
Abstract
Over the last decade, genetic studies, including genome-wide association studies (GWAS), have accelerated the discovery of genes and genomic regions contributing to primary open-angle glaucoma (POAG), a leading cause of irreversible vision loss. Here, we review the findings of genetic studies of POAG published in English prior to September 2019. In total, 74 genomic regions have been associated at a genome-wide level of significance with POAG susceptibility. Recent POAG GWAS provide not only insight into global and ethnic-specific genetic risk factors for POAG susceptibility across populations of diverse ancestry, but also important functional insights underlying biological mechanisms of glaucoma pathogenesis. In this review, we also summarize the genetic overlap between POAG, glaucoma endophenotypes, such as intraocular pressure and vertical cup–disc ratio (VCDR), and other eye disorders. We also discuss approaches recently developed to increase power for POAG locus discovery and to predict POAG risk. Finally, we discuss the recent development of POAG gene-based therapies and future strategies to treat glaucoma effectively. Understanding the genetic architecture of POAG is essential for an earlier diagnosis of this common eye disorder, predictive testing of at-risk patients, and design of gene-based targeted medical therapies none of which are currently available.
摘要
原发性开角型青光眼(POAG)是导致不可逆视力丧失的主要原因, 在过去十年中, 遗传学研究的新发现, 包括全基因组关联研究, 加速了POAG基因和基因组区域的新发现。我们回顾了2019年9月之前英文发表的关于POAG遗传研究的研究成果。在全基因组水平, 共有74个基因组区域与POAG的易感性相关。近期关于POAG的 GWAS研究不仅提供了全球以及不同种族的POAG基因易感性的遗传危险因素, 还提供了这些易感基因在青光眼病理发病机制中的重要作用。在这篇综述中, 我们总结了POAG易感基因之间的相互作用, 青光眼内在表型 (如眼内压和垂直杯盘比) 以及其与他眼病之间的遗传的相关性。我们还讨论了最近研发的增加POAG基因座发现能力及预测POAG风险的方法。最后, 我们讨论了基于POAG基因治疗的最新进展以及有效治疗青光眼的未来策略。了解POAG的遗传信息对于早期诊断这种常见的眼部疾病, 对高危患者进行早期预测以及设计目前尚无可用的基于基因的靶向药物治疗至关重要。
Similar content being viewed by others

POAG genetics advances
Recent GWAS of POAG led to the discovery of numerous genetic loci
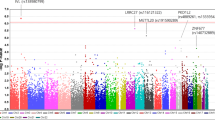
In the last decade, genome-wide association studies (GWAS) have rapidly accelerated the discovery of genetic determinants of numerous diseases and complex traits, with more than 10,000 significant genome-wide associations reported to date [1,2,3]. Unsurprisingly, this GWAS success in identifying risk loci has also applied to primary open-glaucoma (POAG). As of 2017, 16 genomic regions associated with POAG at a genome-wide level of significance (P < 5 × 10−8) had been reported [4], with most studies conducted in European and Asian populations [5,6,7,8,9,10,11,12,13,14]. In the last 3 years, many other loci that contribute to POAG susceptibility have been discovered [15,16,17,18], bringing the total number of POAG loci to 74 (Table 1). This recent, rapid increase in the discovery of POAG loci is likely due to several factors: (1) the emergence of large and multiethnic biobank-based cohorts, such as the UK Biobank [19, 20] and the Kaiser Permanente GERA [21, 22] cohorts; (2) the availability of summary statistics of published GWAS to the scientific community (e.g., GWAS catalogue [2]), which has enabled more rapid confirmation of association loci; (3) the combination of the results from different GWAS in large multiethnic meta-analyses [15, 18]; and (4) the application of recently developed approaches (e.g., gene-based analysis) using GWAS summary statistics [23, 24].
Shared and ethnic-specific genetic associations
Recent GWAS provide insight into global and ethnic-specific genetic risk factors for POAG susceptibility across populations of diverse ancestry. As of 2017, five genomic regions had been reported to be associated with POAG in populations of Asian ancestry, including ABCA1, PMM2, and CDKN2B-AS1 [8, 10, 25]. Associations at all of these loci were either previously reported or have been largely confirmed in populations of European ancestry [9, 13, 15, 18]. A recent Japanese GWAS of POAG confirmed genome-wide associations at known POAG loci and identified seven novel loci, including FNDC3B, ANKRD55-MAP3K1, LMX1B, LHPP, HMGA2, MEIS2, and LOXL1 [17]. Among these newly identified POAG loci, three loci, including LHPP, HMGA2, and MEIS2, replicated neither in European population nor in African population from this study. This suggests the possible existence of genetic-specificities at those LHPP, HMGA2, and MEIS2 loci for POAG for Japanese individuals, and future studies in multiethnic populations may confirm those Asian-specific POAG genetic associations.
Recently, progress has been made in the identification of genetic variants associated with POAG in populations of African ancestry [26,27,28]. Because individuals of African ancestry have three to five times increased POAG risk, and have worse visual field damage and disease progression compared with other populations [29, 30], it is important to elucidate the genetic contribution to POAG pathogenesis to aid identification of high-risk groups. Recent GWAS of glaucoma/POAG in African ancestry populations enabled the identification of genome-wide significant associations at DNAJC24-ELP4, TRIM9-TMX1, FAM86A-RBFOX1, EXOC4, and EN04 loci, and suggestive associations at COL21A1-DST and MNS1-ZNF280D [26,27,28]. However, these associations still await validation in external cohorts, with the exception of TRIM9-TMX1 and FAM86A-RBFOX1, which reached nominal significance in African Americans from GERA [15] and in African Americans from BioMe [28], respectively. In addition, recent genetic studies conducted in populations of African ancestry attempted to replicate the associations at loci previously identified in GWAS predominantly conducted in populations of European or Asian ancestry. While some studies could not replicate any of these loci [26, 31], others provided evidence of association at TXNRD2, FNDC3B, 8q22, AFAP1, TMC01, or CDKN2B-AS1 loci [27, 28, 32, 33]. Using a “local” replication strategy that considered different linkage disequilibrium (LD) patterns across study populations, Bonnemaijer et al. [28] replicated in their African cohorts, the POAG associations at TXNRD2, TMC01, and CDKN2B-AS1, originally identified in European populations [5, 13]. This suggests that POAG-susceptibility loci identified in cohorts of European or Asian ancestry may be relevant to populations of African ancestry. Genetic risk scores based on the previously reported and newly discovered POAG-genetic variants have been shown to predict POAG and explain up to 4% of the overall POAG risk in populations of African ancestry [15, 26,27,28]. These findings provide new insight into the genetic architecture of POAG in populations of African ancestry, and future studies investigating the genetic complexity of POAG in these populations would request studies of extremely large size.
