
Abstract
The mammalian intestine harbors heterogeneous distribution of microbes among which specific taxa (e.g. Lactobacillus) dominate across mammals. Deterministic factors such as nutrient availability and utilization may affect microbial distributions. Due to physiological complexity, mechanisms linking nutrient utilization and the dominance of key taxa remain unclear. Lactobacillus amylovorus is a predominant species in the small intestine of pigs. Employing a pig model, we found that the small intestine was dominated by Lactobacillus and particularly L. amylovorus, and enriched with peptide-bound amino acids (PBAAs), all of which were further boosted after a peptide-rich diet. To investigate the bacterial growth dominance mechanism, a representative strain L. amylovorus S1 was isolated from the small intestine and anaerobically cultured in media with free amino acids or peptides as sole nitrogen sources. L. amylovorus S1 grew preferentially with peptide-rich rather than amino acid-rich substrates, as reflected by enhanced growth and PBAA utilization, and peptide transporter upregulations. Utilization of free amino acids (e.g. methionine, valine, lysine) and expressions of transporters and metabolic enzymes were enhanced simultaneously in peptide-rich substrate. Additionally, lactate was elevated in peptide-rich substrates while acetate in amino acid-rich substrates, indicating distinct metabolic patterns depending on substrate forms. These results suggest that an increased capability of utilizing PBAAs contributes to the dominance of L. amylovorus, indicating amino acid utilization as a deterministic factor affecting intestinal microbial distribution. These findings may provide new insights into the microbe-gut nutrition interplay and guidelines for dietary manipulations toward gut health especially small intestine health.
Similar content being viewed by others


Introduction
The ecological rationales that govern the microbial distribution are complex and involve multiple factors. One of the deterministic factors that affect the microbial distribution is the performance trait of bacteria, such as the bacterial capability of resource or nutrient utilization [1]. Lactobacillus is a dominant taxon in the small intestine across mammals, including pigs [2], rats [3], and rhesus macaque [4], with Lactobacillus amylovorus representing a predominant species [5, 6]. In the small intestine of piglets, L. amylovorus is an abundant species accounting for about 20%, and has been reported to exert carbohydrate metabolic functions [7] and beneficial effect by modulating gut immunity and inflammation [5]. Intestinal bacteria have different utilization preference for amino acids (AAs) and peptides [8,9,10]. A previous study revealed that the small intestinal bacteria of pigs could utilize peptides preferentially for their bacterial protein synthesis compared with free amino acids in vitro [11]. Whether the utilization preference for AAs and peptides may serve as a deterministic factor for Lactobacillus colonization in intestinal tract remains unclear. Given this evidence, it is of great relevance to understand the rationale that gives rise to the dominance of Lactobacillus species in the small intestine of some mammals.
To investigate the microbial preference for AAs and peptides in vivo, dietary inclusion of intact protein or hydrolyzed protein provides a nutritional approach. This approach enables to supply diets that have equal nitrogen source but differ in AA and peptide contents. Protein hydrolysate, compared to intact protein, contains peptide-bound amino acids and other bioactive components that serve as a dietary ingredient in ways of affecting intestinal physiology. We have demonstrated that compared to intact casein, dietary casein hydrolysate stimulated gastric transit, leading to a higher carbohydrate availability in ileal digesta and changing microbial composition [12]. Compared with intact soy protein, soy protein hydrolysates could facilitate the abundance of L. amylovorus in vitro [13]. Another study found that hydrolyzed casein-based diet increased the Lactobacillus abundance in the feces of rats [14].
In the present study, we question how Lactobacillus dominates in the intestine and whether nutrient utilization preference may contribute to the dominance. Employing a pig model, this study proved that dietary casein hydrolysate created a microenvironment with peptide-rich substrate and increased Lactobacillus, particularly in the small intestine. Within Lactobacillus, L. amylovorus was the most dominant species and expanded by peptide-rich substrate. We isolated L. amylovorus S1 from the small intestine of piglets as a model microorganism. Whole-genome resequencing and computational metabolism reconstruction provide new insight into its utilization of peptides and amino acids. To further investigate the rationale behind peptide/amino acid utilization, L. amylovorus S1 was cultured in medium containing substrate enriched with peptides or amino acids. Results showed a nutrient preference toward peptides, which may be a driving force of dominance of L. amylovorus. The approach employed here provides a strategy that can be applied to investigate the mechanisms driving to dominance of other key taxa in the small intestine. This study brings new insights into the dominance of L. amylovorus driven by AA utilization in microenvironment and highlights the important role of peptide-rich diet in the enrichment of lactic acid bacteria in the small intestine of pigs. These findings reinforce the importance of gut microbes in gut nutrition and may provide guidelines for dietary manipulations toward pig gut health especially the small intestine health.
Materials and methods
Experimental design and sample collection
To compare the effects of dietary protein form on the intestinal microbiota, intact casein and casein hydrolysate were selected considering their wide application as dietary ingredients. Casein hydrolysate was prepared by casein (used in Casein-intact group) hydrolysis with food-grade trypsin (Haibo Biotech Inc., Qingdao, China) and the resulting hydrolyzed casein (Casein-pep) was subsequently spray-dried to produce a dried powder. The proportions of AAs and peptides in the casein hydrolysate were 46% and 54%, respectively.
