted and used for qPCR analysis. Primers for qPCR were listed in Supplementary Table 2.
Nuclear extract preparation
Liver tissues were homogenized in lysis buffer (20 mM HEPES, 1 mM EDTA, 250 mM sucrose, 1 mM PMSF, 1 mM Na3VO3 and 0.5 mM DTT, pH 7.4) and centrifuged sequentially at 1100 × g and 4000 × g at 4 °C. Nuclear protein was extracted from the pellets using a high-salt solution (20 mM HEPES, 420 mM NaCl, 0.2 mM EDTA, 0.5 mM DTT, 1 mM PMSF and 1 mM Na3VO3, pH 7.9). The preparation of nuclear and cytosolic proteins from primary hepatocytes was performed using a commercial Nuclear and Cytoplasmic Protein Extraction Kit (P0027, Beyotime).
RNA-sequencing and m6ARIP-sequencing
For RNA-sequencing, total RNA was extracted using Tripure Isolation Reagent (94015120, Roche, Mannheim, Germany) from livers of Mettl3flox/flox and Mettl3-HKO mice at 8 weeks old. Each sample was pooled from four mice for each group. A total amount of 1 μg RNA per sample was used for the RNA sample preparations. Sequencing libraries were generated using NEBNext® Ultra™ RNA Library Prep Kit for Illumina® (NEB, USA) following the manufacturer’s recommendations and index codes were added to attribute sequences to each sample. The clustering of the index-coded samples was performed on a cBot Cluster Generation System using TruSeq PE Cluster Kit v3-cBot-HS (Illumia) according to the manufacturer’s instructions. After cluster generation, the library preparations were sequenced on an Illumina Hiseq X Ten platform and 125 bp/150 bp paired-end reads were generated. Paired-end clean reads were aligned to the mouse reference genome (Ensemble_GRCm38.90) with Hisat2 (version 2.0.4), and the aligned reads were used to quantify mRNA expression by using HTSeq (version 0.9.1).
For m6ARIP-sequencing, total RNA was extracted using Tripure Isolation Reagent (94015120, Roche, Mannheim, Germany) from livers of Mettl3flox/flox and Mettl3-HKO mice at 8 weeks old. Each sample (300 μg total RNA) was pooled from three mice for each group. Poly(A)+RNA was purified using Dynabeads™ mRNA Purification Kit (61006, Invitrogen). Fragmented poly(A)+RNA was incubated with m6A antibody (202003, Synaptic System) for immunoprecipitation using Magna MeRIPTM m6A Kit (17-10499, MERCK). Then, immunoprecipitated mRNA or Input was used for library construction with NEBNext ultra RNA library prep kit for Illumina (New England Biolabs). The library preparations were sequenced on an Illumina Hiseq X platform. Sequenced reads were then mapped to Ensemble_GRCm38.90 whole genome using BWA (V0.7.12).
After mapping reads to the reference genome, exomePeak R package (v2.16.0) was used for the m6A peak identification in each anti-m6A immunoprecipitation group with the corresponding input samples serving as a control, and q value threshold of enrichment of 0.05 was used for all data sets. The m6A-enriched motifs of each group were identified by HOMER (v4.9.1)55. Peak-related genes were confirmed by PeakAnnotator (v7.4), and then GO enrichment analysis of performed to identify the function enrichment results. GO enrichment analysis was implemented by the GOseq R package (v1.26.0), in which gene length bias was corrected. GO terms with corrected P value < 0.05 were considered significantly enriched by peak-related genes.