In contrast to the recent progress made in populations of African ancestry, the genetics of POAG in Hispanic/Latino populations remain largely unexplored, and to date, no genome-wide significant signal has been detected in Hispanic/Latino populations. Choquet et al. [15] examined the associations of the loci previously identified in GWAS of European or Asian populations in their Hispanic/Latino sample, consisting of 411 POAG cases and 4778 controls. Three loci replicated after correction for multiple testing, including TMC01, near CDKN1A, and CDKN2B-AS1; and two others, AFAP1 and ABCA1, were nominally significant. When investigating the associations of the novel POAG loci identified in this study, a suggestive association between PDE7B and POAG risk in Hispanic/Latinos was observed [15]. Further, previously known and newly discovered genetic variants from this study explained 3.3% of POAG variance in Hispanic/Latinos from GERA. Future efforts to identify the genetic factors for POAG risk in this ethnicity may aid in understanding of the genetic architecture of POAG.
Importance of genetic ancestry
Beyond genomic region identification, recent genetic studies of POAG have enabled the determination of ancestry and population substructure [15, 34]. Because variation in POAG prevalence has been observed across populations of diverse ancestry, especially in African Americans who have a higher risk for developing POAG, it is important to investigate whether differences in POAG prevalence are due to genetic ancestry. Choquet et al. [15] investigated the association between genetic ancestry, as represented by genetic principal components, and the risk of POAG within each of the four GERA ethnic groups. A higher risk of POAG was associated with greater northern (versus southern) East Asian ancestry in the East Asian group, greater Native American (versus European) ancestry in the Hispanic/Latino group, and greater African (versus European) ancestry in the African American group. These findings suggest that this within-group variation could be due to genetic risk factors that correlate with genetic ancestry. Consistently, a recent study [34] conducted in an African American cohort assessed the local genetic ancestry at CDKN2B-AS1, an important POAG-associated locus established in populations of European and Asian ancestry. Interestingly, a significant association was observed between POAG risk and local African genetic ancestry at CDKN2B-AS1, and on average, POAG cases were of 90% African descent compared with 58% for controls [34]. It is noteworthy that in this study [34], no significant single SNP-POAG associations at this locus were detected after correcting for multiple testing. These findings highlight the importance of considering the variability in LD patterns across populations and genetic heterogeneity when conducting genetic studies.
Genetic loci discovery led to biological pathway discovery
Although GWAS-identified associations do not directly highlight a specific gene or mechanism, several genetic studies conducted follow-up experiments of candidate genes within identified POAG-genomic regions using animal models and human cell lines. Before 2017, only a few POAG-candidate genes (i.e., SIX6, CDKN2B-AS, and CAV1/2) have been investigated in functional studies [35,36,37,38]. SIX6 encodes a homeobox protein and plays an important role in the development of the eye, especially the morphology of the optic nerve and the formation of the retina [39,40,41]. In vivo (zebrafish) and in vitro assays demonstrated that SIX6 risk variants attenuated protein function, leading to a reduction in the number of retinal ganglion cells, which are the primary cell type affected in glaucoma, thereby increasing POAG risk [35]. Consistently, a SIX6 risk variant (rs33912345) increased the expression of CDKN2A (another well-established POAG and normal tension glaucoma locus), resulting in the senescence of retinal ganglion cells in cell line, animal models, and human glaucoma retinas [37]. Further, mice homozygous for a deletion in CDKN2B-AS are more vulnerable to retinal ganglion cell loss in response to elevated intraocular pressure (IOP), compared with wild-type and heterozygous animals [36]. Vulnerability to retinal ganglion cell loss manifests by microglial reactivity signs both in the retina and the optic nerve of mutated mice [36], which are early indicators of glaucoma onset or progression [42, 43].
CAV1, which is located within the glaucoma susceptibility locus CAV1/CAV2, encodes the caveolin 1. CAV1 has been shown to maintain normal IOP levels by participating to the caveolae formation in Schlemm’s canal and trabecular meshwork (TM), thereby facilitating aqueous humour flow through the eye [38]. Similarly, FMNL2, a gene in one of the recently discovered POAG-loci, that is also associated with IOP[44], supports TM function relevant to aqueous humour outflow regulation [15]. Indeed, suppression of FMNL2 expression using small interfering RNAs (siRNAs) caused TM cell morphological modifications, thereby decreasing contractile activity and the assembly of actin stress fibres [15]. Investigation of expression profiles of genes associated with both IOP and glaucoma also revealed a high enrichment in the TM compared with other human ocular tissues [18]. Functional follow-up experiments have also been conducted for another POAG candidate gene, LMX1B [15, 17], previously reported to cause nail-patella syndrome, a rare developmental disorder, with some patients presenting a similar “glaucoma” phenotype associated with structural defects of the eye [45,46,47]. In mouse models of different genetic backgrounds, Lmx1b mutations can result in high IOP and glaucomatous nerve damage in eyes without developmental defects [15]. This suggests that LMX1B acts via elevated IOP to affect glaucoma susceptibility, with some mutations causing a condition in mice that resembles POAG. These findings provide important functional insights linking genetic susceptibility POAG loci to the underlying mechanisms of glaucoma pathogenesis.
In addition to in vivo/vitro follow-up experiments, in silico analyses turned out to be effective to prioritize the causal gene within the identified locus and to discover biological mechanisms underlying POAG disease. These include publicly available tools and datasets providing genomic annotations, epigenetic marks, drug targets, gene expression, and expression quantitative trait locus information [48,49,50,51,52,53,54]. The interpretation of the non-coding variants that account for the majority of GWAS-identified risk alleles is crucial to understand the biological mechanisms through which these risk variants act. Recent GWAS studies [16,17,18] performed pathway analyses and identified relevant biological pathways that might be involved in POAG pathogenesis; these include “epidermal growth factor receptor signalling”, “response to fluid shear stress”, “abnormal retina morphology”, and “vascular development”. Functional follow-up experiments in cell lines or animal models may confirm the involvement of these biological pathways and provide underlying mechanisms of glaucoma pathogenesis.
Pleiotropy and genetic correlations between POAG, glaucoma endophenotypes, ocular, and systematic diseases
Many of the POAG-associated loci, recently identified by GWAS, are also associated with glaucoma endophenotypes and other ocular conditions. While common variants in POAG-loci [15, 17] FNDCB3 and FMNL2 were known to be associated with IOP [44, 55], common variants in CDKN2B-AS and SIX6 [18] were known to be associated with vertical cup–disc ratio (VCDR) [56]. Similarly, while common variation at LOXL1 POAG locus [17] has been reported to be associated with exfoliation syndrome/exfoliation glaucoma [57,58,59], common variation at MYOF POAG locus [16] has been reported to be associated with refractive error [60]. In addition, common and rare variants in C9, a gene identified to be associated with POAG [16] using a gene-based approach, have been associated with age-related macular degeneration [61,62,63]. These results are consistent with results from a recent study showing significant genetic correlation between POAG and age-related macular degeneration [64].
Genetic correlation analyses, using a technique-cross-trait LD score regression [65], also revealed the relationships between POAG and systemic diseases, including type 2 diabetes and cardiovascular diseases, such as myocardial infarction and ischemic stroke [17]. These findings support previous reports, showing that pleiotropy, a term that refers to individual genetic loci that influence the risk of multiple diseases or affect variation in multiple complex traits, is pervasive [1, 3, 65,66,67].
