
Abstract
X-linked dystonia-parkinsonism is a neurodegenerative disorder caused by a founder retrotransposon insertion, in which a polymorphic hexanucleotide repeat accounts for ~50% of age at onset variability. Employing a genome-wide association study to identify additional factors modifying age at onset, we establish that three independent loci are significantly associated with age at onset (p < 5 × 10−8). The lead single nucleotide polymorphisms collectively account for 25.6% of the remaining variance not explained by the hexanucleotide repeat and 13.0% of the overall variance in age at onset in X-linked dystonia-parkinsonism with the protective alleles delaying disease onset by seven years. These regions harbor or lie adjacent to MSH3 and PMS2, the genes that were recently implicated in modifying age at onset in Huntington’s disease, likely through a common pathway influencing repeat instability. Our work indicates the existence of three modifiers of age at onset in X-linked dystonia-parkinsonism that likely affect the DNA mismatch repair pathway.
Similar content being viewed by others
Introduction
While recent years have seen unparalleled advances in identifying disease genes for monogenic neurogenetic conditions, a mostly unresolved challenge of personalized medicine remains the question as to whether a mutation carrier will manifest the disease and, if so, when and to what extent. Although elucidating the factors that contribute to this variability in (age-related) penetrance and disease expressivity has a high imperative and translational potential, this is typically impeded by the occurrence of private mutations with different effects, variable genetic and environmental background of patients, and small sample sizes. In contrast, X-linked dystonia-parkinsonism (XDP, DYT/PARK-TAF1, OMIM #314250) is a rare disease where patients share a founder mutation and come from the same locale (Panay Island, Philippines). An active group of local clinicians and researchers have made it possible to accumulate relatively large sample sizes for a disease with a global prevalence of <1:1,000,000, making XDP an ideal model to study genetic modifiers of disease expression1.
XDP is an adult-onset movement disorder characterized by striatal neurodegeneration and inherited in an X-linked recessive manner, thus predominantly affecting men. All patients carry the same disease-causing mutation, a ~2.6-kb SINE-VNTR-Alu (SVA)-type retrotransposon insertion in intron 32 of the TAF1 gene2,3. The XDP phenotype, already characterized by the presence of two different movement disorders (dystonia and parkinsonism), shows extraordinary variability in age and site of disease onset, as well as in the type of initial signs and rate of progression. Recently, we showed that the length of a polymorphic (CCCTCT)n repeat within this SVA retrotransposon insertion correlates inversely with age at onset (AAO)4,5 and TAF1 expression, as well as positively with disease severity and cognitive dysfunction4. The number of hexanucleotide repeats, however, accounts for ~50% of the AAO variability in the XDP patient population4,5 and even less of the variable expressivity of XDP4, suggesting additional modifiers.
In this work, we employ a genome-wide association study (GWAS) approach to search for additional factors influencing age-related penetrance of XDP. Our findings (i) confirm that genome-wide significant genetic modifiers can be identified in small but homogeneous samples of patients with monogenic disease; (ii) reveal a biologically plausible mechanism of disease modification in XDP; and (iii) pathophysiologically link XDP to Huntington’s disease (HD, OMIM #143100), another repeat expansion disorder with severe striatal neurodegeneration as an overlapping feature of both diseases.
Results
Genome-wide association study
Using DNA from 353 genetically confirmed XDP patients (Supplementary Table 1), we performed a GWAS in search of genetic modifiers of AAO in XDP. The average AAO (±standard deviation (SD)) of our patient cohort was 41.8 ± 8.4 (range: 21–67) years. The mean (±SD) detected (CCCTCT)n repeat number was 41.6 ± 4.1 (range: 30–55). Repeat numbers showed an inverse correlation with AAO (Pearson product-moment correlation: r = −0.703, p < 2.2 × 10−16), explaining 49.3% of the AAO difference in our 353 XDP patients (R2 = 0.493). Association of AAO with repeat number was previously investigated in 328 of these 353 patients4,5, and their combined analysis yielded an R2 of 0.488.
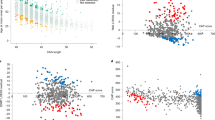
In the screening step, we identified 93 candidate single-nucleotide polymorphisms (SNPs) to cover potential genetic modifiers influencing AAO (Fig. 1a, Supplementary Table 2). Of those, 60 SNPs are located within a 0.33 Mb region on chromosome 5 (rs173441-rs10075662; GRCh37 coordinates: 79893519-80222980) (Fig. 1b), and 33 SNPs are situated on chromosome 7 within a 0.04 Mb area (rs17136307-rs660547; GRCh37 coordinates: 6064799-6106731) (Fig. 1c).

a Manhattan plot of the XDP AAO GWAS. We estimated a series of linear regression models assuming SNPs with a p-value below 5 × 10−8 to be significantly associated with AAO adjusted for repeat number. Using this approach, we identified 93 candidate SNPs to cover a potential genetic modifier influencing AAO. b, c Locus zoom plot of the GWAS data. The structure of linkage disequilibrium and –log10 (p-value) of the identified loci on chromosomes 5 and 7, respectively. Genomic coordinates are given according to GRCh37. Red lines: p = 5 × 10−8; Mb: megabase; each dot: −log10 (p-value) of a SNP; r2: the Pearson coefficient of correlation between any SNP in the graph and the SNP with the lowest p-value in that region. Source data are provided as a Source data file.
