
Abstract
Accumulating evidence indicates that the pluripotency of periodontal ligament stem cells (PDLSCs) is compromised under inflammatory conditions; however, the underlying mechanisms remain largely unexplored. In this study, we hypothesize that the P2X7 receptor (P2X7R) is a key molecule linked to inflammation-associated impairment of PDLSCs. We first investigated P2X7R expression in PDLSCs under normal and inflammatory conditions and then determined the effect of a P2X7R agonist (BzATP) or antagonist (BBG) on PDLSC osteogenesis under various conditions. Gene-modified PDLSCs were used to further examine the role of P2X7R and the signaling pathway underlying P2X7R-enhanced osteogenesis. We found that inflammatory conditions decreased P2X7R expression in PDLSCs and reduced osteogenesis in these cells. In addition, activation of P2X7R by BzATP or overexpression of P2X7R via gene transduction reversed the inflammation-mediated decrease in PDLSC osteogenic differentiation. When selected osteogenesis-related signaling molecules were screened, the PI3K-AKT-mTOR pathway was identified as potentially involved in P2X7R-enhanced PDLSC osteogenesis. Our data reveal a crucial role for P2X7R in PDLSC osteogenesis under inflammatory conditions, suggesting a new therapeutic target to reverse or rescue inflammation-mediated changes in PDLSCs for future mainstream therapeutic uses.
Similar content being viewed by others


Introduction
Periodontitis is one of the most common infectious diseases, and the resulting destruction of tooth-supporting tissues is the main reason for tooth loss in adults1. In addition, periodontitis accelerates the initiation and/or progression of other systemic diseases in distant organs, including obesity, cardiovascular disease, diabetes, and chronic nephritis2. Given the crucial role of the periodontium, the aim of periodontal therapy is not only to prevent infection and hence retard/prevent disease progression but also to regenerate damaged periodontal tissue and/or reconstruct lost tooth-supporting structures, including the alveolar bone, the cementum, and the periodontal ligament (PDL). Thus, stem cell-based approaches (with or without various biomaterials and/or growth factors) have been developed in an attempt to regenerate lost/damaged periodontium composed of different tissues and to achieve improved clinical outcomes3. Based on the preclinical use of mesenchymal stem cells (MSCs) in animal models, PDL-derived stem cells (PDLSCs) represent one of the most reliable sources due to their potential to differentiate into bone-, PDL-, and cementum-forming lineages4. Other cell sources, such as bone marrow-derived MSCs (BMMSCs) and adipose tissue-derived MSCs (ASCs), have also undergone initial testing for periodontal use5. Although the use of culture-expanded stem cells, including but not limited to PDLSCs, for periodontal regeneration has resulted in significant beneficial effects in various animal models, the clinical experience with stem cell-based therapies in periodontics has thus far been disappointing6. Overall, the unsuccessful translation of findings from the preclinical stage to the clinical stage has largely been attributed to changes in the regenerative potential of stem cells in various in vivo situations7,8. Indeed, despite the robust functionality exhibited by stem cells in artificially created bone defects in animal models, predictable tissue regeneration is not achieved when they are applied to treat periodontitis-caused tissue loss, at least in part because these cells may be functionally compromised in an inflammatory environment6.
In fact, there is substantial evidence that PDLSCs lose some degree of their pluripotency in an inflammatory microenvironment, leading to impaired immunomodulation and regenerative potential7,8,9,10. However, the underlying pathways remain largely unknown. Recent data suggest that nuclear factor-κB (NF-κB)11,12, GSK3β13,14, DKK115, general control nonrepressed protein 5 (GCN5)16, and MORF (i.e., lysine acetyltransferase 6B)17 are involved in the impaired osteogenic potential of PDLSCs, and several molecular pathways such as the Wnt11,12,14, mitogen-activated protein kinase (MAPK) and BMP/Smad pathways18 may also be involved in inflammation-associated changes in PDLSC osteogenesis. Regardless, current data cannot provide a complete understanding of phenomena observed with in vitro and in vivo models. Hence, an effective strategy for modulating PDLSCs in a chronic inflammatory environment toward correct differentiation and proper function has thus far been an unrealized goal19. Clearly, the development of clinical therapeutics for use in patients requires effective methods to rescue inflammation-mediated changes of PDLSCs20. Further examination of PDLSC behavior under normal and inflammatory conditions and elucidation of inflammation-related signaling mechanisms governing PDLSC osteogenesis may offer new insight into periodontitis, thereby assisting in the development of improved methods and more effective therapeutic protocols to advance stem cell-based periodontal regeneration 21.
Recently, increasing evidence indicates that as an ATP-gated ion channel expressed in most stem cells, such as BMMSCs22, embryonic stem cells (ESCs), hematopoietic stem/progenitor cells23, neural precursor cells24, and PDLSCs25,26,27, the P2X7 receptor (P2X7R) plays wide-ranging physiological and pathological roles in inflammation, some inflammatory-related neurological disorders, cancer and metabolic disease28. This is also true in the progression of periodontitis. For example, extracellular ATP has been reported to be a key modulator of inflammation, and ATP signaling via P2X7R can initiate the proinflammatory cascade in pathological alveolar bone loss in periodontitis27 as well as influence the homeostasis and function of the periodontium25,26. Similarly, P2X7R expression on BMMSCs has been found to regulate the formation and apoptosis of osteoblasts/osteoclasts at different stages to orchestrate bone metabolism29,30. In this context, P2X7R has a significant positive influence on osteogenesis29,31 and bone formation32,33,34. Moreover, specific P2X7R activation has been shown to impair bone mineralization and alkaline phosphatase (ALP) activation in neonatal rats35. Further evidence shows that the P2X7R agonist BzATP may induce the apoptosis of SaOS-2 (human osteosarcoma cell line) osteoblast36. Although the important role of P2X7R in determining stem cell fate and function has been demonstrated in recent years, P2X7R expression in human PDLSCs in response to disease conditions such as an inflammatory microenvironment is largely unknown. In this study, P2X7R expression in PDLSCs under normal and inflammatory conditions and the effects of 2,3(4-benzoylbenzoyl)-ATP (BzATP, a P2X7R agonist) or Brilliant Blue G (BBG, a P2X7R antagonist) on PDLSC osteogenesis under various conditions were investigated in a coordinated fashion. Gene-modified PDLSCs were employed to further assess the role of P2X7R and the signaling pathway underlying P2X7R-enhanced osteogenesis. Our data will clarify whether and how P2X7R is linked to stem cell function under inflammatory conditions, and the findings may provide a new target for rescuing inflammation-mediated impairment of PDLSC osteogenesis to meet the need for novel therapeutics for periodontitis.
Results
Characterization of human PDLSCs
Primary cells were successfully obtained from PDL tissues from nine donors (aged 27.5 ± 9.5) (Supplementary Fig. 1A), and the subpopulation of these PDL cells possessed the ability to generate new colonies (Supplementary Fig. 1B). Furthermore, these cells continued to multiply, as evidenced by the CCK-8 assay (Supplementary Fig. 1C). The cells were also able to differentiate under osteo-inductive, adipo-inductive, or chondrogenic inductive conditions, producing either mineralized extracellular matrices positively stained with Alizarin red (parts of cells yielded bone trabecular-like structures), oil globules with abundant lipid-rich vacuoles in the cytoplasm or acidic polysaccharides extracellular matrices positively stained with Alcian blue (Supplementary Fig. 1D). Flow cytometric analysis revealed that these colony-forming cells positively expressed MSC markers, such as CD90, CD105, and CD146, but were negative for endothelial cell (CD31) and hematopoiesis-related (CD34 and CD45) markers (Supplementary Fig. 1E).