Heritability and variance explained
Beyond the identification of genomic regions, recent GWAS of POAG have facilitated the quantification of how much of the total additive genetic variation due to segregating variants in the population is tagged by genotyped SNPs [15, 68]. This quantification of “array” or “SNP” heritability is informative with respect to the unknown genetic architecture of the disease. A recent phenome-wide heritability study conducted in the UK Biobank [20], based on self-reported disease information, reported an array heritability estimate of 26.0% (s.e. = 6.0%) for glaucoma [68]. Consistently, Choquet et al. estimated an overall heritability of 26.0% (s.e. = 1.0%) for POAG in the GERA non-Hispanic white ethnic group [15]. However, our current knowledge of genome-wide significant POAG/glaucoma SNPs explains only ∼3% of the genetic contribution to glaucoma susceptibility, suggesting that additional variants remain yet to be discovered [15].
To explain the remaining “missing” heritability of POAG, innovative approaches have been employed. Gharahkhani et al. [16] conducted a meta-analysis of genetic data from OAG and its correlated traits (e.g., IOP, optic disc parameters) to identify new loci. Using this innovative and integrative approach, they found additional loci associated with OAG (i.e., near MYOF, LINC02052, and LMO7) at genome-wide level of significance. They also identified an association with a previously unreported gene, complement factor 9 (C9) [16], using a fast and flexible set-Based Association Test method [69]. This gene would not have been identified in a standard single-variant analysis due to the limited statistical power to detect individual genetic variants with small effect sizes. Similarly, MacGregor et al. [18] conducted gene-based association analyses and identified four genes that were associated with POAG after multiple testing correction, including BICC1, SLC38A3, KALRN, and RELN. Next-generation sequencing has been recently largely used to identify novel associations between low-frequency (or rare) coding variants and disease susceptibility [70]. Zhou et al. [71] conducted a whole exome sequencing (WGS) analysis on 187 patients with early-onset advanced POAG and 103 controls without glaucoma and found enrichment of rare variants in camera-type eye development genes (i.e., CRYBA4, GAS1, GJA8, HES5, MAB21L2, NEUROD4, NR2E1, PAX6, RXRA, SLC25A25, VAX1). Rare variants identified by WGS may have much greater positive predictive values in terms of clinical application compared with common SNPs identified by GWAS, as GWAS do not necessarily identify the causal variants.
Genetics of glaucoma-related traits
The endophenotype approach to glaucoma gene discovery
While case-control GWAS remain the definitive method for identifying genetic variants associated with disease, an alternative approach is to examine a heritable quantitative trait related to the disease. Such traits are termed endophenotypes and examples for glaucoma include IOP and vertical cup–disc ratio . There are several potential advantages of an endophenotype approach. Rather than requiring data from many disease cases, data can be leveraged from healthy population samples as it is the variation of the endophenotype across the whole range of health and disease that drives the association signal. This allows many population cohorts to contribute to analyses, even if the studies have a low prevalence of disease, resulting in large sample sizes and power to detect small associations. Statistical power is also increased by analysing a continuous outcome trait rather than a binary outcome variable. In addition, examining individual traits may help better characterize how discovered genetic variants contribute to disease (e.g., IOP-increasing versus IOP-independent mechanisms for glaucoma). However, caution is required in inferring disease relevance of endophenotype associations, and further studies examining association with disease are required.
Genetic associations with IOP
IOP is the cardinal modifiable risk factor for POAG [72, 73] and is known to be a heritable trait [74]. Understanding what causes variation in the level of IOP within the normal range may shed light on the mechanisms that also contribute to high IOP and, in turn, POAG. Earlier work examining genetic associations with IOP has proven successful at identifying glaucoma risk variants [55, 75]. Combining data from 35,296 participants of 18 population studies contributing to the International Glaucoma Genetics Consortium (IGGC) led to the identification of eight genome-wide significant loci for IOP, the majority of which demonstrated significant association with POAG in independent studies [55]. However, these loci explained only a very small proportion of IOP variability. Two recent studies with considerably larger sample sizes have identified over 100 more IOP-associated loci, demonstrating the importance of a large sample size in GWAS of complex traits to identify small effect associations [44, 76].
The first of these two studies to report was a multiethnic GWAS for IOP in 69,756 individuals of the GERA cohort [44]. IOP measurements were taken as part of routine clinical care, and only measurements taken prior to any IOP-lowering treatment were considered. For participants with multiple longitudinal IOP measurements, the median value was considered; this approach was demonstrated to be more effective than just considering IOP measured at one random timepoint [44]. The GERA analysis replicated the majority of previously reported IOP-associated loci, demonstrating the validity and utility of using opportunistic IOP measurements in a clinical cohort rather than protocoled measurements in a population-based study. Reporting shortly after GERA was a GWAS of IOP in European participants of the UK Biobank study [76]. Results from the UK Biobank GWAS were then meta-analysed with IOP GWAS results from the EPIC-Norfolk Eye Study [77] and the aforementioned IGGC study [75]; the combined analysis included 139,555 participants [76]. An independent group of investigators also conducted a GWAS for IOP in UK Biobank and found similar results [18].
The GERA analysis identified 47 genome-wide significant loci, 40 of which were novel [75], and the UK Biobank analysis identified 112 genome-wide significant loci, 68 of which were novel [76]. The scale of this discovery represents a step change in our knowledge of IOP genetics. The identified loci explained 17% of the variance of IOP in the EPIC-Norfolk Eye Study [76], which is substantial given the variability of IOP caused by diurnal changes and measurement error alone. There is considerable overlap between the GERA and UK Biobank discovered loci. A meta-analysis of the two studies has not been done to date.
Among the significant results were loci at genes previously associated with POAG, but not previously known to influence IOP (AFAP1, TXNRD2, ATXN2) [44, 76]. This strongly suggests that genetic variation at these genes mediate their increased POAG risk via raised IOP, rather than via direct effects on retinal ganglion cells. Also among the significant results were four loci previously reported as conferring susceptibility to primary angle-closure glaucoma (PLEKHA7, HGF, FERMT2, GLIS3) [44, 76], suggesting that angle-closure mechanisms may contribute to variation in IOP even within the normal range.
One of the most significant novel findings was a locus at the Diacylglycerol Kinase Gamma (DGKG) gene (P = 8.9 × 10–52 in UK Biobank meta-analysis). Diacylglycerol is involved in adenosine receptor signalling, which is known to be involved in IOP regulation and is a reported target for IOP-lowering therapy [78]. More generally, DGKG is involved in lipid metabolism, adding weight to the growing evidence that lipid metabolism is a key component of IOP regulation [4].
There were multiple IOP-associated loci at genes previously associated with Mendelian childhood glaucoma (FOXC1, PITX2, LMX1B, LTBP2) [44, 76]. Other IOP-associated loci were at genes involved in ocular development (SIX3, ADAMTS18, MEIS1), eye size (RSPO1), and iris architecture (TRAF3IP1) [76]. Furthermore, genes involved in developmental processes in general were significantly enriched in the UK Biobank results [76]. These findings suggest that common genetic variation may contribute to developmental or anatomical changes that are insufficient to cause glaucoma in childhood but that may lead to a decompensation of IOP in later adult life and potentially POAG.