Out of the 60 SNPs within the candidate region on chromosome 5, 34 variants are located in the MSH3 gene (Fig. 1b). A further 26 SNPs are located in the DHFR gene (n = 4) and in the intergenic regions 5′ from DHFR and 3′ from MSH3 (n = 22) (Supplementary Table 2). All of the SNPs in DHFR and 33 of 34 MSH3 variants are non-coding (intronic). The only coding MSH3 SNP, rs1650697, p.Ile79Val, is in exon 1. The SNP with the most significant association with AAO in our GWAS (p = 1.1 × 10−12) is rs245013 located in intron 12 of MSH3 (ENST00000265081) and conferring an estimated mean decrease in AAO of 3.30 years per alternative allele (Fig. 2). Iterative conditional analyses revealed that some of the SNPs on chromosome 5 continue to be significantly associated with AAO, even if the regression was adjusted for this variant (rs245013) (Supplementary Fig. 1), indicating a second independently associated region within the chromosome 5 locus. This signal includes 6 variants (with p < 5.0 × 10−4 after adjusting for rs245013) situated in a 0.06 Mb area spanning intron 20 of MSH3 to intergenic region 3′ from MSH3. Among the 6 variants, rs33003 in MSH3 intron 23 has the lowest p-value (p = 7.5 × 10−5) after adjustment and confers an estimated mean decrease in AAO of 3.19 years per alternative allele (Fig. 2).

a, b Two independent putative genetic loci on chromosome 5, the alternative alleles of which were found to accelerate disease onset. c The alternative allele of the chromosome 7 signal delays AAO in our patients. Box-plot elements: center line: median; box limits: upper and lower quartiles; whiskers: 1.5× interquartile range; single points: outliers. Source data are provided as a Source data file.
Of the 33 SNPs within the associated region on chromosome 7, 22 are situated in the EIF2AK1 and ANKRD61 genes. Intron 11 of EIF2AK1 harbors rs62456190 (ENST00000199389) (Fig. 1c), the variant with the lowest p-value (1.3 × 10−9), that confers an estimated mean increase in AAO of 3.82 years per alternative allele (Fig. 2). The remaining 11 variants are situated within the intergenic region 5′ from EIF2AK1. Only two SNPs on chromosome 7 are exonic variants (rs4724769, p.Lys18= in exon 1 and rs2302334, p.Met355Leu in exon 3, ENST00000409061), both in ANKRD61. Conditional analyses indicated a single significant region on chromosome 7, as no other SNP remained significant after adjustment for rs62456190 (data not shown). Of note, the genes on chromosome 7 closest to the region associated with AAO in XDP are AIMP2 and PMS2, positioned 1.3/7.1 kb and 21.8/16.0 kb from the associated region/lead SNP (rs62456190), respectively.
When aggregating the p-values of single SNPs per gene, only MSH3 was significantly associated with AAO (p = 1.72 × 10−12). We did not observe any genetic pathway to be significantly associated with AAO at the stringent significance threshold (data not shown).
In summary, the lead SNPs resulting from our GWAS and conditional analyses suggest the existence of three potential genetic modifiers of AAO in XDP, two within or immediately adjacent to MSH3 and DHFR, and one within or immediately adjacent to ANKRD61, EIF2AK1, and PMS2 (Fig. 1). The three most significant SNPs (one from each of the three regions) collectively account for 25.6% of the remaining variance (not explained by the hexanucleotide repeat) and 13.0% of the overall variance in AAO in XDP. Since the SVA repeat number is also a genetic factor, the variance explained by the combination of repeat number and the three independent loci can be interpreted as a heritability, leading to an overall heritability of 63% (where the repeat number accounts for 79% of the heritability and the three independent loci for the remaining 21%). Patients carrying the harmful or protective genotypes at all three SNPs are expected to have an AAO 7.9 years earlier or 7.0 years later, respectively, than predicted by the number of XDP-relevant hexanucleotide repeats alone.
Analyses of variants in MSH3 exon 1
Using Sanger sequencing, we validated genotyping of the missense SNP closest to the strongest signal in our GWAS (MSH3 exon 1: rs1650697, c.235A>G, p.Ile79Val) in a subset of 285 XDP patients (Supplementary Table 1) with a validation rate of over 99%.
Sequence analysis of MSH3 exon 1 revealed ten single-nucleotide and in-frame sequence length polymorphisms in close proximity of p.Ile79Val (Supplementary Table 3). We sequenced exon 1 of MSH3 in 95 additional XDP patients whose mean AAO and hexanucleotide repeat number did not differ from the GWAS cohort (Supplementary Table 3). After adjustment for SVA hexanucleotide repeat number, we calculated the impact on AAO of eight variants (including p.Ile79Val) in MSH3 exon 1 with a MAF of >0.01 in 380 XDP patients in whom this exon was sequenced. Seven of these variants were associated with AAO with different directions of effect (Supplementary Table 3). Of note, the absence or presence of three in-frame sequence length polymorphisms [c.162_179del (p.Ala57_Ala62del), c.199_207del (p.Pro67_Pro69del), and c.181_189dup (p.Ala61_Pro63dup) the first two of which were always detected together; Supplementary Table 3] form alleles of three different sizes: (i) wild-type MSH3, (ii) a 27-nucleotide/9-amino-acid shorter, and (iii) a nine-nucleotide/3-amino-acid longer form previously described as 6a, 3a, and 7a, respectively (Fig. 3a)6.