ATAC-sequencing
ATAC-seq was performed using a standard protocol with some modifications56,57. Liver samples were pooled from the livers of Mettl3-HKO and Mettl3flox/flox mice (n = 4 for each group). Nuclei were extracted from liver samples, and the nuclei pellets were resuspended in the Tn5 transposase reaction mix. The transposition reaction was incubated at 37 °C for 30 min. Equimolar adapter1 and adapter2 were added after transposition, and PCR was then performed to amplify the library. After PCR, libraries were purified with the AMPure beads, and library quality was assessed with Qubit. Clustering of the index-coded samples was performed on a cBot Cluster Generation System using TruSeq PE Cluster Kit v3-cBot-HS (Illumina). After cluster generation, the library preparations were sequenced on an Illumina NovaSeq 6000 platform and 150 bp paired-end reads were generated. Nextera adaptor sequences were firstly trimmed from the reads using skewer (V0.2.2). These reads were aligned to a reference genome using BWA (V0.7.12), with standard parameters. These reads were then filtered for high quality (MAPQ 13), non-mitochondrial chromosome, and properly paired reads (longer than 18 nt). All peak calling was performed with macs2 using “macs2 callpeak —nomodel —keepdup all —call-summits”. For simulations of peaks called per input read, aligned and de-duplicated BAM files were used without any additional filtering.
HDAC1 and HDAC2 activity assays
Liver tissues from Mettl3flox/flox and Mettl3-HKO mice were homogenized in an L-RIPA lysis buffer (50 mM Tris, pH 7.5, 1% Nonidet P-40, 150 mM NaCl, 2 mM EGTA, 1 mM Na3VO4, 100 mM NaF, 10 mMNa4P2O7, 1 mM phenylmethylsulfonyl fluoride). HDAC1 or HDAC2 was immunoprecipitated from 200 μg of total lysate with 0.9 μg HDAC1 (10197-1-AP, Proteintech) or HDAC2 (12922-3-AP, Proteintech) antibody. As a negative control, we used 0.9 μg IgG antibody (2729, Cell Signaling Technology) to detect non-specific-bound HDAC activity. Immunoprecipitates were then assayed for HDAC activity using an Epigenase HDAC Activity/Inhibition Direct Assay Kit (Epigentek) according to the manufacturer’s protocol. HDAC1/2 activity values were corrected for non-specific (IgG)-bound HDAC activity and divided by the average HDAC1/2 activity value in Mettl3flox/flox mouse livers to determine the relative HDAC1/2 activity.
Statistical analysis
Data were analyzed using GraphPad Prism 6.02. Data were presented as means ± SEM. Differences between groups were analyzed by unpaired two-tailed Student’s t tests. P < 0.05 was considered statistically significant. *P < 0.05. **P < 0.01.
Reporting summary
Further information on research design is available in the Nature Research Reporting Summary linked to this article.
Data availability
The ATAC-seq and RNA-seq data generated in this study have been deposited in the GEO database under accession code GSE141325. The m6ARIP-seq data generated in this study have been deposited in the GEO database under accession code GSE142835. The m6ARIP-seq data generated in this study are provided in the Supplementary Information/Source Data file. The published RNA-seq data used in this study are available in the GEO database under accession codes GSE43314 and GSE11934035,36,37. All other data generated or analyzed during this study are included in this published article (and its supplementary information files). Source data are provided with this paper.
References
Younossi, Z. et al. Global burden of NAFLD and NASH: trends, predictions, risk factors and prevention. Nat. Rev. Gastroenterol. Hepatol. 15, 11–20 (2018).
Younossi, Z. M. et al. Global epidemiology of nonalcoholic fatty liver disease-Meta-analytic assessment of prevalence, incidence, and outcomes. Hepatology 64, 73–84 (2016).
Diehl, A. M. & Day, C. Cause, pathogenesis, and treatment of nonalcoholic steatohepatitis. N. Engl. J. Med. 377, 2063–2072 (2017).
Anstee, Q. M., Reeves, H. L., Kotsiliti, E., Govaere, O. & Heikenwalder, M. From NASH to HCC: current concepts and future challenges. Nat. Rev. Gastroenterol. Hepatol. 16, 411–428 (2019).
Machado, M. V. & Diehl, A. M. Pathogenesis of nonalcoholic steatohepatitis. Gastroenterology 150, 1769–1777 (2016).