Post-trabecular meshwork IOP regulation
One of the most striking findings from both IOP GWAS studies was evidence for an important role of vascular endothelial processes in IOP regulation. Genes involved in “vascular endothelial cell morphology” were the most significantly enriched gene set in GERA [44], and genes involved in “angiogenesis” were the most significantly enriched gene set in the UK Biobank study [76] (both studies used different approaches for examining enrichment). In contrast to the TM, which comprises epithelial cells, Schlemm’s canal and collector channels are composed of endothelial cells that have a similar phenotype to lymphatic vessels [79]. The major drivers for the vascular endothelial gene-set enrichment were variants in ANGPT1, ANGPT2, and VEGF-C. ANGPT1 and ANGPT2 are primary TEK (receptor tyrosine kinase) ligands. Mutations of TEK cause primary congenital glaucoma [80]. This suggests that, while rare mutations affecting angiopoietin-TEK signalling can cause congenital disease, more common genetic variation with less deleterious functional consequence can cause less severe changes and a decompensation of IOP only manifest in later life. TEK receptors are highly expressed in Schlemm’s canal [81], and disruption of angiopoietin-TEK signalling in mice causes absent Schlemm’s canal development [82]. VEGF-C increases VEGFR-3 tyrosine kinase signalling in lymphatic endothelial cells and anterior chamber delivery of VEGF-C in adult mice-induced Schlemm’s canal growth and a sustained reduction in IOP [79]. Put together, there is clear emerging evidence that post-TM structures (Schlemm’s canal and collector channels) are critical for IOP regulation, challenging the dogma that POAG is primarily a disease of TM. There also appears to be potential for regulators of lymphangiogenesis as targets for glaucoma therapy.
Association of IOP loci with POAG
It is important to determine whether genetic variants associated with higher IOP also confer higher risk for POAG. The GERA investigators tested their IOP-associated loci for association with clinically coded POAG in the same GERA cohort; 89% of variants were directionally consistent with IOP-increasing risk alleles having an odds ratio estimate >1 for POAG [44]. In the UK Biobank study [76], the association of the IOP loci with POAG was examined in 3853 cases and 33,480 controls from the independent NEIGHBORHOOD study [13]. There was a strikingly linear trend between the effect estimates for IOP from UK Biobank and POAG from NEIGHBORHOOD when plotted [76]. In addition, 48 variants were nominally associated with POAG (P < 0.05), of which 14 were significant at a Bonferroni-corrected threshold [76].
Prediction of POAG using IOP-associated loci
The UK Biobank investigators also examined whether the IOP loci, together with age, sex, and three known POAG-associated polymorphisms showing no evidence of association with IOP (at MYOC, SIX6, and CDKN2B-AS1), were predictive of POAG in NEIGHBORHOOD using a regression-based model. The results were particularly striking for high-tension POAG with an area under the receiver operating characteristic curve (AUC) of 76% [76]. This suggests that genetic markers, measurable at birth, have a substantial ability to predict later life IOP and risk of POAG, opening up the possibility of targeted population screening to aid earlier POAG detection and prevention of sight loss.
The potential for the UK Biobank IOP loci to aid detection of POAG was also examined in 1734 cases of advanced POAG and 2938 controls from the Australian and New Zealand Registry of Advanced Glaucoma (ANZRAG) [18]. MacGregor and colleagues derived a polygenic risk score based on their identified IOP loci and the CDKN2B-AS1 and SIX6 loci. Participants in the top decile of this score were at 5.6 (95% CI, 4.1–7.6) times increased odds of advanced POAG compared with participants in the bottom decile [18].
Genetic associations with central corneal thickness
While a thinner central corneal thickness (CCT) has been associated with increased POAG incidence [83], progression [84], and conversion from ocular hypertension [85], it remains uncertain whether the relationship is biologically causal, or whether it is driven by corneal artefact influencing IOP measurement. A landmark GWAS for CCT was reported in 2013 and identified 16 genome-wide significant loci; only one of these loci (at FNDC3B) was found to be associated with POAG and in an unexpected direction (the CCT-decreasing allele was protective for POAG) [86]. More recently, a larger GWAS of over 25,000 European and Asian participants identified 44 loci associated with CCT at genome-wide significance [87]. None of these loci were significantly associated with POAG (comparing 5008 cases with 35,472 controls) after correction for multiple testing [87]. Furthermore, there was no significant correlation between the CCT and POAG effect sizes for the CCT-associated variants (r = −0.17, P = 0.2). This is in contrast to the significant correlation identified between CCT and keratoconus effect sizes (r = −0.62, P = 5.3 × 10−5) [87]. Another GWAS of CCT and other corneal parameters was recently conducted in an Icelandic population; similarly, no significant CCT-associated loci were associated with glaucoma (either POAG or primary angle-closure glaucoma) [88]. Put together, the evidence suggests that CCT is not an endophenotype for POAG and supports the hypothesis that the CCT-glaucoma association observed in studies is due to IOP measurement artefact rather than biological causality.
Genetic associations with optic disc parameters
Initial glaucoma-related GWAS suggested that VCDR is a good endophenotype for glaucoma. The CDKN2B-AS1 locus was first reported in a VCDR GWAS [56] before being identified as POAG-associated subsequently [5]. Following this, large VCDR GWAS meta-analyses have identified over 50 associated loci, but only 9 of these were associated with POAG [75]. This suggests that some of the genetically driven variation in population VCDR is reflecting non-glaucomatous processes and may instead reflect baseline anatomy, for example. Similarly, many genome-wide significant loci have been identified for optic disc rim area and cup area, but the majority of these do not demonstrate significant association with POAG [75]. To date, there have been no reported genome-wide significant associations with circumpapillary retinal nerve-fibre layer thickness, though a recent study demonstrated a significant association between a known POAG-risk variant in SIX6 and retinal nerve-fibre layer thickness in a European adult population [89].
The road to personalized glaucoma management
Risk prediction and screening
Identifying glaucoma disease-related genes makes it possible to use disease associated or causative genetic variants to assess populations at risk. For glaucoma pre-symptomatic screening is particularly important, as patients are unaware of disease-related visual symptoms until later stages when the optic nerve is severely damaged, and treatment is not optimally effective. Current approaches for population screening involving IOP measurement, and/or optic nerve evaluation, are expensive and may only be effective for targeted screening of high-risk groups [90]. The discovery of genetic variants related to disease risk allows for the development of a gene-based screening approach that could identify patients at increased risk.
Substantial progress has been made toward gene-based screening. For patients with disease onset prior to age 50, disease-causing mutations in genes known to cause early-onset forms for glaucoma can be detected by DNA sequencing tests [91, 92]. Families and patients found to have a mutation in one of these genes can benefit from informed genetic counselling and treatment and surveillance plans tailored to individual disease risk [93]. A limitation is the relatively low diagnostic yield (20%) after testing for the genes currently known to cause early-onset glaucoma [92], suggesting that further work in this area would be fruitful.