a cDNA and amino-acid sequences showing the wild-type sequence, the shortest (c.162_179del;199_207del), and the longest (c.181_189dup) allele of MSH3 exon 1, i.e., 6a, 3a, and 7a alleles, respectively. The sequences are based on the MSH3 transcript ID ENST00000265081.7 (https://www.ensembl.org). Blue shading highlights the region deleted in the shortest allele (stretch of six alanines (A) and three other amino acids (proline, alanine, proline—PAP)). Red shading indicates the region duplicated in the longest allele. M: methionine (start codon); S: serine, R: arginine, AFPPQLPPHI: amino-acid sequence from the length polymorphism to the end of MSH3 exon 1. b Difference between the observed and predicted (based on the hexanucleotide repeat number) AAO negatively correlates with the number of 7a alleles in a genotype. Box-plot elements: center line: median; box limits: upper and lower quartiles; whiskers: 1.5× interquartile range; points: outliers. c Disease onset is delayed by ~4 years in heterozygous carriers of the shortest allele (3a/6a) in comparison to the patients homozygous for the longest MSH3 allele (7a/7a). Red color designates a harmful effect (earlier AAO), while blue depicts a protective effect (later AAO). The effect estimates are based on the linear regression model. Source data are provided as a Source data file.
Interestingly, we found that the lengths of these three variants inversely correlate with AAO in XDP. When considering the residual AAO variability (difference between the observed AAO and the AAO expected based on the hexanucleotide repeat number), patients carrying the 7a allele showed the most pronounced mean reduction in AAO, while patients heterozygous for the 3a allele had the highest mean increase in AAO (Fig. 3b and c). Of note, in our cohort, only two patients were homozygous for the shortest (3a) allele, and their AAOs were considerably higher than predicted based on the hexanucleotide repeat number (expected AAO of 38 and 46 years vs. observed AAO of 55 and 64 years).
In addition to testing single variant effects, we also assessed the association between the six haplotypes we identified in our cohort (with a frequency of >1%) and AAO (Supplementary Table 4). The four most common haplotypes showed a strong association with AAO, with H1 and H3 having an increasing (+1.70 and +3.21 years, respectively) and H2 and H4 a decreasing (−1.94 and −3.70 years, respectively) effect on AAO.
Effect of Huntington’s disease polygenic risk score on age at onset in XDP
We constructed a polygenic risk score (PRS) according to Gusella7 to predict the AAO in XDP and we observed an increase of 1.30 years (p = 1.06 × 10−9) for every increase of one in the PRS. In total, the PRS explained about 10% of the remaining variance in AAO, while the three lead SNPs of our GWAS explained more then 25% of the remaining variance. Overall, the PRS for HD was constructed based on a GWAS performed in observations from a different genetic population. Thus, the effect estimates are not necessarily transferable to a Filipino population.
Expression quantitative trait locus query and MSH3 and PMS2 expression analysis
Our query of the Genotype-Tissue Expression (GTEx) Portal8 revealed that rs245013 is a significant expression quantitative trait locus (eQTL) for MSH3 in multiple brain tissues including cortex (normalized effect size (NES) 0.32, p = 3.6 × 10−6), amygdala (NES: 047, p = 6.0 × 10−6), nucleus accumbens (basal ganglia) (NES: 0.29, p = 1.3 × 10−8), and putamen (NES: 3.1, p = 3.1 × 10−6) (Supplementary Data 1, Supplementary Fig. 2). The strongest signal within the second locus on chromosome 5 (rs33003) was not found in the GTEx Portal. The only coding MSH3 SNP, rs1650697, is a significant single-tissue eQTL for MSH3 in the cortex (NES: −0.45, p = 2.3 × 10−7) and nucleus accumbens (NES −0.34, p = 2.3 × 10−8) (Supplementary Fig. 2), and whole blood (NES −0.58, p = 4.5 × 10−63) (Supplementary Data 1). The lead SNP on chromosome 7 (rs62456190) is a significant eQTL for PMS2 in coronary artery (NES: 0.42 p = 7.1 × 10−9), pancreas (NES: 0.40, p = 8.4 × 10−11), pituitary (NES: 0.36, p = 3.3 × 10−7, etc. (top associations based on NES, Supplementary Data 1).
Queries performed using the BrainSeq dataset9 from the eQTL Catalog10 as well as the United Kingdom Brain Expression Consortium (UKBEC) dataset did not identify significant eQTLs for MSH3 (Supplementary Table 5, Supplementary Data 2)11.
MSH3 expression was not associated with any of the lead SNPs on chromosome 5 nor with variants in exon 1 of MSH3 at a significance level of 0.05/7 = 0.0071 (Supplementary Table 6A) in quantitative PCR experiments with blood-derived RNA. Expression of PMS2 did not correlate with the lead SNP on chromosome 7 (Supplementary Table 6B). Despite these associations not reaching significance, the effect direction in our analyses and in GTEx was the same for the SNPs available at this portal.
Discussion
Although penetrance of the XDP-causing mutation is widely considered to be complete, only at ~70 years of age all hemizygous SVA retrotransposon carriers will invariably manifest the phenotype. In our cohort, 94.5%, 57.0%, 19.2%, and 3.5% of individuals were not affected by XDP when they were 30, 40, 50, and 60 years old, respectively. Close to 50% of this AAO variability is due to the polymorphic repeat within the disease-causing SVA retrotransposon insertion4,5. Using a GWAS approach, we explored additional genetic modifiers of age-related disease penetrance in 353 unrelated XDP patients and identified three genome-wide significant signals. Combined with the influence of the hexanucleotide repeat in the XDP haplotype, these signals account for nearly two-thirds of the AAO variability in XDP.