P2X7R expression in PDLSCs and osteogenesis under inflammatory conditions
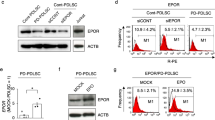
Under inflammatory culture conditions (medium supplemented with TNF-α and IL-1β), the level of P2X7R mRNA in PDLSCs was significantly decreased compared with that under normal conditions (p < 0.05). Incubation of PDLSCs in the osteo-inductive microenvironment led to an increase in P2X7R expression under normal conditions (p < 0.01), but this increase was largely abolished by the addition of inflammatory factors (p < 0.001) (Fig. 1a). The observed decrease in P2X7R protein expression in PDLSCs in response to inflammatory factors (p < 0.001) and increase in P2X7R expression under osteo-inductive conditions were confirmed by western blotting (p < 0.001). Although P2X7R protein expression in PDLSCs also decreased upon the addition of inflammatory factors under osteogenic conditions, it was not statistically significant (Fig. 1b).

a Quantitative analysis of P2X7R mRNA expression in PDLSCs in normal α-MEM culture medium, inflammatory culture medium (medium supplemented with TNF-α and IL-1β), osteo-inductive culture medium, and inflammatory osteo-inductive culture medium (osteo-inductive culture medium supplemented with TNF-α and IL-1β). b Western blotting of P2X7R in PDLSCs in normal α-MEM culture medium, inflammatory culture medium (medium supplemented with TNF-α and IL-1β), osteo-inductive culture medium, and inflammatory osteo-inductive culture medium (osteo-inductive culture medium supplemented with TNF-α and IL-1β). Data are presented as the mean ± S.D. for n from 4 to 6; ns indicates no significant difference between the indicated columns; *p < 0.05, **p < 0.01, and ***p < 0.001 indicate significant differences between the indicated columns
To ascertain the effect of P2X7R expression on osteogenesis, we observed osteogenic differentiation (in terms of ALP staining, Alizarin red staining, quantitative real-time polymerase chain reaction (qRT-PCR), and western blotting) in response to a specific P2X7R antagonist (BBG) or agonist (BzATP) under normal or inflammatory conditions. Following osteogenic induction for 7 or 21 days, the PDLSCs formed mineralized nodules that were positively stained by ALP or Alizarin red staining (Fig. 2a, b). However, the ability of the cells to undergo osteogenic differentiation was largely impaired by the addition of either inflammatory factors or the P2X7R antagonist (BBG), whereas the P2X7R agonist (BzATP) was able to rescue the inflammation-induced reduction in osteogenesis. According to qRT-PCR analysis, the PDLSCs expressed osteoblast differentiation-related transcription factors such as Runx2 and OCN under osteo-inductive conditions, but both Runx2 and OCN gene levels were significantly decreased in response to inflammatory factors (p < 0.001) or the P2X7R antagonist (p < 0.01). Conversely, BzATP treatment led to a significant increase in Runx2 and OCN mRNA levels under inflammatory conditions (p < 0.001; Fig. 2c, d). Similarly, western blot analysis confirmed that Runx2 and OCN expression was significantly decreased under inflammatory conditions as well as under osteo-inductive conditions with P2X7R antagonist addition (p < 0.001; Fig. 2e, f). In contrast, Runx2 and OCN protein expression under inflammatory osteo-inductive conditions was increased by BzATP-induced activation of P2X7R (p < 0.001 or p < 0.01; Fig. 2e, f). To further confirm the BzATP-induced effects were P2X7 specific, P2X7-selective antagonist A740003 was used to inhibit the osteogenesis of PDLSCs under inflammatory osteo-inductive conditions (Supplementary Fig. 2).

a ALP staining of PDLSCs under 7-day osteogenic culture conditions with or without BBG/BzATP addition (scale bar = 1 mm). b Alizarin red staining of PDLSCs under 21-day osteogenic culture conditions with or without BBG/BzATP addition (scale bar = 500 μm). c Runx2 and d OCN mRNA expression (determined by qRT-PCR) in PDLSCs following a 7-day culture. e Runx2 and f OCN protein expression (determined by western blotting) in PDLSCs following a 7-day culture. Data are presented as the mean ± S.D. for n = 4; **p < 0.01 and ***p < 0.001 indicate significant differences between the indicated columns
Osteogenesis of P2X7R gene-modified PDLSCs under inflammatory conditions
To further investigate P2X7R-regulated osteogenesis under inflammatory conditions, we constructed an adenovirus vector to stably upregulate P2X7R in PDLSCs. As transduction of PDLSCs using an adenovirus vector carrying the P2X7R gene (Ad-P2X7) at 50 pfu/cell (multiplicity of infection (MOI) = 50) or 100 pfu/cell (MOI = 100) resulted in significant cell death, we excluded these two concentrations in further experiments. qRT-PCR results showed that P2X7R gene expression in Ad-P2X7 groups increased 10-fold at MOI = 10 and 23-fold at MOI = 20 when compared with the Ad-control group (Fig. 3a). In addition, a western blot assay revealed that PDLSC transduction with Ad-P2X7 at MOI = 10 or MOI = 20 resulted in a 4-fold or 10-fold increase, respectively, in P2X7R protein expression compared with Ad-control transduction (Fig. 3b). Hence, PDLSCs modified by Ad-P2X7 at 20 pfu/cell were chosen for subsequent experiments. Immunofluorescence staining demonstrated that the P2X7R gene was successfully expressed in PDLSCs after transduction with Ad-P2X7 at 20 pfu/cell (Fig. 3d). Based on CCK-8 assay results, these gene-modified cells exhibited enhanced proliferation capacity compared with Ad-control-modified cells, particularly at days 5 and 6 (p < 0.05; Fig. 3c).

a P2X7R mRNA expression in PDLSCs (assessed by qRT-PCR) in response to transduction with Ad-control or Ad-P2X7 at 10 pfu/cell (MOI = 10) or 20 pfu/cell (MOI = 20); cells without transduction were used as the control group. b P2X7R protein expression in PDLSCs (assessed by western blotting) in response to transduction with Ad-control or Ad-P2X7 at 10 pfu/cell (MOI = 10) or 20 pfu/cell (MOI = 20); cells without transduction were used as the control group. c Proliferative activity of Ad-P2X7-transfected PDLSCs (MOI = 20) and Ad-control-transfected PDLSCs assessed by the CCK-8 assay. d Representative images (confocal micrograph) of PDLSCs transfected with Ad-P2X7 or Ad-control (MOI = 20), which carries the green fluorescence protein (GFP) gene and P2X7R (immunofluorescence staining with a P2X7R antibody, red) (scale bar = 20 μm). Data are presented as the mean ± S.D. for n = 3; ns indicates no significant difference between the indicated columns; *p < 0.05, **p < 0.01, and ***p < 0.001 indicate significant differences between the indicated columns
To determine the effect of P2X7R gene transduction on osteogenesis under inflammatory conditions, we evaluated the osteogenesis of gene-modified cells in response to exposure to inflammatory factors or a P2X7R antagonist (BBG). Gene-modified PDLSCs exhibited robust osteogenic differentiation, irrespective of the addition of inflammatory factors, but their osteogenic potential was impaired when BBG was added (p < 0.001) (Fig. 4a, b). In western blot assays, gene-modified PDLSCs showed higher levels of Runx2 and OCN expression, even under inflammatory conditions, though inflammation did slightly reduce the level of OCN in gene-modified cells (p < 0.05). This enhanced Runx2 and OCN expression in gene-modified PDLSCs was blocked by the addition of BBG under osteo-inductive conditions (p < 0.001 or 0.01) (Fig. 4c, d).