Genetic screening to detect patients at high risk for adult-onset (after age 50) POAG also appears promising. Recent GWAS for POAG and related traits have successfully identified over 100 loci associated with disease risk have a receiver operator characteristic of up to 76%, suggesting that genetic risk factors can be an effective tool for discriminating POAG cases from controls [76]. More recently, genetic risk scores have demonstrated that cases with an excess of risk variants have earlier onset of disease [94] and greater risk of disease development [18] compared with individuals with fewer risk alleles. More comprehensive polygenic risk scores comprising larger numbers of POAG risk alleles may yield sufficiently accurate tests that screening based on genetics alone may be useful.
Gene-based therapies
Glaucoma gene discovery also makes possible the design of novel therapies that target the actual molecular events responsible for disease and recent advances in methods for gene-replacement and CRISPR/cas gene-editing support the feasibility of gene-based therapies for glaucoma in the relatively near future [95,96,97]. Currently, there are no FDA approved gene-based methods for glaucoma; however, several strategies have been suggested by recent research.
Genetic defects in MYOC, coding for Myocilin, are known to cause early-onset open-angle glaucoma inherited as a dominant trait. Disease-causing mutations are known to cause protein misfolding and cell toxicity due to endoplasmic reticulum accumulation [98]. Recent studies have shown that removing myocilin by CRISPR/cas gene-editing can lower IOP in transgenic mice carrying a deleterious MYOC mutation [97]. This very promising result suggests that CRISPR/cas could be used to target MYOC mutations in humans. Similarly, recent studies identifying TEK and ANGPT1 mutations in humans with early-onset glaucoma and due to abnormal Schlemm’s canal development and function suggest that restoring TEK signalling function could be therapeutic [80, 99]. As ANGPT1 has been associated with IOP and POAG in humans [15, 76], therapies targeting TEK signalling could be useful for both early-onset and adult-onset disease. Other early-onset glaucoma genes may also be targets for gene-based therapies including CYP1B1, known to cause recessive congenital and juvenile onset glaucoma [100], and PAX6, responsible for aniridia [101].
Genetic variants associated with POAG appear to impact a number of different biological processes and pathways and several of these could suggest effective therapeutic approaches. Four POAG and IOP loci include genes known to be involved in lipid metabolism (ABCA1, CAV1, DGKG, ARHGEF12) [12, 76]. Interestingly recent studies support protective effects of statin therapy on glaucoma [102]. Collectively, these results suggest that therapies targeting lipid and cholesterol metabolism could be effective treatment strategies, especially in patients with high tension glaucoma. Another potentially interesting therapeutic target is mitochondrial function. TXNRD2, thioredoxin reductase 2, codes for a mitochondrial protein required for reducing oxidative stress and maintaining redox homeostasis. TXNRD2 genomic variants have been associated with both POAG and IOP [13, 76], and in the UK Biobank IOP GWAS, four other genomic loci related to mitochondrial function were also associated with IOP (ME3, VPS13C, GCAT, PTCD2). These results suggest that mitochondrial function can contribute to IOP variation and that maintaining mitochondrial function could help regulate IOP and reduce POAG risk. Further discovery of POAG associated risk variants will identify additional targets for POAG gene-based therapies with effective and even curative potential.
References
Visscher PM, Wray NR, Zhang Q, Sklar P, McCarthy MI, Brown MA, et al. 10 years of GWAS discovery: biology, function, and translation. Am J Hum Genet. 2017;101:5–22.
MacArthur J, Bowler E, Cerezo M, Gil L, Hall P, Hastings E, et al. The new NHGRI-EBI catalog of published genome-wide association studies (GWAS Catalog). Nucleic Acids Res. 2017;45(D1):D896–D901.
Tam V, Patel N, Turcotte M, Bosse Y, Pare G, Meyre D. Benefits and limitations of genome-wide association studies. Nat Rev Genet. 2019;20:467–84.
Wiggs JL, Pasquale LR. Genetics of glaucoma. Hum Mol Genet. 2017;26(R1):R21–R7.
Burdon KP, Macgregor S, Hewitt AW, Sharma S, Chidlow G, Mills RA, et al. Genome-wide association study identifies susceptibility loci for open angle glaucoma at TMCO1 and CDKN2B-AS1. Nat Genet. 2011;43:574–8.
Thorleifsson G, Walters GB, Hewitt AW, Masson G, Helgason A, DeWan A, et al. Common variants near CAV1 and CAV2 are associated with primary open-angle glaucoma. Nat Genet. 2010;42:906–9.
Wiggs JL, Yaspan BL, Hauser MA, Kang JH, Allingham RR, Olson LM, et al. Common variants at 9p21 and 8q22 are associated with increased susceptibility to optic nerve degeneration in glaucoma. PLoS Genet. 2012;8:e1002654.
Osman W, Low SK, Takahashi A, Kubo M, Nakamura Y. A genome-wide association study in the Japanese population confirms 9p21 and 14q23 as susceptibility loci for primary open angle glaucoma. Hum Mol Genet. 2012;21:2836–42.
Gharahkhani P, Burdon KP, Fogarty R, Sharma S, Hewitt AW, Martin S, et al. Common variants near ABCA1, AFAP1 and GMDS confer risk of primary open-angle glaucoma. Nat Genet. 2014;46:1120–5.
Chen Y, Lin Y, Vithana EN, Jia L, Zuo X, Wong TY, et al. Common variants near ABCA1 and in PMM2 are associated with primary open-angle glaucoma. Nat Genet. 2014;46:1115–9.
Li Z, Allingham RR, Nakano M, Jia L, Chen Y, Ikeda Y, et al. A common variant near TGFBR3 is associated with primary open angle glaucoma. Hum Mol Genet. 2015;24:3880–92.
Springelkamp H, Iglesias AI, Cuellar-Partida G, Amin N, Burdon KP, van Leeuwen EM, et al. ARHGEF12 influences the risk of glaucoma by increasing intraocular pressure. Hum Mol Genet. 2015;24:2689–99.
Bailey JN, Loomis SJ, Kang JH, Allingham RR, Gharahkhani P, Khor CC, et al. Genome-wide association analysis identifies TXNRD2, ATXN2 and FOXC1 as susceptibility loci for primary open-angle glaucoma. Nat Genet. 2016;48:189–94.
Thorleifsson G, et al. Common variants near CAV1 and CAV2 are associated with primary open-angle glaucoma. Nat Genet. 2010;42:906–9.
Choquet H, Paylakhi S, Kneeland SC, Thai KK, Hoffmann TJ, Yin J, et al. A multiethnic genome-wide association study of primary open-angle glaucoma identifies novel risk loci. Nat Commun. 2018;9:2278.
Gharahkhani P, Burdon KP, Cooke Bailey JN, Hewitt AW, Law MH, Pasquale LR, et al. Analysis combining correlated glaucoma traits identifies five new risk loci for open-angle glaucoma. Sci Rep. 2018;8:3124.
Shiga Y, Akiyama M, Nishiguchi KM, Sato K, Shimozawa N, Takahashi A, et al. Genome-wide association study identifies seven novel susceptibility loci for primary open-angle glaucoma. Hum Mol Genet. 2018;27:1486–96.