Our study has several noteworthy and somewhat unexpected implications, the importance of which reaches beyond XDP. Although to date, over 50,000 unique associations between genetic variants and diseases or traits at genome-wide significance have been reported12,13, they typically required large sample sizes (>1,000 participants) and had small effect sizes (often <10% of heritability/variability explained by the sum of several associated loci)13,14,15. Our finding is particularly interesting and encouraging, as it demonstrates that significant modifiers of disease penetrance can be detected even in relatively small but carefully selected and homogeneous samples of mutation carriers. The significance of the MSH3-related locus—and the likely large effect that it exerts—are further supported by the near-significant effect that was observed in an even smaller-sized sample of HD patients (n = 218; TRACK-HD cohort)16.
A specific analytical challenge was the high degree of genetic homology in our sample stemming from a small locale with close relatedness. To account for this, we estimated the cryptic relatedness, excluded samples that were related (first cousins or closer) and included principal components into the analysis to control for any further population stratification. After adjusting the significance level by Bonferroni correction, a relatively high number of samples were significantly associated with the AAO. Since most of the significant variants were imputed, all of them were in strong linkage disequilibrium with the three lead SNPs.
The strongest signal was found on chromosome 5 within the MSH3 gene, where alternative alleles at two independent loci are associated with an earlier disease onset. On the other hand, the alternative allele of the chromosome 7 signal is correlated with the AAO increase in our patients. Interestingly, although strictly speaking not within the associated region on chromosome 7, the PMS2 gene is situated in close proximity to it. This is remarkable given that both MSH3 and PMS2 encode proteins involved in DNA mismatch repair (MMR), a cellular pathway responsible for the repair of mismatched base pairs in DNA. It has been hypothesized that the MMR process introduces instability in stretches of various, partially unwound repeat expansions in an attempt to amend the mismatch. Indeed, the downregulation17 or reduced expression18 of MSH3 has been shown to reduce instability and expansion of tri- and hexanucleotide repeats. Absence or reduced expression of PMS2 in mouse or cellular models provided contradictory results, i.e., reduction19 and accumulation20 of repeat expansions. Thus, the two genes that are either within (MSH3) or directly adjacent (PMS2) to the regions associated with AAO in XDP in our GWAS encode proteins that not only interact with each other and participate in the same cellular pathway, but their combined action has also been hypothesized to contribute to instability in stretches of nucleotide repeats similar to those within the XDP-causing insertion. Although the pathway analysis that we performed did not indicate any genetic pathway significantly associated with AAO at the stringent significance threshold that we used, this is not surprising, given our low sample size and consequently the relatively low power of this analysis.
Importantly, in 2017, MSH3 was linked to HD disease progression in a human study that also employed a GWAS16. More recently, MSH3 and PMS2 were implicated in modifying AAO in HD through GWAS approaches, albeit in much larger sample sizes (>9,000 patients)7. HD is a severe movement disorder caused by trinucleotide repeat (CAG)n expansions in the coding region of the HTT gene resulting in an extended stretch of glutamine residues. Although the main presentation of HD is chorea, neuropathologically, HD and XDP both affect primarily the striatum and display loss of medium spiny neurons21,22. Both diseases typically affect patients in their early forties and pre-symptomatic neurodegeneration of the striatum is a phenomenon seen in both HD21 and XDP23,24. Furthermore, clinical signs of HD may include dystonia and parkinsonism. Finally, HD and XDP are adult-onset neurodegenerative disorders precipitated/modified by nucleotide repeat expansions, while missense variants in both disease genes, HTT and TAF1, respectively, cause a childhood-onset neurodevelopmental syndrome with intellectual disability25,26.
None of the SNPs in genes encoding other proteins relevant to MMR reached genome-wide significance in our study, despite the fact that some of them (i) showed signals of various strengths in the recent GWAS investigating association with AAO in >9,000 HD patients7 or (ii) have been reported as associated with somatic expansion scores in blood DNA of >700 HD patients27. Possibly, our study was not powered to detect signals beyond the strongest associations that we were already able to identify. Alternatively, these other MMR genes may not play an important role in XDP, or the modifying alleles may be absent or much less common in the Filipino population.