Kazankov, K. et al. The role of macrophages in nonalcoholic fatty liver disease and nonalcoholic steatohepatitis. Nat. Rev. Gastroenterol. Hepatol. 16, 145–159 (2019).
Day, C. P. & James, O. F. Steatohepatitis: a tale of two “hits”? Gastroenterology 114, 842–845 (1998).
Friedman, S. L., Neuschwander-Tetri, B. A., Rinella, M. & Sanyal, A. J. Mechanisms of NAFLD development and therapeutic strategies. Nat. Med. 24, 908–922 (2018).
Donnelly, K. L. et al. Sources of fatty acids stored in liver and secreted via lipoproteins in patients with nonalcoholic fatty liver disease. J. Clin. Invest. 115, 1343–1351 (2005).
Schuster, S., Cabrera, D., Arrese, M. & Feldstein, A. E. Triggering and resolution of inflammation in NASH. Nat. Rev. Gastroenterol. Hepatol. 15, 349–364 (2018).
Sanyal, A. J. Past, present and future perspectives in nonalcoholic fatty liver disease. Nat. Rev. Gastroenterol. Hepatol. 16, 377–386 (2019).
Miquilena-Colina, M. E. et al. Hepatic fatty acid translocase CD36 upregulation is associated with insulin resistance, hyperinsulinaemia and increased steatosis in non-alcoholic steatohepatitis and chronic hepatitis C. Gut 60, 1394–1402 (2011).
Bieghs, V. et al. Role of scavenger receptor A and CD36 in diet-induced nonalcoholic steatohepatitis in hyperlipidemic mice. Gastroenterology 138, 2477–2486.e2473 (2010).
Haukeland, J. W. et al. Systemic inflammation in nonalcoholic fatty liver disease is characterized by elevated levels of CCL2. J. Hepatol. 44, 1167–1174 (2006).
Miura, K., Yang, L., Rooijen, N. V., Ohnishi, H. & Seki, E. Hepatic recruitment of macrophages promotes nonalcoholic steatohepatitis through CCR2. Am. J. Physiol. Gastrointest. Liver Physiol. 302, G1310–G1321 (2012).
Baeck, C. et al. Pharmacological inhibition of the chemokine CCL2 (MCP-1) diminishes liver macrophage infiltration and steatohepatitis in chronic hepatic injury. Gut 61, 416–426 (2012).
Lefebvre, E. et al. Antifibrotic effects of the dual CCR2/CCR5 antagonist cenicriviroc in animal models of liver and kidney fibrosis. PLoS One 11, e0158156 (2016).
Liu, J. et al. A METTL3-METTL14 complex mediates mammalian nuclear RNA N6-adenosine methylation. Nat. Chem. Biol. 10, 93–95 (2014).
Ping, X. L. et al. Mammalian WTAP is a regulatory subunit of the RNA N6-methyladenosine methyltransferase. Cell Res. 24, 177–189 (2014).
Frye, M., Harada, B. T., Behm, M. & He, C. RNA modifications modulate gene expression during development. Science 361, 1346–1349 (2018).
Yoon, K. J. et al. Temporal control of mammalian cortical neurogenesis by m(6)A methylation. Cell 171, 877–889 e817 (2017).
Wang, C. X. et al. METTL3-mediated m6A modification is required for cerebellar development. PLoS Biol. 16, e2004880 (2018).
Lin, Z. et al. Mettl3-/Mettl14-mediated mRNA N(6)-methyladenosine modulates murine spermatogenesis. Cell Res. 27, 1216–1230 (2017).
Fustin, J. M. et al. RNA-methylation-dependent RNA processing controls the speed of the circadian clock. Cell 155, 793–806 (2013).
Batista, P. J. et al. m(6)A RNA modification controls cell fate transition in mammalian embryonic stem cells. Cell Stem Cell 15, 707–719 (2014).