a ALP staining (scale bar = 1 mm) of Ad-control-transfected or Ad-P2X7-transfected PDLSCs (left) and the quantitative result (right). b Alizarin red staining (scale bar = 500 μm) of Ad-control-transfected or Ad-P2X7-transfected PDLSCs (left) and the quantitative result (right). Western blotting analysis of c Runx2 and d OCN protein expression in Ad-control-transfected or Ad-P2X7-transfected PDLSCs. Data are presented as the mean ± S.D. for n = 4; ns indicates no significant difference between the indicated columns; *p < 0.05, **p < 0.01, and ***p < 0.001 indicate significant differences between the indicated columns
PI3K-AKT-mTOR signaling is involved in P2X7R-induced osteogenesis
Target gene screening: We further explored the possible mechanism involved in the osteogenesis of gene-modified PDLSCs. Given that the MAPK, Wnt/β-catenin and PI3K-AKT families have been demonstrated to participate in osteogenesis29, we used qRT-PCR to screen for molecules from these three families putatively involved in P2X7R-enhanced osteogenesis (AP-1, FOS-B, Wnt-3α, GSK-3β, CTNNB, FOX-O, NF-κB, and mTOR). Relatively stable expression of the Wnt-3α gene was observed in gene-modified PDLSCs, regardless of the osteo-inductive conditions (inflammatory factors with or without BzATP and normal with or without BBG), and expression of three additional genes (AP-1, GSK-3β, and CTNNB) was relatively stable, irrespective of the addition of inflammatory factors (Fig. 5a). Therefore, these four genes were excluded as potential mediators of P2X7R-enhanced PDLSC osteogenesis under inflammatory conditions. The other four genes (FOS-B, FOX-O, NFκB, and mTOR) displaying visible changes in response to inflammatory factors and the addition of BzATP/BBG were subjected to quantitative analysis. As no significant reductions in FOS-B, FOX-O, and NFκB gene-expression levels were found when BBG was added to cells (p > 0.05) (Fig. 5b–d), these genes were also excluded.

a Heat maps of osteogenesis-related genes that may be involved in the observed enhanced osteogenesis of Ad-P2X7-transfected PDLSCs. Quantitative analysis of b mTOR, c NFκB, and d FOX-O mRNA expression in Ad-P2X7-transfected PDLSCs under normal or inflammatory conditions with or without the addition of P2X7 agonist (BzATP) or P2X7 antagonist (BBG). Data are presented as the mean ± S.D. for n = 3; ns indicates no significant difference between the indicated columns; **p < 0.01 and ***p < 0.001 indicate significant differences between the indicated columns
Inflammation significantly reduced mTOR gene expression (p < 0.01), whereas BzATP addition abolished this reduction (p < 0.01). In addition, the presence of BBG significantly inhibited mTOR gene expression (p < 0.01) (Fig. 5e). Therefore, we chose mTOR and its upstream signaling molecules PI3K and AKT as target genes for further investigation to elucidate the possible mechanism by which PDLSC osteogenesis is enhanced by P2X7R.
Phosphorylation in the PI3K-AKT-mTOR signaling pathway: Western blot analysis was applied to examine PI3K-AKT-mTOR pathway components potentially involved in P2X7R-enhanced PDLSC osteogenesis. Ad-P2X7-transduced PDLSCs possessed higher levels of phosphorylated PI3K (P-PI3K), phosphorylated AKT (P-AKT), mTOR and phosphorylated mTOR (P-mTOR) under inflammatory conditions than did Ad-control-transduced PDLSCs (p < 0.001; Fig. 6). The P2X7 antagonist BBG decreased mTOR expression by 74% and P-mTOR levels by nearly half (p < 0.001). In contrast, activation of P2X7R BzATP increased P-AKT levels by nearly 1.5-fold (p < 0.001) but led to a decrease in mTOR (over 70%; p < 0.001) and P-mTOR (over 50%; p < 0.01) levels in gene-modified PDLSCs (Fig. 6c).

a Images and quantitative analysis of PI3K and P-PI3K levels measured by western blotting of protein derived from PDLSCs with Ad-control transduction or with Ad-P2X7 transduction at day 7. b Images and quantitative analysis of AKT and P-AKT levels measured by western blotting of protein derived from PDLSCs with Ad-control transduction or with Ad-P2X7 transduction at day 7. c Images and quantitative analysis of mTOR and P-mTOR levels measured by western blotting of protein derived from PDLSCs with Ad-control transduction or with Ad-P2X7 transduction at day 7. Data are presented as the mean ± S.D. for n = 3–8; ns indicates no significant difference between the indicated columns; *p < 0.05, **p < 0.01, and ***p < 0.001 indicate significant differences between the indicated columns
Involvement of PI3K-AKT-mTOR signaling in PDLSC osteogenesis under inflammatory conditions: The P2X7R antagonist BBG (10 μM) and specific inhibitors of PI3K (NVP-BKM120, 1 μM), AKT (MK2206, 10 μM) and mTOR (Rapamycin, 1 μM) were employed to further confirm the involvement of PI3K-AKT-mTOR signaling in PDLSC osteogenesis under inflammatory conditions. As expected, treatment of gene-modified PDLSCs with BBG, NVP-BKM120, MK2206, or Rapamycin under inflammatory conditions largely blocked osteogenesis in Ad-P2X7-transfected PDLSCs, as evidenced by ALP and Alizarin red staining (Fig. 7a, b), and levels of Runx2 and OCN protein expression were significantly decreased (Fig. 7c, d).

a ALP staining (scale bar = 1 mm) and the quantitative result. b Alizarin red staining (scale bar = 500 μm) and the quantitative result of Ad-P2X7-transfected PDLSCs. Western blotting analysis of c Runx2 and d OCN expression in Ad-P2X7-transfected PDLSCs. Data are presented as the mean ± S.D. for n = 4; ns indicates no significant difference between the indicated columns; *p < 0.05, **p < 0.01, and ***p < 0.001 indicate significant differences between the indicated columns
Discussion
Although PDLSCs have been proposed for decades for use in periodontal regeneration, outcomes arising from clinical stem cell-based therapies have been far from successful3,4,5. Accumulating evidence suggests that the failure of stem cells to exert their functionality is most likely due to the inhospitable local microenvironment of the diseased periodontium, in which multiple molecules and processes participate in pathological interactions and the altered host-biofilm interplay of periodontitis, including inflammation11, hypoxia37, and ATP38. As a key inflammatory mediator, ATP acts as not only an energy supply molecule but also a signal transducer that is released upon stimulation (e.g., stress or inflammation) or due to passive leakage by damaged or dying cells. Accordingly, ATP plays a key role in the pathophysiology of alveolar bone loss in periodontitis39,40. Extracellular ATP can activate purinergic receptors, including those in the metabotropic receptor family (P2Y, G protein-coupled receptors) and the ionotropic receptor family (P2X, ionotropic receptors)41. To date, seven P2X receptors and eight P2Y receptors have been identified in mammals. P2X7R belongs to the P2X receptor family, and its activation results in the formation of plasma membrane pores to allow for significant calcium influx; however, passage of molecules up to 900 Da following large pore formation may lead to rapid cell death42. Previous studies have shown that extracellular ATP enhances IL-1β expression in human PDL cells via P2X7R25 and induces release of the proinflammatory cytokines IL-8 and CCL20 from human PDLSCs27. Regardless, the role of P2X7R in PDLSC functionality in an inflammatory environment remains unknown.
In this study, a series of assays were designed to identify the effects of P2X7R on the osteogenic differentiation of PDLSCs in inflammatory microenvironments and the underlying mechanisms involved in P2X7R-associated changes in osteogenesis. We isolated PDLSCs from volunteers with good periodontal health (Supplementary Fig. 1); based on our previous experience, TNF-α (10 ng/mL) and IL-1β (5 ng/mL) were added to culture media to mimic the inflammatory microenvironment in vitro13,43. P2X7R expression in PDLSCs was demonstrated at both the gene and protein levels, whereby P2X7R expression was decreased in the presence of inflammatory factors but significantly increased in an osteo-inductive environment in the absence of inflammatory factors (Fig. 1). The results suggest that P2X7R expression is closely linked to the cellular microenvironment, at least with regard to inflammation and osteo-induction, a finding that is consistent with previous evidence that P2X7R expression increases with osteoblast differentiation35,44 and that P2X7R activation may enhance osteoblast differentiation and matrix mineralization45. Considering that P2X7R expression changes significantly in response to inflammatory and/or osteo-inductive conditions, the role of P2X7R in the inflammation-mediated decrease in PDLSC osteogenesis warrants further exploration.