Such population-specific effects apply to length polymorphisms in exon 1 of MSH3. Namely, while the frequency of the 6a allele (wild-type length) in a mixed-ethnicity HD patient population is comparable to that in our study (~58%6 vs. 54%), the prevalence of the shortest (3a) and longest (7a) MSH3 alleles differed considerably (~26% and ~10%6 in HD vs. 10% and 32% in XDP). In the East Asian population, the frequency of the 7a allele also seems to be higher in comparison to the 3a allele (Supplementary Table 3). Furthermore, out of 16 different haplotypes previously detected in ethnically heterogeneous HD and myotonic dystrophy type 1 (DM1, OMIM # 160900) cohorts6, only six were observed in our exclusively Filipino XDP cohort of similar size (3a, 6a, 7a, 7b, 7c, and 7d, with the latter three arising from 7a and various combinations of two rare SNPs (rs2405875, p.Ala54Ala and rs2405877, p.Pro64Ala) in several patients). Nevertheless, the 3a allele is associated with delayed disease onset in HD (increasing AAO by 1 year), as is in our study (increasing AAO by 2 years)6. The same protective association was reported for HD progression and for AAO in DM1 patients6, although this could not be shown in relatively small cohorts of spinocerebellar ataxia 3 (SCA3, OMIM # 109150) and Friedreich’s ataxia (OMIM # 229300)28. In addition, we demonstrated that the longest (7a) MSH3 allele correlated with an earlier disease onset (almost 3 years) in homozygous carriers compared to patients with wild-type MSH3 length (6a). In a study of >100 HD patients, the 3a and 7a alleles were linked with lower and higher MSH3 expression in comparison to 6a, respectively6. Our expression analyses using blood-derived RNA observed the same effect that, however, did not reach statistical significance, possibly because of low sample size (Supplementary Table 6A). The GTEx portal queries revealed an increase of MSH3 and PMS2 expression associated with alternative alleles in rs245013 and rs62456190, respectively, and a reduction of MSH3 expression correlated with an alternative allele in rs1650697, showing the same effect expression as our own analyses (Supplementary Table 6). Of note, eQTL data imply that the loci we identified may explain a fraction of the expression differences in our genes of interest and serve as a guide as to which SNPs/genes to follow-up on in future functional studies. Out of >700 SNPs genotyped or imputed in our study and accessible through the GTEx portal, <50 showed genome-wide significance in our GWAS. Possible reasons for this relatively low number include (i) small minor allele frequency in the studied population (XDP patients) and (ii) limited sample size not allowing for the detection of small effects. On the other hand, SNPs not present as eQTLs in the GTEx, but identified through our GWAS, may tag a modifier acting through mechanisms other than a change of gene expression. For example, very recently, nuclear localization29,30 and nuclear export29 signals (NLSs and NLEs, respectively) have been identified within MSH3 as important for determining the levels of this protein in the nucleus and its ability to cross the nuclear envelope. Furthermore, NLS1 (encoded by MSH3 exon 2) lies in close proximity to the length polymorphism in exon 1 and it has been shown that the shortest MSH3 (encoded by the 3a allele) is more prone to staying in the cytoplasm in comparison to the wildtype (6a) protein29. This is in agreement with the increase of AAO that is observed in XDP patients carrying the 3a allele, as the absence of MSH3 from nucleus would prevent it from introducing instability and would thus be protective.
From a personalized medicine point of view, our findings may help prioritize patients for clinical trials and may even serve as a basis for gene- or modifier-targeted therapeutic approaches. The recent randomized, double-blind, phase 1-2a trial with an antisense oligonucleotide in patients with early HD resulted in dose-dependent reductions in concentrations of mutant huntingtin31. Although it is conceivable that similar treatment options may be applicable to XDP, currently, it is unclear whether repeats with the SVA insertion are expressed or targetable. Thus, lowering MSH3 levels and thereby negatively influencing somatic instability (as was previously suggested for HD32) may be, at present, a more attainable treatment option for XDP.
In conclusion, our GWAS for genetic factors influencing age-related penetrance of XDP revealed strong signals within biologically relevant genes implying shared disease-modifying pathways with HD. More generally speaking, our findings may bear additional potential for a better understanding of other repeat expansion disorders.
Methods
Study participants and ethical review
All individuals (patients and controls) investigated in the present study were of Filipino ethnic origin. Upon genome-wide SNP genotyping of DNA samples from 458 men with XDP, eight samples with low genotyping quality and samples from 97 patients related to the included individuals were removed before further analyses (Supplementary Table 1). Therefore, pairwise kinship coefficients were calculated for all samples and pairs with a kinship coefficient greater or equal 0.125 (first-cousin or closer) were defined as related. From each resulting family cluster, a single individual was selected based on the call rate on genotype level. Thus, we performed the GWAS in the remaining 353 participants (332 previously reported3,4,5,33,34 and 21 newly analyzed). Samples from 37 more recently enrolled XDP patients and 162 healthy Filipino control individuals were investigated in post-GWAS genetic analyses. All participants gave informed consent and underwent a standardized neurological examination by movement disorder specialists. Participant enrollment and data analysis were approved by the Ethics Committees of the University of Lübeck, Germany, Massachusetts General Hospital, Boston, USA, Metropolitan Medical Center, Manila, Philippines, and Jose Reyes Medical Center, Manila, Philippines.
Genetic and statistical analyses
Genomic DNA of study participants was extracted from peripheral blood leukocytes. The presence of the SVA retrotransposon insertion in intron 32 of TAF1 and the number of hexanucleotide repeats within this insertion was investigated by capillary-electrophoresis-based fragment analysis performed on the ABI 3500xL Genetic Analyzer (Applied Biosystems) and analyzed using GeneMapper software (Applied Biosystems). The fragments examined in this way were initially PCR amplified using primers and conditions listed in Supplementary Table 74. To enable sizing of the repeats, one of the primers was labeled with a FAM tag.
DNA was subsequently analyzed by genome-wide SNP genotyping using the Infinium Global Screening Array with Custom Content (GSA; Illumina Inc.), which contains 645,896 variants. The 443,059 SNPs that passed quality control (PLINK1.9, SNP call rate >98%, minor allele frequency (MAF) > 1%, sample call rate >98%, deviation from mean heterozygosity ≤5 standard deviations) were imputed using SHAPEIT235 + IMPUTE236 with the public part of the HRC reference panel (release 1.1, The European Genome-phenome Archive, EGAS00001001710)37 to a total of 39,083,699 variants. After filtering out all SNPs with a minor allele frequency lower than 5% or an imputation info score below 0.3, a total of 4,990,999 SNPs remained to be analyzed for association. As an additional quality control step, we checked our samples for deviations from the East Asian population (Supplementary Fig. 3).
rs1650697 was additionally genotyped by Sanger sequencing of MSH3 exon 1 in 380 patients and 162 healthy Filipino controls (Supplementary Tables 1 and 7). We used gnomAD38 to assess the frequencies of these variants globally, and in East and South Asian populations.