Lin, S. & Gregory, R. I. Methyltransferases modulate RNA stability in embryonic stem cells. Nat. Cell Biol. 16, 129–131 (2014).
Wang, Y. et al. METTL3 is essential for postnatal development of brown adipose tissue and energy expenditure in mice. Nat. Commun. 11, 1648 (2020).
Li, X., Jiang, Y., Sun, X., Wu, Y. & Chen, Z. METTL3 is required for maintaining β-cell function. Metabolism 116, 154702 (2021).
Wang, Y. et al. N(6)-methyladenosine RNA modification regulates embryonic neural stem cell self-renewal through histone modifications. Nat. Neurosci. 21, 195–206 (2018).
Barbieri, I. et al. Promoter-bound METTL3 maintains myeloid leukaemia by m(6)A-dependent translation control. Nature 552, 126–131 (2017).
Choe, J. et al. mRNA circularization by METTL3-eIF3h enhances translation and promotes oncogenesis. Nature 561, 556–560 (2018).
Liu, J. et al. m(6)A mRNA methylation regulates AKT activity to promote the proliferation and tumorigenicity of endometrial cancer. Nat. Cell Biol. 20, 1074–1083 (2018).
Lu, J. et al. Mechanisms of RNA N(6)-methyladenosine in hepatocellular carcinoma: from the perspectives of etiology. Front. Oncol. 10, 1105 (2020).
Zhong, L. et al. METTL3 induces AAA development and progression by modulating N6-methyladenosine-dependent primary miR34a [rocessing. Mol. Ther. Nucleic Acids 21, 394–411 (2020).
Zhang, F. et al. Gene expression profile analysis of type 2 diabetic mouse liver. PLoS ONE 8, e57766 (2013).
Xiong, X. et al. Landscape of intercellular crosstalk in healthy and NASH liver revealed by single-cell secretome gene analysis. Mol. Cell 75, 644–660 e645 (2019).
Xiong, X. et al. Mapping the molecular signatures of diet-induced NASH and its regulation by the hepatokine Tsukushi. Mol. Metab. 20, 128–137 (2019).
Scholler, E. et al. Interactions, localization, and phosphorylation of the m(6)A generating METTL3-METTL14-WTAP complex. RNA 24, 499–512 (2018).
Xie, W., Ma, L. L., Xu, Y. Q., Wang, B. H. & Li, S. M. METTL3 inhibits hepatic insulin sensitivity via N6-methyladenosine modification of Fasn mRNA and promoting fatty acid metabolism. Biochem. Biophys. Res. Commun. 518, 120–126 (2019).
Ricchi, M. et al. Differential effect of oleic and palmitic acid on lipid accumulation and apoptosis in cultured hepatocytes. J. Gastroenterol. Hepatol. 24, 830–840 (2009).
Kudo, H. et al. Lipopolysaccharide triggered TNF-α-induced hepatocyte apoptosis in a murine non-alcoholic steatohepatitis model. J. Hepatol. 51, 168–175 (2009).
Bauer, N. C., Doetsch, P. W. & Corbett, A. H. Mechanisms regulating protein localization. Traffic 16, 1039–1061 (2015).
Youn, J. H. & Shin, J. S. Nucleocytoplasmic shuttling of HMGB1 is regulated by phosphorylation that redirects it toward secretion. J. Immunol. 177, 7889–7897 (2006).
Gaudet, P. et al. The neXtProt knowledgebase on human proteins: 2017 update. Nucleic Acids Res. 45, D177–D182 (2017).
Orchard, S. et al. The MIntAct project—IntAct as a common curation platform for 11 molecular interaction databases. Nucleic Acids Res. 42, D358–D363 (2014).
Kawano, Y. & Cohen, D. E. Mechanisms of hepatic triglyceride accumulation in non-alcoholic fatty liver disease. J. Gastroenterol. 48, 434–441 (2013).
Liu, J. et al. N (6)-methyladenosine of chromosome-associated regulatory RNA regulates chromatin state and transcription. Science 367, 580–586 (2020).