As a specific P2X7R agonist, BzATP is more stable and approximately tenfold more potent than ATP, with similar consequences11,46. In this study, dose–response data indicated that 100 μM BzATP significantly enhanced osteogenesis under inflammatory conditions, with Runx2 and OCN expression in PDLSCs being increased by 4.62-fold and 9.6-fold, respectively, in osteo-inductive medium (Fig. 2). Further testing using P2X7-selective antagonist A740003 confirmed that the BzATP-induced effects are P2X7 specific (Supplementary Fig. 2). BBG is also a specific P2X7R antagonist, and treatment of PDLSCs with BBG at the previously reported concentration of 10 μM42 decreased the formation of mineralized nodules under osteo-inductive conditions and abolished increases in Runx2 and OCN expression at both the mRNA and protein levels, suggesting that P2X7R activity is linked to PDLSC osteogenesis (Fig. 2). These results are in accordance with previous findings based on BMMSCs22,29. For example, shockwave treatment was found to enhance ATP release and induce osteogenic differentiation in human BMMSCs in a P2X7R-dependent manner, whereas blocking P2X7R with siRNA abolished osteogenesis29. In another study, BMMSCs derived from postmenopausal women exhibited a P2X7R-dependent increase in osteogenic differentiation and mineralization; ATP/BzATP elicited reversible membrane blebbing (zeiosis) in these cells as well as progressively increased Runx2 and osterix expression, ALP activity and mineralization in a Rho-kinase-dependent manner 22.
Given that PDLSCs expressed higher levels of P2X7R under osteo-inductive conditions and that inflammation decreased P2X7R expression, we further explored whether the regulation of P2X7R in PDLSCs can reverse the impaired PDLSC osteogenesis observed under inflammatory conditions. Adenovirus vectors carrying the P2X7R gene were used to enhance P2X7R expression on PDLSCs, and we found that transfection at an MOI = 20 significantly increased P2X7R expression (p < 0.001; Fig. 3a, b). Moreover, the proliferative capacity of PDLSCs was also enhanced at this concentration, particularly at days 5 and 6 (p < 0.05; Fig. 3c). Furthermore, compared with unmodified cells, gene-transduced PDLSCs exhibited a significant increase in osteogenesis ability under inflammatory conditions, and inhibiting P2X7R with BBG abolished gene-enhanced osteogenesis (Fig. 4). Although the roles of P2X7R in PDLSCs under inflammatory conditions have not yet been reported, it has been reported that P2X7R is involved in mechanical stimulation-induced inflammatory cytokine release by human PDL cells25. Indeed, previous studies have demonstrated that P2X7R has positive functions in shockwave-induced osteogenesis in human MSCs29, osteogenic differentiation of MSCs in a mouse model of osteoporosis34 and tension force-induced osteogenesis in osteoblasts31,47. In addition to these in vitro stem cell experiments, an in vivo study reported that P2X7R-knockout mice have deficient periosteal bone formation and excessive trabecular bone resorption48, and in humans, P2X7R gain-of-function polymorphisms result in reduced fracture risk and increased bone mineral density33. Despite mounting evidence that the promotion of osteogenesis during skeletal development is coupled to P2X7R activation in osteoblasts32, this is not always the case. For example, specific P2X7R activation was found to impair bone mineralization and osteoblast ALP activation in neonatal rats35, and the P2X7R agonist BzATP-induced apoptosis in SaOS-2 (human osteosarcoma cell line) osteoblasts36. In another study, BALB/cJ P2X7R-knockout mice displayed reduced levels of serum C-telopeptide fragment (s-CTX; an indicator of bone resorption), higher bone mineral density, and increased bone strength49. Although inconsistent data have been reported, the crucial role of P2X7R in osteogenesis and skeletal mechanotransduction has been confirmed. However, this receptor is downregulated under pathological conditions (e.g., inflammation due to pathogen involvement or high-glucose concentrations due to diabetes), hereby blunting its molecular effects (reviewed in ref. 50). It is now proposed that ATP concentrations sufficiently high to activate P2X7R are present only in damaged cells/tissues under pathophysiological conditions or when a high-intensity mechanical load is placed on the skeleton. Osteoclasts, osteoblasts and MSCs play different roles under these conditions, which may partially explain the sometimes yield inconsistent results of in vitro cellular studies and in vivo knockout studies 50.
Based on data obtained from the aforementioned experiments, we conclude that P2X7R expressed on PDLSCs plays a positive role in PDLSC osteogenic differentiation under inflammatory conditions. To further explore the mechanism underlying P2X7R-induced PDLSC osteogenesis, we used qRT-PCR to evaluate 14 osteogenesis-related genes from the MAPK18,37,51,52, Wnt/β-catenin12,14,15,16 and PI3K-AKT13,53,54 families in P2X7R-overexpressing PDLSCs, genes that were previously reported to be related to PDLSC osteogenesis. We excluded Wnt-3α because it exhibited relatively stable expression irrespective of the incubation conditions, and another three genes (AP-1, GSK-3β, and CTNNB) were excluded because their expression levels were not significantly altered by inflammatory factors. FOS-B, FOX-O, and NFκB were subjected to quantitative analysis, and the results showed no statistically significant changes in expression in response to BBG treatment (Fig. 5b–d). The mTOR gene in the PI3K-AKT family was found to likely be involved in P2X7R-enhanced PDLSC osteogenesis (Fig. 5).
Phosphorylation often activates a protein. Thus, to completely assess activation of a cell signaling molecule, both total and phosphorylated levels of proteins in the PI3K-AKT-mTOR pathway were examined by western blotting. We found that overexpression of P2X7R by adenovirus significantly increased mTOR expression and phosphorylation of PI3K, AKT, and mTOR. Blocking P2X7R activation with BBG decreased phosphorylation of PI3K and AKT and expression of mTOR (Fig. 6). Furthermore, suppression of PI3K-AKT-mTOR signaling by specific antagonists (BKM120, MK2206, and Rapamycin) abolished P2X7R-induced osteogenesis, as confirmed by western blotting and ALP and Alizarin red staining (Fig. 7). These findings were in accordance with those of previous studies, in which dysregulation of PI3K-AKT-mTOR signaling was associated with osteosarcoma cell proliferation and growth55 and inhibition of the PI3K-AKT pathway suppressed osteosarcoma cell proliferation and growth56. In addition, integrin alpha 5-induced osteogenesis of PDLSCs57 and hepatocyte growth factor-induced osteogenesis of human BMMSCs58 demonstrate the involvement of a PI3K-AKT-dependent pathway. In fact, the function of mTOR in skeletal development and disease has been well documented (reviewed in ref. 54), and it is now clear that mTOR (including mTORC1 and mTORC2) is a common effector downstream of multiple bone anabolic signals, such as BMP259, β-catenin60 and Wnt (Wnt10β, Wnt7β, and Wnt3α) signaling61, wherein it promotes osteoclastogenesis both directly and indirectly (reviewed in ref. 54). Further evidence indicates that inhibition of mTOR by Rapamycin impairs the proliferation and osteogenic differentiation of mouse BMMSCs and hence reduces trabecular bone mass. In addition, mTOR-knockout transgenic mice exhibit reduced limb length resulting from shrunken epiphyseal growth plates, as well as dysfunctional intramembranous and endochondral ossification62, and activation of mTORC1 by insulin-like growth factor-1 has been shown to induce osteoblast differentiation in BMMSCs63,64. Indeed, other growth factors, such as platelet-derived growth factor receptor-β and Src homology 2-domain-containing inositol 5'-phosphatase 1, can also enhance the proliferation and osteogenic differentiation of MSCs through PI3K-AKT-mTOR/β-catenin signaling65,66. Recent evidence suggests that mTORC2 is also involved in age-related bone loss and the osteoblast switch 67,68.