To evaluate the association and impact of different genetic variants on AAO in XDP, we employed a multi-step approach. Initially, we performed a GWAS on normalized AAO (i.e., centralized by mean and standardized by standard deviation) to identify candidate SNPs using a linear model, taking into account genotype uncertainty resulting from imputation (SNPTEST v2.5.1). We included the polymorphic (CCCTCT)n repeat number as a covariate in the model. Hence, the resulting regression estimates were adjusted for the repeat number. Additionally, we included the first 10 principal components into the model, to adjust for a potential population stratification. We considered SNPs with a p-value below 5.0 × 10−8 as significant, to account for multiple testing in the GWAS context.
Afterward, we performed a pathway analysis (MAGMA v1.08)39 aggregating the previously calculated p-values in 11,348 genes and 14,571 different genetic pathways40. Accounting for the number of genes and pathways, we considered genes with a p-value below 4.4 × 10−6 (0.05/11348) and pathways with a p-value below 3.4 × 10−6 as significant.
Next, significantly associated SNPs were reanalyzed using a conditional strategy (using R3.6)41. Therefore, we iteratively recalculated the regression coefficients of all SNPs per region adjusted for the SNP with the lowest p-value in this region, in order to determine whether one or several SNPs are responsible for the signal within a given chromosomal region.
Unsolicited sequence changes adjacent to rs1650697 were genotyped using Sanger sequencing and analyzed using linear models, again adjusting for the hexanucleotide repeat number to evaluate the association of these variants with AAO. If more than one single sequence change was found in a region, we estimated haplotypes including these variants using a standard expectation maximization algorithm and analyzed the influence of different haplotypes on AAO by linear regression, and adjusted for hexanucleotide repeat number. In cases where regions included in-frame indels, we additionally analyzed changes in length at the protein level by applying linear regression on AAO adjusted for hexanucleotide repeat number.
Since we observed an overlap between single-nucleotide polymorphism associated with AAO in HD7 and variants detected by our GWAS, we constructed a PRS for AAO in HD using gender-specific effect estimates of the 21 lead SNPs from7 and calculated this for all of our 353 XDP patients from the GWAS. With this, we calculated a linear regression model predicting AAO in XDP by the SVA repeat number and the PRS.
Expression analyses
We investigated the expression traits associated with variants of interest using the GTEx portal (gtexportal.org, Data Source: GTEx Analysis Release V8 (dbGaP Accession phs000424.v8.p2))8, the BrainSeq dataset9 from the eQTL Catalog10, and the UKBEC dataset11.
In addition, we analyzed MSH3 and PMS2 expression levels in blood via quantitative PCR (qPCR) analysis of cDNA synthesized from blood-derived RNA (n = 40). Real-time qPCR was performed using SYBR Green chemistry (Fermentas, Waltham, MA) on a Light Cycler 480 (Roche Applied Science). Expression levels of genes, measured in triplicates, were determined using the Advanced Relative Quantification method by the Roche LightCycler software (release 1.5.0)13 and normalized against the expression of GAPDH and YWHAZ. Amplification conditions and primers are listed in Supplementary Table 7. The influence of investigated genotypes on corresponding expression levels was analyzed using linear models.
Reporting summary
Further information on research design is available in the Nature Research Reporting Summary linked to this article.
Data availability
The authors declare that all data supporting the findings of this study are included in the article and its supplementary information files or are available from the corresponding authors upon reasonable request. Furthermore, the authors used the following databases: (i) The Haplotype Reference Consortium (HRC) reference panel (release 1.1, The European Genome-phenome Archive EGAD00001002729), (ii) The Genome Aggregation Database (gnomAD) (v2.1.1, https://gnomad.broadinstitute.org/gene/ENSG00000113318?dataset=gnomad_r2_1), (iii) The Genotype-Tissue Expression (GTEx) project (https://www.gtexportal.org/home/gene/MSH3#eQTLBlock, Analysis Release V8 (dbGaP Accession phs000424.v8.p2)), (iv) the BrainSeq dataset from the eQTL (expression quantitative trait locus) Catalog (https://www.ebi.ac.uk/eqtl/Studies/), (v) as well as the United Kingdom Brain Expression Consortium (UKBEC) (http://www.braineac.org/). The GWAS summary statistics (source data for Fig. 1) are available through the GWAS Catalog accession GCST90014263 (ftp://ftp.ebi.ac.uk/pub/databases/gwas/summary_statistics/GCST90014263). Source data are provided with this paper.
Code availability
The authors declare that the codes used in this study are available from the corresponding author upon reasonable request.
References
Pauly, M. G. et al. Expanding data collection for the MDSGene database: X-linked dystonia-parkinsonism as use case example. Mov. Disord. 35, 1933–1938 (2020).
Makino, S. et al. Reduced neuron-specific expression of the TAF1 gene is associated with X-linked dystonia-parkinsonism. Am. J. Hum. Genet. 80, 393–406 (2007).
Domingo, A. et al. New insights into the genetics of X-linked dystonia-parkinsonism (XDP, DYT3). Eur. J. Hum. Genet. 23, 1334–1340 (2015).
Bragg, D. C. et al. Disease onset in X-linked dystonia-parkinsonism correlates with expansion of a hexameric repeat within an SVA retrotransposon in TAF1. Proc. Natl Acad. Sci. USA 114, E11020–E11028 (2017).