Xu, W. et al. METTL3 regulates heterochromatin in mouse embryonic stem cells. Nature 591, 317–321 (2021).
Ren, X. et al. A small-molecule inhibitor of NF-κB-inducing kinase (NIK) protects liver from toxin-induced inflammation, oxidative stress, and injury. FASEB J. 31, 711–718 (2017).
Demers, A. et al. PCSK9 induces CD36 degradation and affects long-chain fatty acid uptake and triglyceride metabolism in adipocytes and in mouse liver. Arterioscler. Thromb. Vasc. Biol. 35, 2517–2525 (2015).
Roberts, T. C. et al. Quantification of nascent transcription by bromouridine immunocapture nuclear run-on RT-qPCR. Nat. Protoc. 10, 1198–1211 (2015).
Jia, L., Jiang, Y., Li, X. & Chen, Z. Purbeta promotes hepatic glucose production by increasing Adcy6 transcription. Mol. Metab. 31, 85–97 (2020).
Li, X. et al. Activation of NF-κB-inducing kinase in islet β cells causes β cell failure and diabetes. Mol. Ther. J. Am. Soc. Gene Ther. 28, 2430–2441 (2020).
Qin, W. et al. A long non-coding RNA, APOA4-AS, regulates APOA4 expression depending on HuR in mice. Nucleic Acids Res. 44, 6423–6433 (2016).
Dominissini, D., Moshitch-Moshkovitz, S., Salmon-Divon, M., Amariglio, N. & Rechavi, G. Transcriptome-wide mapping of N(6)-methyladenosine by m(6)A-seq based on immunocapturing and massively parallel sequencing. Nat. Protoc. 8, 176–189 (2013).
Buenrostro, J. D., Giresi, P. G., Zaba, L. C., Chang, H. Y. & Greenleaf, W. J. Transposition of native chromatin for fast and sensitive epigenomic profiling of open chromatin, DNA-binding proteins and nucleosome position. Nat. Methods 10, 1213–1218 (2013).
Corces, M. R. et al. An improved ATAC-seq protocol reduces background and enables interrogation of frozen tissues. Nat. Methods 14, 959–962 (2017).
Acknowledgements
This study was supported by the National Natural Science Foundation of China Grant (92057110 and 31971083) and the Natural Science Foundation of Heilongjiang Province (YQ2019C011). X.L. is supported by the Chinese Postdoctoral Science Foundation (AUGA4130900619). We thank Novogene for their assistance in ATAC-seq, RNA-seq, and M6ARIP-seq experiments.
Author information
Authors and Affiliations
Contributions
X.L. and B.Y. performed most of the experiments. M.L. initiated this project and researched data. Y.W., N.D., C.L. and M.G. researched data. Z.Y. and L.X. collected human liver samples and clinic data. S.Z. and Y.Z. provided research materials. Z.C. conceived and designed the project, researched data, and wrote the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review information
Nature Communications thanks the anonymous reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Li, X., Yuan, B., Lu, M. et al. The methyltransferase METTL3 negatively regulates nonalcoholic steatohepatitis (NASH) progression. Nat Commun 12, 7213 (2021). https://doi.org/10.1038/s41467-021-27539-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-021-27539-3
This article is cited by
-
Methyltransferase-like proteins in cancer biology and potential therapeutic targeting
Journal of Hematology & Oncology (2023)
-
The malignancy suppression and ferroptosis facilitation of BCL6 in gastric cancer mediated by FZD7 repression are strengthened by RNF180/RhoC pathway
Cell & Bioscience (2023)
-
Writing and erasing ceramides to alter liver disease
Nature Metabolism (2023)
-
m6A modification-tuned sphingolipid metabolism regulates postnatal liver development in male mice
Nature Metabolism (2023)
-
Deficiency of WTAP in islet beta cells results in beta cell failure and diabetes in mice
Diabetologia (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