Combined with these previous data, our findings reveal the importance of the PI3K-AKT-mTOR axis in P2X7R-induced osteogenic differentiation and mineralization of PDLSCs under inflammatory conditions, thus pointing to a new therapeutic target for combating bone loss in periodontal disease. Based on data obtained from this study and the published literature, we summarized the molecules related to PI3K-AKT-mTOR signaling and P2X7R activation in a signaling network, as shown in Fig. 8. The limitations of our study should also be acknowledged. We used an in vitro model to explore the function of P2X7R under inflammatory conditions, though the actual function of P2X7R in vivo should be explored. Furthermore, the safety of using Ad-P2X7 to upregulate P2X7R expression in vivo should be taken into consideration. Much work remains before we can draw a clear pathway for P2X7R-mediated osteogenesis under inflammatory conditions.

Schematic illustration of the potential molecules related to PI3K-AKT-mTOR signaling that are involved in P2X7R-enhanced osteogenesis of PDLSCs
Conclusion
The present study investigated P2X7R expression in PDLSCs under normal and inflammatory conditions and examined how BzATP and BBG influence PDLSC osteogenesis under various conditions in a coordinated fashion. Furthermore, the role of P2X7R and the signaling pathway underlying P2X7R-enhanced osteogenesis under inflammatory conditions were investigated using gene-modified PDLSCs. This is the first study on interrogating the role of P2X7R in inflammation-compromised PDLSC osteogenic differentiation, and the PI3K-AKT-mTOR signaling pathway was found to be involved in this process. Our data identify a crucial role for P2X7R in osteogenesis and the response to inflammation that affects the functionality of PDLSCs under inflammatory conditions. However, the exact roles of P2X7R, alone or in combination with other cell-surface receptor(s), remain unknown and warrant in-depth investigation. Given that P2X7R may exert different influences on osteogenesis, further studies using appropriate animal models are required before P2X7R can be developed as a new therapeutic target to rescue inflammation-mediated changes in PDLSCs.
Materials and methods
Isolation and purification of PDLSCs
Extracted human third molars or orthodontic teeth used for PDLSC isolation were donated by volunteers, including 4 males and 5 females, aged 18–45 years with good periodontal health. Tooth extraction was performed at the Dental Clinic of School of Stomatology, Fourth Military Medical University (FMMU), and all donors signed the informed consent form. The use of extracted human teeth for cell isolation and subsequent research was approved by the Ethics Committee at FMMU. PDLSCs were isolated based on our reported protocols, with slight modification8. Briefly, freshly extracted teeth were repeatedly rinsed with sterile phosphate-buffered saline (PBS; Corning, New York, USA), and PDL tissue was scraped from the middle of the root surface. The collected tissue was cut into blocks (1 × 1 ×1 mm3) and digested in 3 mg/mL collagenase type I (Sigma-Aldrich, St. Louis, USA) for 1 h. Finally, the digested blocks were cultured in 6-well plates (Invitrogen, Carlsbad, CA, USA) in α-minimum essential medium (α-MEM, Invitrogen) containing 100 U/mL penicillin (Invitrogen), 100 μg/mL streptomycin (Invitrogen), and 10% fetal bovine serum (FBS; Sijiqing, Hangzhou, China). The medium was changed every 3 days until the primary cells migrated from the tissue blocks and achieved confluence, after which, the limiting dilution technique was applied for PDLSC purification. PDLSCs at passage 2–4 (P2–P4) were used in the experiments.
Characterizations of PDLSCs
Flow cytometric analysis: A cell (P2) suspension was divided into sterile Eppendorf tubes (Eppendorf, Hamburg, Germany) and incubated at 4 °C with fluorescein isothiocyanate-conjugated anti-human CD105, CD90, or CD45; phycoerythrin-conjugated anti-human CD31 or CD146; or allophycocyanin-conjugated anti-human CD34 (eBioscience, CA, USA) at a 1:1000 dilution. Cells incubated without antibodies were used as the negative control. After a 1-h incubation, excess antibodies were removed, and PDLSCs were resuspended in PBS (300 μL). The immunophenotypes of the PDLSCs were analyzed using a Beckman Coulter Epics XL cytometer (Beckman Coulter, Fullerton, CA, USA).
Colony formation assay: PDLSCs (P3) were plated into 100-mm culture dishes (Invitrogen) at 1 × 103 cells/dish in complete α-MEM; the medium was refreshed every 3 days. After 14 days, the cells were fixed in 4% paraformaldehyde (Servicebio, Wuhan, China) for 20 min and stained with 0.1% crystal violet (Sigma-Aldrich) for 15 min. Colonies were counted, and inverted microscopy was used to exclude colonies that contained fewer than 50 cells.
Cell proliferation assay: PDLSCs (P3) were seeded in 96-well plates (Invitrogen) at 2 × 104 cells/well. After 24 h under different culture conditions, 20 μL of Cell Counting Kit-8 solution (Dojindo Corporation, Tokyo, Japan) was added to each well, and the plates were incubated for another 2 h before the absorbance was measured at 450 nm using an Infinite M200 PRO microplate reader (TECAN, Männedorf, Switzerland); measurements were performed at the same time of day during the 7-day culture.
Cell differentiation assay: PDLSCs (P3) were plated in 6-well dishes at 5 × 105 cells/well. When the cells reached 80% confluence, they were incubated in osteo-inductive medium, adipo-inductive medium or chondrogenic inductive medium to ascertain their multilineage differentiation capacity. The osteo-inductive medium contained α-MEM and 10% FBS supplemented with 100 U/mL penicillin, 100 μg/mL streptomycin, 10 mM β-glycerophosphate, 50 μg/mL l-ascorbic-2-phosphate and 10 nM dexamethasone (all from Sigma-Aldrich). The adipogenic medium contained α-MEM and 10% FBS supplemented with 100 U/mL penicillin, 100 μg/mL streptomycin, 0.5 mM 3-isobutyl-1-methylxanthine (IBMX; MP Biomedicals, Illkrich, France), 1 μM dexamethasone, 0.1 mM indomethacin and 10 μg/mL insulin (all from Sigma-Aldrich). The chondrogenic differentiation medium containing dexamethasone, ascorbate, ITS+ supplement, sodium pyruvate, proline, and TGF-β3 were purchased from Cyagen (Guangzhou, Guangdong, China). After a 3-week induction (for osteo-induction or adipo-induction) or 4-week induction (for chondrogenic induction), differentiated PDLSCs were fixed with 4% paraformaldehyde, and mineralized nodes, lipid droplets or acidic polysaccharides were visualized by Alizarin red staining (Heart, Xi’an, China), Oil red O staining (Heart), and Alcian blue staining (Cyagen), as indicated. To quantify the Alizarin red staining for statistical analysis, the mineralized nodes were dissolved in 2% cetylpyridinium chloride for 15 min, and OD values were measured at 560 nm ref. 8.
P2X7R expression in PDLSCs under inflammatory conditions
To investigate the effect of inflammation on P2X7R expression in PDLSCs, we used TNF-α (10 ng/mL) and IL-1β (5 ng/mL; both from Novoprotein Scientific Inc., Shanghai, China) to establish an in vitro inflammatory microenvironment according to previous publications69,70. mRNA and protein expression of P2X7R in PDLSCs under normal and osteogenic conditions was determined by quantitative real-time polymerase chain reaction (qRT-PCR) (see Section qRT-PCR) and western blot analysis (see Section Western blot analysis), respectively. Cells cultivated in parallel in a normal microenvironment were used as the control.
Osteogenesis of PDLSCs in response to P2X7R agonist or antagonist under inflammatory conditions
A specific agonist and antagonist were used to clarify how P2X7R may affect PDLSC osteogenesis under inflammatory conditions. We employed the P2X7 agonist 3′-O-(4-benzoylbenzoyl) adenosine 5′-triphosphate (BzATP; Sigma-Aldrich) at a concentration of 100 μM, as selected based on dose-response assays. According to previous data42, we utilized the P2X7 antagonist Brilliant Blue G (BBG; Sigma-Aldrich) at a concentration of 10 μM. The osteogenic differentiation potential of PDLSCs was evaluated by Alizarin red staining (see Section Cell differentiation assay) and ALP staining (see Section ALP staining), and osteogenesis-related gene/protein expression (Runx2 and OCN) was determined by qRT-PCR (see Section qRT-PCR) and western blot analysis (see Section Western blot analysis). Further, osteogenesis-related gene/protein expressions in PDLSCs in response to the addition of P2X7-selective antagonist A740003 (MCE, NJ, USA) at a concentration of 0.01, 0.1, 1, 10, and 100 μM, respectively were also determined (see Sections qRT-PCR and Western blot analysis) to confirm the BzATP-induced effects are P2X7 specific.