Westenberger, A. et al. A hexanucleotide repeat modifies expressivity of X-linked dystonia parkinsonism. Ann. Neurol. 85, 812–822 (2019).
Flower, M. et al. MSH3 modifies somatic instability and disease severity in Huntington’s and myotonic dystrophy type 1. Brain 142, 1876–1886 (2019).
Gusella, J. F. CAG repeat not polyglutamine length determines timing of Huntington’s disease onset article CAG repeat not polyglutamine length determines timing of Huntington’s disease onset. https://doi.org/10.1016/j.cell.2019.06.036 (2019).
The Genotype-Tissue Expression (GTEx) Project. https://gtexportal.org/; 11/30/20 (2021).
Jaffe, A. E. et al. Developmental and genetic regulation of the human cortex transcriptome illuminate schizophrenia pathogenesis. Nat. Neurosci. 21, 1117–1125 (2018).
Kerimov, N. et al. eQTL catalogue: a compendium of uniformly processed human gene expression and splicing QTLs. Preprint at bioRxiv https://doi.org/10.1101/2020.01.29.924266 (2020).
Ramasamy, A. et al. Genetic variability in the regulation of gene expression in ten regions of the human brain. Nat. Neurosci. 17, 1418–1428 (2014).
Buniello, A. et al. The NHGRI-EBI GWAS Catalog of published genome-wide association studies, targeted arrays and summary statistics 2019. Nucleic Acids Res. 47, D1005–D1012 (2019).
Tam, V. et al. Benefits and limitations of genome-wide association studies. Nat. Rev. Genet. 20, 467–484 (2019).
Manolio, T. A. Bringing genome-wide association findings into clinical use. Nat. Rev. Genet. 14, 549–558 (2013).
Manolio, T. A. et al. Finding the missing heritability of complex diseases. Nature 461, 747–753 (2009).
Moss, D. J. H. et al. Identification of genetic variants associated with Huntington’s disease progression: a genome-wide association study. Lancet Neurol. 16, 701–711 (2017).
Halabi, A., Ditch, S., Wang, J. & Grabczyk, E. DNA mismatch repair complex MutSβ promotes GAA·TTC repeat expansion in human cells. J. Biol. Chem. 287, 29958–29967 (2012).
Tomé, S. et al. MSH3 polymorphisms and protein levels affect CAG repeat instability in Huntington’s disease mice. PLoS Genet. 9, e1003280 (2013).
Gomes-Pereira, M., Fortune, M. T., Ingram, L., McAbney, J. P. & Monckton, D. G. Pms2 is a genetic enhancer of trinucleotide CAG·CTG repeat somatic mosaicism: implications for the mechanism of triplet repeat expansion. Hum. Mol. Genet. 13, 1815–1825 (2004).
Bourn, R. L. et al. Pms2 suppresses large expansions of the (GAA·TTC)n sequence in neuronal tissues. PLoS ONE 7, e47085 (2012).
Aylward, E. H. et al. Onset and rate of striatal atrophy in preclinical Huntington disease. Neurology 63, 66–72 (2004).
Hanssen, H. et al. Imaging gradual neurodegeneration in a basal ganglia model disease. Ann. Neurol. 86, 517–526 (2019).
Walter, U. et al. Sonographic alteration of substantia nigra is related to parkinsonism-predominant course of X-linked dystonia-parkinsonism. Park. Relat. Disord. 37, 43–49 (2017).
Hanssen, H. et al. Striatal neurodegeneration in the presymptomatic phase of X-linked dystonia-parkinsonism. Brain 141, 2995–3008 (2018).
O’Rawe, J. A. et al. TAF1 variants are associated with dysmorphic features, intellectual disability, and neurological manifestations. Am. J. Hum. Genet. 97, 922–932 (2015).
Lopes, F. et al. Identification of novel genetic causes of Rett syndrome-like phenotypes. J. Med. Genet. 53, 190–199 (2016).
Ciosi, M. et al. A genetic association study of glutamine-encoding DNA sequence structures, somatic CAG expansion, and DNA repair gene variants, with Huntington disease clinical outcomes. EBioMedicine 48, 568–580 (2019).
Yau, W. Y. et al. The repeat variant in MSH3 is not a genetic modifier for spinocerebellar ataxia type 3 and Friedreich’s ataxia. Brain 143, e25 (2020).
Tseng-Rogenski, S. S. et al. The human DNA mismatch repair protein MSH3 contains nuclear localization and export signals that enable nuclear-cytosolic shuttling in response to inflammation. Mol. Cell. Biol. 40, e00029-20 (2020).
Williams, G. M. et al. HDAC3 deacetylates the DNA mismatch repair factor MutSβ to stimulate triplet repeat expansions. Proc. Natl Acad. Sci. USA 2020, 13223 (2020).
Tabrizi, S. J. et al. Targeting huntingtin expression in patients with Huntington’s disease. N. Engl. J. Med. 380, 2307–2316 (2019).
Tabrizi, S. J., Ghosh, R. & Leavitt, B. R. Huntingtin lowering strategies for disease modification in Huntington’s disease. Neuron 101, 801–819 (2019).
Aneichyk, T. et al. Dissecting the causal mechanism of X-linked dystonia-parkinsonism by integrating genome and transcriptome assembly. Cell 172, 897–909.e21 (2018).