MacGregor S, Ong JS, An J, Han X, Zhou T, Siggs OM, et al. Genome-wide association study of intraocular pressure uncovers new pathways to glaucoma. Nat Genet. 2018;50:1067–71.
Bycroft C, Freeman C, Petkova D, Band G, Elliott LT, Sharp K, et al. The UK Biobank resource with deep phenotyping and genomic data. Nature. 2018;562:203–9.
Sudlow C, Gallacher J, Allen N, Beral V, Burton P, Danesh J, et al. UK biobank: an open access resource for identifying the causes of a wide range of complex diseases of middle and old age. PLoS Med. 2015;12:e1001779.
Banda Y, Kvale MN, Hoffmann TJ, Hesselson SE, Ranatunga D, Tang H, et al. Characterizing race/ethnicity and genetic ancestry for 100,000 subjects in the Genetic Epidemiology Research on Adult Health and Aging (GERA) Cohort. Genetics. 2015;200:1285–95.
Kvale MN, Hesselson S, Hoffmann TJ, Cao Y, Chan D, Connell S, et al. Genotyping informatics and quality control for 100,000 subjects in the Genetic Epidemiology Research on Adult Health and Aging (GERA) Cohort. Genetics. 2015;200:1051–60.
Mishra A, Macgregor S. VEGAS2: software for more flexible gene-based testing. Twin Res Hum Genet. 2015;18:86–91.
Watanabe K, Taskesen E, van Bochoven A, Posthuma D. Functional mapping and annotation of genetic associations with FUMA. Nat Commun. 2017;8:1826.
Nakano M, Ikeda Y, Tokuda Y, Fuwa M, Omi N, Ueno M, et al. Common variants in CDKN2B-AS1 associated with optic-nerve vulnerability of glaucoma identified by genome-wide association studies in Japanese. PLoS One. 2012;7:e33389.
Hoffmann TJ, Tang H, Thornton TA, Caan B, Haan M, Millen AE, et al. Genome-wide association and admixture analysis of glaucoma in the women’s health initiative. Hum Mol Genet. 2014;23:6634–43.
Taylor KD, Guo X, Zangwill LM, Liebmann JM, Girkin CA, Feldman RM, et al. Genetic architecture of primary open-angle glaucoma in individuals of african descent: the African Descent and Glaucoma Evaluation Study III. Ophthalmology. 2019;126:38–48.
Bonnemaijer PWM, Iglesias AI, Nadkarni GN, Sanyiwa AJ, Hassan HG, Cook C, et al. Genome-wide association study of primary open-angle glaucoma in continental and admixed African populations. Hum Genet. 2018;137:847–62.
Khachatryan N, Medeiros FA, Sharpsten L, Bowd C, Sample PA, Liebmann JM, et al. The African Descent and Glaucoma Evaluation Study (ADAGES): predictors of visual field damage in glaucoma suspects. Am J Ophthalmol. 2015;159:777–87.
Kyari F, Abdull MM, Bastawrous A, Gilbert CE, Faal H. Epidemiology of glaucoma in sub-saharan Africa: prevalence, incidence and risk factors. Middle East Afr J Ophthalmol. 2013;20:111–25.
Williams SE, Carmichael TR, Allingham RR, Hauser M, Ramsay M. The genetics of POAG in black South Africans: a candidate gene association study. Sci Rep. 2015;5:8378.
Cao D, Jiao X, Liu X, Hennis A, Leske MC, Nemesure B, et al. CDKN2B polymorphism is associated with primary open-angle glaucoma (POAG) in the Afro-Caribbean population of Barbados, West Indies. PLoS One. 2012;7:e39278.
Liu Y, Hauser MA, Akafo SK, Qin X, Miura S, Gibson JR, et al. Investigation of known genetic risk factors for primary open angle glaucoma in two populations of African ancestry. Invest Ophthalmol Vis Sci. 2013;54:6248–54.
Restrepo NA, Laper SM, Farber-Eger E, Crawford DC. Local genetic ancestry in CDKN2B-AS1 is associated with primary open-angle glaucoma in an African American cohort extracted from de-identified electronic health records. BMC Med Genomics. 2018;11 Suppl 3 :70.
Carnes MU, Liu YP, Allingham RR, Whigham BT, Havens S, Garrett ME, et al. Discovery and functional annotation of SIX6 variants in primary open-angle glaucoma. PLoS Genet. 2014;10:e1004372.
Gao S, Jakobs TC. Mice homozygous for a deletion in the glaucoma susceptibility locus ink4 show increased vulnerability of retinal ganglion cells to elevated intraocular pressure. Am J Pathol. 2016;186:985–1005.
Skowronska-Krawczyk D, Zhao L, Zhu J, Weinreb RN, Cao G, Luo J, et al. P16INK4a upregulation mediated by six6 defines retinal ganglion cell pathogenesis in glaucoma. Mol Cell. 2015;59:931–40.
Elliott MH, Ashpole NE, Gu X, Herrnberger L, McClellan ME, Griffith GL, et al. Caveolin-1 modulates intraocular pressure: implications for caveolae mechanoprotection in glaucoma. Sci Rep. 2016;6:37127.
Kawakami K, Sato S, Ozaki H, Ikeda K. Six family genes-structure and function as transcription factors and their roles in development. Bioessays. 2000;22:616–26.
Kumar JP. The sine oculis homeobox (SIX) family of transcription factors as regulators of development and disease. Cell Mol Life Sci. 2009;66:565–83.
Iglesias AI, Springelkamp H, van der Linde H, Severijnen LA, Amin N, Oostra B, et al. Exome sequencing and functional analyses suggest that SIX6 is a gene involved in an altered proliferation-differentiation balance early in life and optic nerve degeneration at old age. Hum Mol Genet. 2014;23:1320–32.
Bosco A, Steele MR, Vetter ML. Early microglia activation in a mouse model of chronic glaucoma. J Comp Neurol. 2011;519:599–620.
Bosco A, Romero CO, Breen KT, Chagovetz AA, Steele MR, Ambati BK, et al. Neurodegeneration severity can be predicted from early microglia alterations monitored in vivo in a mouse model of chronic glaucoma. Dis Model Mech. 2015;8:443–55.
Choquet H, Thai KK, Yin J, Hoffmann TJ, Kvale MN, Banda Y, et al. A large multi-ethnic genome-wide association study identifies novel genetic loci for intraocular pressure. Nat Commun. 2017;8:2108.
Mimiwati Z, Mackey DA, Craig JE, Mackinnon JR, Rait JL, Liebelt JE, et al. Nail-patella syndrome and its association with glaucoma: a review of eight families. Br J Ophthalmol. 2006;90:1505–9.
Sweeney E, Fryer A, Mountford R, Green A, McIntosh I. Nail patella syndrome: a review of the phenotype aided by developmental biology. J Med Genet. 2003;40:153–62.
Lichter PR, Richards JE, Downs CA, Stringham HM, Boehnke M, Farley FA. Cosegregation of open-angle glaucoma and the nail-patella syndrome. Am J Ophthalmol. 1997;124:506–15.