Osteogenesis of gene-modified PDLSCs under inflammatory conditions
Adenoviral vector transduction: Considering that gene-enhanced cells can express higher levels of P2X7R irrespective of incubation conditions, PDLSCs were transfected with adenoviral vectors (Likeli Bioscience Inc., Beijing, China) containing P2X7R cDNA (Ad-P2X7). To identify a suitable MOI, cells were transfected with 10, 20, 50, and 100 pfu/cell; cells transfected with an adenoviral construct lacking the P2X7R sequence (Ad-control) were used as negative control. Adenovirus-mediated gene transfer was performed by adding a minimal volume of medium containing 2% FBS and adenovirus. After an 8-h incubation, the full volume of medium was replaced with α-MEM containing 10% FBS. The ensuing assays were performed 24 h after adenoviral infection.
Immunofluorescence staining for P2X7R in gene-modified cells: To evaluate the outcome of cell transfection, Ad-P2X7-transfected PDLSCs were seeded in 35-mm culture dishes designed for confocal microscopy (Thermo Fisher, MA, USA) and cultured in α-MEM containing 10% FBS. After 8 h of transduction and 24 h of incubation, adherent cells were fixed with 4% paraformaldehyde, permeabilized with 0.5% Triton X-100 (Sigma-Aldrich) for 10 min and blocked with 1% bovine serum albumin (BSA; Sigma-Aldrich) for 30 min at room temperature. The cells were then incubated overnight at 4 °C with an antibody against P2X7R (1:500, Abcam, ab48871); the samples were washed with PBS before being incubated for 2 h in the dark at room temperature with Alexa Fluor 594 AffiniPure donkey anti-rabbit IgG secondary antibody (1:500; Yeasen Biotech Company, Shanghai, China). Finally, the cells were counterstained for 10 min in the dark at room temperature with 4′,6-diamidino-2-phenylindole (DAPI; Abcam, ab104139). Images were obtained using a confocal laser scanning microscope (FV1000; Olympus, Tokyo, Japan).
Osteogenesis of gene-modified PDLSCs in response to inflammation and/or P2X7R antagonist: The osteogenic differentiation potential of gene-enhanced PDLSCs was evaluated in terms of ALP staining (see Section ALP staining), Alizarin red staining (see Section Cell differentiation assay), and western blot analysis (Runx2 and OCN; see Section Western blot analysis).
Potential signaling pathways involved in P2X7R-induced osteogenesis
Screening and identification of potential target genes: To explore the potential mechanisms of P2X7R-induced osteogenesis, we first used qRT-PCR to screen for possible molecules involved in this process. Genetic evidence has shown that the MAPK and Wnt/β-catenin pathways are important during osteogenesis, and stimulation of P2X7R in BMMSCs under normal conditions is associated with PI3K-AKT-mTOR activation29. Therefore, we selected genes in the MAPK (AP-1 and FOS-B), Wnt/β-catenin (Wnt-3α, GSK-3β and CTNNB), and PI3K-AKT (FOX-O, NF-κB and mTOR) pathways for qRT-PCR screening to identify genes sensitive to both a P2X7R agonist and antagonist. We considered genes more likely to be involved in P2X7R-induced osteogenesis if their expression levels changed significantly in response to either the P2X7R agonist or antagonist.
Upstream signals involved in the potential pathway: Western blot analysis (see Section Western blot analysis) was applied to evaluate protein expression profiles in PDLSCs (with Ad-P2X7R transfection or Ad-control transfection) in response to inflammation and/or addition of P2X7R agonist/antagonist.
Osteogenic abilities of gene-enhanced PDLSCs in response to P2X7 antagonist or pathway inhibition: The osteogenic differentiation potential of Ad-P2X7-transfected PDLSCs following addition of the P2X7R antagonist BBG or inhibitors (MCE) of the identified signaling molecules (specific inhibitors were chosen based on identified genes, see Section Target gene screening) was evaluated in terms of ALP staining (see Section ALP staining), Alizarin red staining (see Section Cell differentiation assay), and western blot analysis (Runx2 and OCN; see Section Western blot analysis).
ALP staining
ALP activity in PDLSCs was analyzed by ALP staining. In brief, PDLSCs were seeded at 1 × 105 cells/well in 6-well plates, and ALP staining was performed after 7-day osteo-inductive culture using a BCIP/NBT ALP staining kit (Beyotime Institute of Biotechnology, China) according to the instructions. ALP quantitative analysis was performed using an alkaline phosphatase assay kit (Nanjing Jiancheng Bioengineering Institute, China) according to the instructions.
qRT-PCR
Total RNA was extracted from PDLSCs (prepared for analysis) using Takara MiniBEST Universal RNA Extraction Kit according to the manufacturer’s instructions (Takara Biological Incorporated Company, Kyoto, Japan) and was reverse transcribed using RevertAid First Strand cDNA Synthesis Kit (Yeasen Biotech Company). qRT-PCR was performed on cDNA samples diluted tenfold in double-distilled water using SYBR Green Master Mix (Yeasen Biotech Company) and a CFX Connect™ Real-Time PCR Detection System (Bio-Rad, Hercules, CA, USA). Amplification was performed under the following conditions: denaturation at 95 °C for 5 min; 40 cycles at 95 °C for 10 s, 55 °C for 20 s, and 72 °C for 20 s, with melt curve analysis from 65 to 95 °C in increments of 0.5 °C. The gene-specific qRT-PCR primers were specific for humans, and the sequences of all primers used in the present study are shown in Supplementary Table. 1.
Western blot analysis
The protein expression profiles of PDLSCs (prepared for analysis) were measured by western blotting. Briefly, total cellular protein was extracted in lysis buffer (Heart) containing phosphatase inhibitors (Sigma-Aldrich). The protein concentration was measured using a BCA Protein Assay Kit (Heart). Forty micrograms of protein was separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis (10% for P2X7R, P-PI3K, PI3K, P-AKT, and AKT; 15% for OCN; and 6% for P-mTOR and mTOR). The proteins were transferred onto PVDF membranes (Millipore, Billerica, MA, USA), which were then blocked in 5% BSA at room temperature for 1 h, incubated overnight at 4 °C with primary antibodies against P2X7R (1:1000; CST, #13809), GAPDH (1:1000; Proteintech, Chicago, #10494-1-AP), OCN (1:1000; Abcam, ab93876), Runx2 (1:1000; CST, #12556), PI3K (1:1000; CST, #4257), P-PI3K (1:1000; CST, #4228), AKT (1:1000; CST, #4691), P-AKT (1:1000; CST, #4060), mTOR (1:1000; CST, #2983), and P-mTOR (1:1000; CST, #5536), and further incubated for 1 h at 22 °C with an anti-rabbit secondary antibody (1:5000; Jackson, USA). The blots were developed using Western Chemiluminescent HRP Substrate (EMD Millipore; Massachusetts; USA). Quantification of the final images was performed using Quantity One software (Version 4.6.2).
Statistical analysis
All results are presented as the mean and standard deviation (mean ± S.D.) for n from 3 to 8; experiments for each cell line were performed independently and in triplicate. GraphPad Prism 7.03 software was utilized for statistical tests. We used Student's t-test for analysis of two unpaired groups, and one-way ANOVA followed by Tukey’s posttest for analysis of multiple groups. Statistical significance was established at p < 0.05.