Nolte, D., Niemann, S. & Muller, U. Specific sequence changes in multiple transcript system DYT3 are associated with X-linked dystonia parkinsonism. Proc. Natl Acad. Sci. USA 100, 10347–10352 (2003).
O’Connell, J. et al. A general approach for haplotype phasing across the full spectrum of relatedness. PLoS Genet. 10, e1004234 (2014).
Howie, B. N., Donnelly, P. & Marchini, J. A flexible and accurate genotype imputation method for the next generation of genome-wide association studies. PLoS Genet. 5, e1000529 (2009).
McCarthy, S. et al. A reference panel of 64,976 haplotypes for genotype imputation. Nat. Genet. 48, 1279–1283 (2016).
Karczewski, K. J. et al. The mutational constraint spectrum quantified from variation in 141,456 humans. Nature 581, 434–443 (2020).
de Leeuw, C. A., Mooij, J. M., Heskes, T. & Posthuma, D. MAGMA: generalized gene-set analysis of GWAS data. PLoS Comput. Biol. 11, e1004219 (2015).
Mi, H., Muruganujan, A., Ebert, D., Huang, X. & Thomas, P. D. PANTHER version 14: More genomes, a new PANTHER GO-slim and improvements in enrichment analysis tools. Nucleic Acids Res. 47, D419–D426 (2019).
Spain, S. L. & Barrett, J. C. Strategies for fine-mapping complex traits. Hum. Mol. Genet. 24, R111–R119 (2015).
Acknowledgements
The study was supported by the Deutsche Forschungsgemeinschaft (DFG; FOR 2488 to C.K., N.B., K.L., F.J.K., I.R.K., and A.W.; SFB 936 [project C5] to C.K.), the Collaborative Center for X-linked Dystonia-Parkinsonism at Massachusetts General Hospital (to N.B., C.K., L.J.O., N.S., T.M.-B., and C.G.), the Else Kröner Fresenius Foundation (to N.B.), intramural funds from the University of Lübeck (to C.K. and M.D.-M.), and a career development award from the Hermann and Lilly Schilling Foundation (to C.K.). Furthermore, this work received infrastructure support by the DFG Cluster of Excellence “Precision Medicine in Chronic Inflammation” (PMI, EXC2167 to A.F.). We thank the XDP patients and their relatives for their participation in this study; the Sunshine Care Foundation and members of their staff (including genetic councilors N.G.M. Ganza and B.B. Lagarde) for the infrastructure they have established and for their devoted and hard work; Dr. L.V. Lee, Dr. R. Kaji, and Dr. T. Kawarai for referral of DNA samples and clinical information; Dr. L. Bertram who was involved in early scientific discussions of the study, acquisition of funding, and supervision of the collection of the GSA-based genotyping data; and H. Pawlack for technical assistance, including genotyping of DNA samples.
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Contributions
B.-H.L. performed the genome-wide association analysis and data interpretation and co-drafted the manuscript; C.K. participated in conception and design, collected and interpreted data, and significantly contributed to the writing of the manuscript and later its critical revision; J.P. did the sequencing, genotyping, and expression analyses and critically revised the manuscript; A.D. interpreted data and critically revised the manuscript; N.B. collected and analyzed clinical data and critically revised the manuscript; K.G. interpreted data and critically revised the manuscript; R.L.R., R.D.J., G.S., and C.C.E.D. collected and interpreted data and critically revised the manuscript; M.W. performed array genotyping (GSA) for the discovery study and critically revised the manuscript; S.S. and M.D.-M. interpreted data and critically revised the manuscript; J.Q. and P.O. performed sequencing/genotyping and critically revised the manuscript; P.A., C.G., N.S., T.M.-B., U.M., and H.H. collected data and critically revised the manuscript; F.K., A.R., and P.B. did the genotyping analysis and critically revised the manuscript; A.F. performed array genotyping (GSA) for the discovery study and critically revised the manuscript; V.D. collected data and critically revised the manuscript; K.L. participated in conception and design and critically revised the manuscript; L.J.O. performed genotyping and the haplotype analysis, interpreted data and critically revised the manuscript; F.J.K. participated in conception and design, did the expression analyses, interpreted data, and critically revised the manuscript; I.R.K. participated in conception and design, did the genome-wide association analysis, interpreted data, and critically revised the manuscript; A.W. participated in conception and design, analyzed and interpreted data, and co-drafted the manuscript. All authors gave their final approval of the version to be published and agreed to be accountable for all aspects of the work.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Peer review information Nature Communications thanks Günter Höglinger, Lesley Jones and the other, anonymous, reviewer for their contribution to the peer review of this work. Peer reviewer reports are available.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Laabs, BH., Klein, C., Pozojevic, J. et al. Identifying genetic modifiers of age-associated penetrance in X-linked dystonia-parkinsonism. Nat Commun 12, 3216 (2021). https://doi.org/10.1038/s41467-021-23491-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-021-23491-4
This article is cited by
-
Mechanisms underlying phenotypic variation in neurogenetic disorders
Nature Reviews Neurology (2023)
-
Oculomotor abnormalities indicate early executive dysfunction in prodromal X-linked dystonia-parkinsonism (XDP)
Journal of Neurology (2023)
-
Identification of representative trees in random forests based on a new tree-based distance measure
Advances in Data Analysis and Classification (2023)
-
Endemic parkinsonism: clusters, biology and clinical features
Nature Reviews Neurology (2023)
-
Tissue-specific and repeat length-dependent somatic instability of the X-linked dystonia parkinsonism-associated CCCTCT repeat
Acta Neuropathologica Communications (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