Gamazon ER, Wheeler HE, Shah KP, Mozaffari SV, Aquino-Michaels K, Carroll RJ, et al. A gene-based association method for mapping traits using reference transcriptome data. Nat Genet. 2015;47:1091–8.
Gusev A, Ko A, Shi H, Bhatia G, Chung W, Penninx BW, et al. Integrative approaches for large-scale transcriptome-wide association studies. Nat Genet. 2016;48:245–52.
Zhu Z, Zhang F, Hu H, Bakshi A, Robinson MR, Powell JE, et al. Integration of summary data from GWAS and eQTL studies predicts complex trait gene targets. Nat Genet. 2016;48:481–7.
Pers TH, Karjalainen JM, Chan Y, Westra HJ, Wood AR, Yang J, et al. Biological interpretation of genome-wide association studies using predicted gene functions. Nat Commun. 2015;6:5890.
Consortium EP. An integrated encyclopedia of DNA elements in the human genome. Nature. 2012;489:57–74.
Roadmap Epigenomics C, Kundaje A, Meuleman W, Ernst J, Bilenky M, Yen A, et al. Integrative analysis of 111 reference human epigenomes. Nature. 2015;518:317–30.
Consortium GT. The Genotype-Tissue Expression (GTEx) project. Nat Genet. 2013;45:580–5.
Hysi PG, Cheng CY, Springelkamp H, Macgregor S, Bailey JNC, Wojciechowski R, et al. Genome-wide analysis of multi-ancestry cohorts identifies new loci influencing intraocular pressure and susceptibility to glaucoma. Nat Genet. 2014;46:1126–30.
Ramdas WD, van Koolwijk LM, Ikram MK, Jansonius NM, de Jong PT, Bergen AA, et al. A genome-wide association study of optic disc parameters. PLoS Genet. 2010;6:e1000978.
Thorleifsson G, Magnusson KP, Sulem P, Walters GB, Gudbjartsson DF, Stefansson H, et al. Common sequence variants in the LOXL1 gene confer susceptibility to exfoliation glaucoma. Science. 2007;317:1397–400.
Aung T, Ozaki M, Lee MC, Schlotzer-Schrehardt U, Thorleifsson G, Mizoguchi T, et al. Genetic association study of exfoliation syndrome identifies a protective rare variant at LOXL1 and five new susceptibility loci. Nat Genet. 2017;49:993–1004.
Pasutto F, Zenkel M, Hoja U, Berner D, Uebe S, Ferrazzi F, et al. Pseudoexfoliation syndrome-associated genetic variants affect transcription factor binding and alternative splicing of LOXL1. Nat Commun. 2017;8:15466.
Verhoeven VJ, Hysi PG, Wojciechowski R, Fan Q, Guggenheim JA, Hohn R, et al. Genome-wide meta-analyses of multiancestry cohorts identify multiple new susceptibility loci for refractive error and myopia. Nat Genet. 2013;45:314–8.
Klein RJ, Zeiss C, Chew EY, Tsai JY, Sackler RS, Haynes C, et al. Complement factor H polymorphism in age-related macular degeneration. Science. 2005;308:385–9.
Seddon JM, Yu Y, Miller EC, Reynolds R, Tan PL, Gowrisankar S, et al. Rare variants in CFI, C3 and C9 are associated with high risk of advanced age-related macular degeneration. Nat Genet. 2013;45:1366–70.
Fritsche LG, Igl W, Bailey JN, Grassmann F, Sengupta S, Bragg-Gresham JL, et al. A large genome-wide association study of age-related macular degeneration highlights contributions of rare and common variants. Nat Genet. 2016;48:134–43.
Cuellar-Partida G, Craig JE, Burdon KP, Wang JJ, Vote BJ, Souzeau E, et al. Assessment of polygenic effects links primary open-angle glaucoma and age-related macular degeneration. Sci Rep. 2016;6:26885.
Bulik-Sullivan B, Finucane HK, Anttila V, Gusev A, Day FR, Loh PR, et al. An atlas of genetic correlations across human diseases and traits. Nat Genet. 2015;47:1236–41.
Pickrell JK, Berisa T, Liu JZ, Segurel L, Tung JY, Hinds DA. Detection and interpretation of shared genetic influences on 42 human traits. Nat Genet. 2016;48:709–17.
Sivakumaran S, Agakov F, Theodoratou E, Prendergast JG, Zgaga L, Manolio T, et al. Abundant pleiotropy in human complex diseases and traits. Am J Hum Genet. 2011;89:607–18.
Ge T, Chen CY, Neale BM, Sabuncu MR, Smoller JW. Phenome-wide heritability analysis of the UK Biobank. PLoS Genet .2017;13:e1006711.
Bakshi A, Zhu Z, Vinkhuyzen AA, Hill WD, McRae AF, Visscher PM, et al. Fast set-based association analysis using summary data from GWAS identifies novel gene loci for human complex traits. Sci Rep. 2016;6:32894.
Timpson NJ, Greenwood CMT, Soranzo N, Lawson DJ, Richards JB. Genetic architecture: the shape of the genetic contribution to human traits and disease. Nat Rev Genet. 2018;19:110–24.
Zhou T, Souzeau E, Sharma S, Landers J, Mills R, Goldberg I, et al. Whole exome sequencing implicates eye development, the unfolded protein response and plasma membrane homeostasis in primary open-angle glaucoma. PLoS One. 2017;12:e0172427.
de Voogd S, Ikram MK, Wolfs RC, Jansonius NM, Hofman A, de Jong PT. Incidence of open-angle glaucoma in a general elderly population: the Rotterdam Study. Ophthalmology. 2005;112:1487–93.
Garway-Heath DF, Crabb DP, Bunce C, Lascaratos G, Amalfitano F, Anand N, et al. Latanoprost for open-angle glaucoma (UKGTS): a randomised, multicentre, placebo-controlled trial. Lancet. 2015;385:1295–304.
Sanfilippo PG, Hewitt AW, Hammond CJ, Mackey DA. The heritability of ocular traits. Surv Ophthalmol. 2010;55:561–83.
Springelkamp H, Iglesias AI, Mishra A, Hohn R, Wojciechowski R, Khawaja AP, et al. New insights into the genetics of primary open-angle glaucoma based on meta-analyses of intraocular pressure and optic disc characteristics. Hum Mol Genet. 2017;26:438–53.
Khawaja AP, Cooke Bailey JN, Wareham NJ, Scott RA, Simcoe M, Igo RP Jr., et al. Genome-wide analyses identify 68 new loci associated with intraocular pressure and improve risk prediction for primary open-angle glaucoma. Nat Genet. 2018;50:778–82.
Khawaja AP, Chan MP, Hayat S, Broadway DC, Luben R, Garway-Heath DF, et al. The EPIC-Norfolk Eye Study: rationale, methods and a cross-sectional analysis of visual impairment in a population-based cohort. BMJ Open. 2013;3:e002684. pii.
Agarwal R, Agarwal P. Newer targets for modulation of intraocular pressure: focus on adenosine receptor signaling pathways. Expert Opin Ther Targets. 2014;18:527–39.