Change history
20 December 2023
This article has been retracted. Please see the Retraction Notice for more detail: https://doi.org/10.1038/s41419-023-06390-y
References
Pihlstrom, B. L., Michalowicz, B. S. & Johnson, N. W. Periodontal diseases. Lancet 366, 1809–1820 (2005).
Kinane, D. F., Stathopoulou, P. G. & Papapanou, P. N. Periodontal diseases. Nat. Rev. Dis. Primers 3, 17038 (2017).
Chen, F. M., Sun, H. H., Lu, H. & Yu, Q. Stem cell-delivery therapeutics for periodontal tissue regeneration. Biomaterials 33, 6320–6344 (2012).
Bright, R., Hynes, K., Gronthos, S. & Bartold, P. M. Periodontal ligament-derived cells for periodontal regeneration in animal models: a systematic review. J. Periodontal Res. 50, 160–172 (2015).
Tassi, S. A., Sergio, N. Z., Misawa, M. Y. O. & Villar, C. C. Efficacy of stem cells on periodontal regeneration: systematic review of pre-clinical studies. J. Periodontal Res. 52, 793–812 (2017).
Chen, F. M. et al. Treatment of periodontal intrabony defects using autologous periodontal ligament stem cells: a randomized clinical trial. Stem Cell Res. Ther. 7, 33 (2016).
Park, J. C. et al. Isolation and characterization of human periodontal ligament (PDL) stem cells (PDLSCs) from the inflamed PDL tissue: in vitro and in vivo evaluations. J. Clin. Periodontol. 38, 721–731 (2011).
Wu, R. X., Bi, C. S., Yu, Y., Zhang, L. L. & Chen, F. M. Age-related decline in the matrix contents and functional properties of human periodontal ligament stem cell sheets. Acta Biomater. 22, 70–82 (2015).
Liu, D. et al. Mesenchymal stem cells derived from inflamed periodontal ligaments exhibit impaired immunomodulation. J. Clin. Periodontol. 39, 1174–1182 (2012).
Tang, H. N. et al. Stem cells derived from “inflamed” and healthy periodontal ligament tissues and their sheet functionalities: a patient-matched comparison. J. Clin. Periodontol. 43, 72–84 (2016).
Liu, N. et al. High levels of beta-catenin signaling reduce osteogenic differentiation of stem cells in inflammatory microenvironments through inhibition of the noncanonical Wnt pathway. J. Bone Miner. Res. 26, 2082–2095 (2011).
Chen, X. et al. Nuclear factor-κB modulates osteogenesis of periodontal ligament stem cells through competition with β-catenin signaling in inflammatory microenvironments. Cell Death Dis. 4, e510 (2013).
Kong, X. et al. GSK3β is a checkpoint for TNF-α-mediated impaired osteogenic differentiation of mesenchymal stem cells in inflammatory microenvironments. Biochim. Biophys. Acta 1830, 5119–5129 (2013).
Liu, W. et al. Canonical Wnt signaling differently modulates osteogenic differentiation of mesenchymal stem cells derived from bone marrow and from periodontal ligament under inflammatory conditions. Biochim. Biophys. Acta 1840, 1125–1134 (2014).
Liu, Q. et al. DKK1 rescues osteogenic differentiation of mesenchymal stem cells isolated from periodontal ligaments of patients with diabetes mellitus induced periodontitis. Sci. Rep. 5, 13142 (2015).
Li, B. et al. GCN5 modulates osteogenic differentiation of periodontal ligament stem cells through DKK1 acetylation in inflammatory microenvironment. Sci. Rep. 6, 26542 (2016).
Xue, P. et al. Decreased MORF leads to prolonged endoplasmic reticulum stress in periodontitis-associated chronic inflammation. Cell Death Differ. 23, 1862–1872 (2016).
Mao, C. Y. et al. Double-edged-sword effect of IL-1beta on the osteogenesis of periodontal ligament stem cells via crosstalk between the NF-kappaB, MAPK and BMP/Smad signaling pathways. Cell Death Dis. 7, e2296 (2016).
Liu, A. Q., Hu, C. H., Jin, F., Zhang, L. S. & Xuan, K. Contributions of bioactive molecules in stem cell-based periodontal regeneration. Int. J. Mol. Sci. 19, E1016 (2018).
Griffin, M. D. et al. Concise review: adult mesenchymal stromal cell therapy for inflammatory diseases: how well are we joining the dots? Stem Cells 31, 2033–2041 (2013).
Bassir, S. H. et al. Potential for stem cell-based periodontal therapy. J. Cell. Physiol. 231, 50–61 (2016).
Noronha-Matos, J. B. et al. P2X7-induced zeiosis promotes osteogenic differentiation and mineralization of postmenopausal bone marrow-derived mesenchymal stem cells. FASEB J. 28, 5208–5222 (2014).
Feng, W. et al. High level P2X7-mediated signaling impairs function of hematopoietic stem/progenitor cells. Stem Cell Rev. 12, 305–314 (2016).
Lovelace, M. D. et al. P2X7 receptors mediate innate phagocytosis by human neural precursor cells and neuroblasts. Stem Cells 33, 526–541 (2015).
Kanjanamekanant, K., Luckprom, P. & Pavasant, P. Mechanical stress-induced interleukin-1beta expression through adenosine triphosphate/P2X7 receptor activation in human periodontal ligament cells. J. Periodontal Res. 48, 169–176 (2013).
Kanjanamekanant, K., Luckprom, P. & Pavasant, P. P2X7 receptor-Pannexin1 interaction mediates stress-induced interleukin-1 beta expression in human periodontal ligament cells. J. Periodontal Res. 49, 595–602 (2014).
Trubiani, O. et al. Expression of P2X7 ATP receptor mediating the IL8 and CCL20 release in human periodontal ligament stem cells. J. Cell. Biochem. 115, 1138–1146 (2014).
Pasqualetto, G., Brancale, A. & Young, M. T. The molecular determinants of small-molecule ligand binding at P2X receptors. Front. Pharmacol. 9, 58 (2018).
Sun, D. et al. Shockwaves induce osteogenic differentiation of human mesenchymal stem cells through ATP release and activation of P2X7 receptors. Stem Cells 31, 1170–1180 (2013).
Agrawal, A. & Gartland, A. P2X7 receptors: role in bone cell formation and function. J. Mol. Endocrinol. 54, R75–R88 (2015).
Kariya, T. et al. Tension force-induced ATP promotes osteogenesis through P2X7 receptor in osteoblasts. J. Cell. Biochem. 116, 12–21 (2015).
Panupinthu, N. et al. P2X7 nucleotide receptors mediate blebbing in osteoblasts through a pathway involving lysophosphatidic acid. J. Biol. Chem. 282, 3403–3412 (2007).
Husted, L. B. et al. Functional polymorphisms in the P2X7 receptor gene are associated with osteoporosis. Osteoporos. Int. 24, 949–959 (2013).
Wang, N., Agrawal, A., Jorgensen, N. R. & Gartland, A. P2X7 receptor regulates osteoclast function and bone loss in a mouse model of osteoporosis. Sci. Rep. 8, 3507 (2018).
Orriss, I. R. et al. The regulation of osteoblast function and bone mineralisation by extracellular nucleotides: the role of p2x receptors. Bone 51, 389–400 (2012).
Gartland, A., Hipskind, R. A., Gallagher, J. A. & Bowler, W. B. Expression of a P2X7 receptor by a subpopulation of human osteoblasts. J. Bone Miner. Res. 16, 846–856 (2001).
Wu, Y. et al. The osteogenic differentiation of PDLSCs is mediated through MEK/ERK and p38 MAPK signalling under hypoxia. Arch. Oral Biol. 58, 1357–1368 (2013).
Kawase, T., Okuda, K. & Yoshie, H. Extracellular ATP and ATPγS suppress the proliferation of human periodontal ligament cells by different mechanisms. J. Periodontol. 78, 748–756 (2007).
Lim, J. C. & Mitchell, C. H. Inflammation, pain, and pressure—purinergic signaling in oral tissues. J. Dent. Res. 91, 1103–1109 (2012).
Binderman, I., Gadban, N. & Yaffe, A. Extracellular ATP is a key modulator of alveolar bone loss in periodontitis. Arch. Oral. Biol. 81, 131–135 (2017).
Burnstock, G. The past, present and future of purine nucleotides as signalling molecules. Neuropharmacology 36, 1127–1139 (1997).
Jiang, L. H., Mackenzie, A. B., North, R. A. & Surprenant, A. Brilliant blue G selectively blocks ATP-gated rat P2X7 receptors. Mol. Pharmacol. 58, 82–88 (2000).
Yu, Y. et al. Effects of short-term inflammatory and/or hypoxic pretreatments on periodontal ligament stem cells: in vitro and in vivo studies. Cell Tissue Res. 366, 311–328 (2016).
Orriss, I. R., Knight, G. E., Ranasinghe, S., Burnstock, G. & Arnett, T. R. Osteoblast responses to nucleotides increase during differentiation. Bone 39, 300–309 (2006).
Panupinthu, N. et al. P2X7 receptors on osteoblasts couple to production of lysophosphatidic acid: a signaling axis promoting osteogenesis. J. Cell Biol. 181, 859–871 (2008).
Park, J. H. & Kim, Y. C. P2X7 receptor antagonists: a patent review (2010–2015). Expert Opin. Ther. Pat. 27, 257–267 (2017).
Sun, C. et al. Tensile strength suppresses the osteogenesis of periodontal ligament cells in inflammatory microenvironments. Mol. Med. Rep. 16, 666–672 (2017).
Ke, H. Z. et al. Deletion of the P2X7 nucleotide receptor reveals its regulatory roles in bone formation and resorption. Mol. Endocrinol. 17, 1356–1367 (2003).
Syberg, S. et al. Genetic background strongly influences the bone phenotype of P2X7 receptor knockout mice. J. Osteoporos. 2012, 391097 (2012).
Jørgensen, N. R. The purinergic P2X7 ion channel receptor—a ‘repair’ receptor in bone. Curr. Opin. Immunol. 52, 32–38 (2018).
Niu, C. et al. Gold nanoparticles promote osteogenic differentiation of human periodontal ligament stem cells via the p38 MAPK signaling pathway. Mol. Med. Rep. 16, 4879–4886 (2017).
Wang, Y. et al. Mineral trioxide aggregate enhances the osteogenic capacity of periodontal ligament stem cells via NF-kappaB and MAPK signaling pathways. J. Cell. Physiol. 233, 2386–2397 (2018).
Wang, Y. et al. Roxarsone induces angiogenesis via PI3K/Akt signaling. Cell Biosci. 6, 54 (2016).
Chen, J. & Long, F. mTOR signaling in skeletal development and disease. Bone Res. 6, 1 (2018).
Keremu, A. et al. NRSN2 promotes osteosarcoma cell proliferation and growth through PI3K/Akt/MTOR and Wnt/β-catenin signaling. Am. J. Cancer Res. 7, 565–573 (2017).
Zhang, J., Yu, X. H., Yan, Y. G., Wang, C. & Wang, W. J. PI3K/Akt signaling in osteosarcoma. Clin. Chim. Acta 444, 182–192 (2015).
Wang, H. et al. Priming integrin alpha 5 promotes the osteogenic differentiation of human periodontal ligament stem cells due to cytoskeleton and cell cycle changes. J. Proteom. 179, 122–130 (2018).
Wen, Q. et al. The multiplicity of infection-dependent effects of recombinant adenovirus carrying HGF gene on the proliferation and osteogenic differentiation of human bone marrow mesenchymal stem cells. Int. J. Mol. Sci. 19, E734 (2018).
Karner, C. M., Lee, S. Y. & Long, F. Bmp induces osteoblast differentiation through both Smad4 and mTORC1 signaling. Mol. Cell. Biol. 37, e00253–00216 (2017).
Liu, X. et al. Lifelong accumulation of bone in mice lacking Pten in osteoblasts. Proc. Natl Acad. Sci. USA 104, 2259–2264 (2007).
Chen, J. et al. WNT7B promotes bone formation in part through mTORC1. PLoS Genet. 10, e1004145 (2014).
Fitter, S. et al. mTORC1 plays an important role in skeletal development by controlling preosteoblast differentiation. Mol. Cell Biol. 37, e00668–00616 (2017).
Singha, U. K. et al. Rapamycin inhibits osteoblast proliferation and differentiation in MC3T3-E1 cells and primary mouse bone marrow stromal cells. J. Cell Biochem. 103, 434–446 (2008).
Xian, L. et al. Matrix IGF-1 maintains bone mass by activation of mTOR in mesenchymal stem cells. Nat. Med. 18, 1095–1101 (2012).
Gharibi, B., Ghuman, M. S. & Hughes, F. J. Akt- and Erk-mediated regulation of proliferation and differentiation during PDGFRβ-induced MSC self-renewal. J. Cell Mol. Med. 16, 2789–2801 (2012).
Iyer, S., Viernes, D. R., Chisholm, J. D., Margulies, B. S. & Kerr, W. G. SHIP1 regulates MSC numbers and their osteolineage commitment by limiting induction of the PI3K/Akt/β-catenin/Id2 axis. Stem Cells Dev. 23, 2336–2351 (2014).
Li, C. J. et al. MicroRNA-188 regulates age-related switch between osteoblast and adipocyte differentiation. J. Clin. Invest. 125, 1509–1522 (2015).
Lai, P. et al. Loss of rictor with aging in osteoblasts promotes age-related bone loss. Cell Death Dis. 7, e2408–e2408 (2016).
Rossa, C. Jr, Liu, M., Patil, C. & Kirkwood, K. L. MKK3/6-p38 MAPK negatively regulates murine MMP-13 gene expression induced by IL-1beta and TNF-alpha in immortalized periodontal ligament fibroblasts. Matrix Biol. 24, 478–488 (2005).
Yang, H. et al. Comparison of mesenchymal stem cells derived from gingival tissue and periodontal ligament in different incubation conditions. Biomaterials 34, 7033–7047 (2013).
Acknowledgments
We acknowledge the funding support from the NSFC (Nature Science Foundation of China for Dr. Fa-Ming Chen from 2016 to 2020; No. 81530050) and Shaanxi Key Scientific and Technological Innovation Team (2017KCT-32).
Author information
Authors and Affiliations
Contributions
X.-Y.X. and X.-T.H. devised and performed the research, collected and analyzed the data, and drafted and revised the manuscript. F.-M.C. devised the research, provided funding, integrated the data and revised the manuscript. X.L., J.W., Y.X., and Y.-Z.T. conducted the study and helped collect the data and write the manuscript. All co-authors contributed to discussions throughout the study and approved the final version of the manuscript for publication.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Edited by Y. Shi
This article has been retracted. Please see the retraction notice for more detail: https://doi.org/10.1038/s41419-023-06390-y
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Xu, XY., He, XT., Wang, J. et al. RETRACTED ARTICLE: Role of the P2X7 receptor in inflammation-mediated changes in the osteogenesis of periodontal ligament stem cells. Cell Death Dis 10, 20 (2019). https://doi.org/10.1038/s41419-018-1253-y
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41419-018-1253-y
This article is cited by
-
Translating proteome and transcriptome dynamics of periodontal ligament stem cell-derived secretome/conditioned medium in an in vitro model of periodontitis
BMC Oral Health (2024)
-
P2X7Rs: new therapeutic targets for osteoporosis
Purinergic Signalling (2023)
-
Methylated MFGE8 Promotes Early Brain Injury After Subarachnoid Hemorrhage and Inhibiting Autophagy of Nerve Cell
Translational Stroke Research (2023)
-
LncRNA GACAT2 binds with protein PKM1/2 to regulate cell mitochondrial function and cementogenesis in an inflammatory environment
Bone Research (2022)
-
Moderate-intensity exercise alleviates pyroptosis by promoting autophagy in osteoarthritis via the P2X7/AMPK/mTOR axis
Cell Death Discovery (2021)