Aspelund A, Tammela T, Antila S, Nurmi H, Leppanen VM, Zarkada G, et al. The Schlemm’s canal is a VEGF-C/VEGFR-3-responsive lymphatic-like vessel. J Clin Invest. 2014;124:3975–86.
Souma T, Tompson SW, Thomson BR, Siggs OM, Kizhatil K, Yamaguchi S, et al. Angiopoietin receptor TEK mutations underlie primary congenital glaucoma with variable expressivity. J Clin Invest. 2016;126:2575–87.
Kizhatil K, Ryan M, Marchant JK, Henrich S, John SW. Schlemm’s canal is a unique vessel with a combination of blood vascular and lymphatic phenotypes that forms by a novel developmental process. PLoS Biol. 2014;12:e1001912.
Thomson BR, Heinen S, Jeansson M, Ghosh AK, Fatima A, Sung HK, et al. A lymphatic defect causes ocular hypertension and glaucoma in mice. J Clin Invest. 2014;124:4320–4.
Jiang X, Varma R, Wu S, Torres M, Azen SP, Francis BA, et al. Baseline risk factors that predict the development of open-angle glaucoma in a population: the Los Angeles Latino Eye Study. Ophthalmology. 2012;119:2245–53.
Leske MC, Heijl A, Hyman L, Bengtsson B, Dong L, Yang Z, et al. Predictors of long-term progression in the early manifest glaucoma trial. Ophthalmology. 2007;114:1965–72.
Gordon MO, Beiser JA, Brandt JD, Heuer DK, Higginbotham EJ, Johnson CA, et al. The Ocular Hypertension Treatment Study: baseline factors that predict the onset of primary open-angle glaucoma. Arch Ophthalmol. 2002;120:714–20. discussion829-30.
Lu Y, Vitart V, Burdon KP, Khor CC, Bykhovskaya Y, Mirshahi A, et al. Genome-wide association analyses identify multiple loci associated with central corneal thickness and keratoconus. Nat Genet 2013;45:155–63.
Iglesias AI, Mishra A, Vitart V, Bykhovskaya Y, Hohn R, Springelkamp H, et al. Cross-ancestry genome-wide association analysis of corneal thickness strengthens link between complex and Mendelian eye diseases. Nat Commun 2018;9:1864.
Ivarsdottir EV, Benonisdottir S, Thorleifsson G, Sulem P, Oddsson A, Styrkarsdottir U, et al. Sequence variation at ANAPC1 accounts for 24% of the variability in corneal endothelial cell density. Nat Commun 2019;10:1284.
Khawaja AP, Chan MPY, Yip JLY, Broadway DC, Garway-Heath DF, Viswanathan AC, et al. A common glaucoma-risk variant of six6 alters retinal nerve fiber layer and optic disc measures in a european population: the EPIC-Norfolk Eye Study. J Glaucoma 2018;27:743–9.
Burr JM, Mowatt G, Hernandez R, Siddiqui MA, Cook J, Lourenco T, et al. The clinical effectiveness and cost-effectiveness of screening for open angle glaucoma: a systematic review and economic evaluation. Health Technol Assess 2007;11:1–190. iii-iv, ix-x.
Souzeau E, Tram KH, Witney M, Ruddle JB, Graham SL, Healey PR, et al. Myocilin predictive genetic testing for primary open-angle glaucoma leads to early identification of at-risk individuals. Ophthalmology 2017;124:303–9.
Allen KF, Gaier ED, Wiggs JL. Genetics of primary inherited disorders of the optic nerve: clinical applications. Cold Spring Harb Perspect Med 2015;5:a017277.
Wiggs JL, Pierce EA. Genetic testing for inherited eye disease: who benefits? JAMA Ophthalmol 2013;131:1265–6.
Fan BJ, Bailey JC, Igo RP, Jr., Kang JH, Boumenna T, Brilliant MH, et al. Association of a primary open-angle glaucoma genetic risk score with earlier age at diagnosis. JAMA Ophthalmol. 2019;137:1190–4.
Borras T. The pathway from genes to gene therapy in glaucoma: a review of possibilities for using genes as glaucoma drugs. Asia Pac J Ophthalmol (Philos) 2017;6:80–93.
O’Callaghan J, Crosbie DE, Cassidy PS, Sherwood JM, Flugel-Koch C, Lutjen-Drecoll E, et al. Therapeutic potential of AAV-mediated MMP-3 secretion from corneal endothelium in treating glaucoma. Hum Mol Genet. 2017;26:1230–46.
Jain A, Zode G, Kasetti RB, Ran FA, Yan W, Sharma TP, et al. CRISPR-Cas9-based treatment of myocilin-associated glaucoma. Proc Natl Acad Sci USA 2017;114:11199–204.
Kwon YH, Fingert JH, Kuehn MH, Alward WL. Primary open-angle glaucoma. N Engl J Med 2009;360:1113–24.
Thomson BR, Souma T, Tompson SW, Onay T, Kizhatil K, Siggs OM, et al. Angiopoietin-1 is required for Schlemm’s canal development in mice and humans. J Clin Invest 2017;127:4421–36.
Wiggs JL, Langgurth AM, Allen KF. Carrier frequency of CYP1B1 mutations in the United States (an American Ophthalmological Society thesis). Trans Am Ophthalmol Soc 2014;112:94–102.
Yongblah K, Alford SC, Ryan BC, Chow RL, Howard PL. Protecting Pax6 3’ UTR from microrna-7 partially restores pax6 in islets from an aniridia mouse model. Mol Ther Nucleic Acids 2018;13:144–53.
Kang JH, Boumenna T, Stein JD, Khawaja A, Rosner BA, Wiggs JL, et al. Association of statin use and high serum cholesterol levels with risk of primary open-angle glaucoma. JAMA Ophthalmol 2019;137:756–65.
Acknowledgements
HC is supported by a National Eye Institute (NEI) grant R01 EY027004 and a National Institute of Diabetes and Digestive and Kidney Diseases grant R01 DK116738. JLW is supported by the two following NIH/NEI grants: R01 EY022305 and P30 EY014104. APK is supported by a Moorfields Eye Charity Career Development Fellowship.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Choquet, H., Wiggs, J.L. & Khawaja, A.P. Clinical implications of recent advances in primary open-angle glaucoma genetics. Eye 34, 29–39 (2020). https://doi.org/10.1038/s41433-019-0632-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41433-019-0632-7
This article is cited by
-
Population screening for glaucoma in UK: current recommendations and future directions
Eye (2022)
-
Solving neurodegeneration: common mechanisms and strategies for new treatments
Molecular Neurodegeneration (2022)
-
A Comparison of Genomic Advances in Exfoliation Syndrome and Primary Open-Angle Glaucoma
Current Ophthalmology Reports (2021)
-
GLIS1 regulates trabecular meshwork function and intraocular pressure and is associated with glaucoma in humans
Nature Communications (2021)
-
Genome-wide meta-analysis identifies 127 open-angle glaucoma loci with consistent effect across ancestries
Nature Communications (2021)