The animal experiment was approved by the Animal Care and Use Committee of Nanjing Agricultural University, in compliance with the Regulations for the Administration of Affairs Concerning Experimental Animals of China. After a 7 day period acclimatization, a total of sixteen cross-bred (Duroc × Landrace × Yorkshire) castrated male pigs (63 ± 2 days of age, average body weight 19.09 ± 0.61 kg) were subjected to corn-fish meal diet containing either intact casein (n = 8) or hydrolyzed casein (n = 8), at a content of 45 g per 1 kg diet. The nutritional composition of the two diets were formulated equally (Table S1). The pigs were housed individually in stainless steel metabolism cages and maintained at 24 ± 1 °C with free access to water. Pigs were offered equal amount of meal twice daily to ensure the equal dietary intake. The body weight was recorded and used for calculating the average daily weight gain. The health of each pig was closely monitored throughout the experiment. On day 29 of the experiment, all pigs were anesthetized using an intravenous injection of 4% sodium pentobarbital solution (40 mg/kg body weight) and sacrificed by exsanguination. The intestinal tract was removed immediately. The luminal digesta from the middle section of each intestinal segment (duodenum, jejunum, ileum, and colon) were collected and snap-frozen by liquid nitrogen immediately after sampling to avoid post mortem changes, and then transferred and stored at −80 °C until analysis.
DNA extraction, sequencing, and bioinformatics
Total genomic DNA from 0.3 g digesta of duodenum, jejunum, ileum, and colon, were extracted per sample using the DNA Stool Mini Kit (QIAGEN, Hilden, Germany) following the manufacturer’s protocols. The V3–V4 region of the bacterial 16S rRNA gene was amplified by PCR using universal bacterial primers (F: 5′-barcode-TAC GGR AGG CAG CAG-3′; R: 5′-AGG GTA TCT AAT CCT-3′) [15], where the barcode is an eight-base sequence unique to each sample [16]. All the polymerase chain reaction (PCR) processes used a 20 μL TransStart Fastpfu DNA polymerase reaction system, which included 4 μL 5× FastPfu buffer, 2 μL 2.5 mmol/L dNTPs, 0.8 μL 5 μmol/L Forward primer, 0.8 μL Reverse primer, 0.4 μL FastPfu polymerase, and 10 ng of DNA. Amplification was performed as follows: the first pre-degeneration was performed at 95 °C for 5 min, followed by 27 cycles of denaturation (95 °C, 30 s). Annealing (55 °C, 30 s) and elongation (72 °C, 45 s) were performed before an extension was done for 10 min at 72 °C. Purified amplicons were pooled in equimolar concentrations and paired-end sequenced (2 × 250) on a MiSeq platform (Illumina Inc., CA, USA) according to the standard protocols at the Majorbio Bio-Pharm Technology Co., Ltd. (Shanghai, China).
Raw sequence data generated from 16S rRNA gene sequencing were demultiplexed, quality-filtered using QIIME (version 1.9.1). Bases with an average quality score more than 20 were retained. Operational taxonomic units (OTUs) were clustered with 97% similarity cutoff using UPARSE (version 7.1) and chimeric sequences were identified and removed using UCHIME version 4.1. Taxonomy assignment of OTUs was performed by matching the representative sequences to 16S rRNA gene database (SILVA version 138) using Ribosomal Database Project Classifier (version 2.2). Species richness estimator and diversity indices were calculated using QIIME (version 1.9.1, http://qiime.org/scripts/split_libraries_fastq.html). Rarefaction curves were generated using R software with ggplot2 (version 3.3.4; https://www.r-project.org/). Weighted UniFrac principal coordinate analysis (PCoA) based on OTUs was performed by using QIIME (version 1.9.1) in vegan R package (version 4.1.2). Differences in gut bacterial taxa were analyzed by Linear discriminant analysis (LDA) effect size (LEfSe) analysis [17], with LDA scores >2 as discriminant taxa. The relative abundance of different bacterial taxa was expressed as percentages. Graphing was performed using Graphpad Prism 8 (La Jolla, CA, USA) and R statistics (version 4.1.2) in ggplot2 package (version 3.3.4). The analysis of molecular variance (AMOVA) [18] was performed to compare the difference in composition of the intestinal microbiota between two groups by using mothur (version 1.1, https://www.mothur.org/). The relative abundance of different bacterial taxa was expressed as percentages.
Determination of total Lactobacillus population by qPCR assay
Quantities of Lactobacillus, L. amylovorus, L. paracasei, L. salivarius, L. reuteri and L. johnsonii in intestinal digesta were determined by qPCR. All samples were analyzed in triplicates. The plasmids containing insert of 16S rRNA genes were constructed using pMD18-T according to the kit instructions (D101A, Takara, Kusatsu, Japan). The target plasmids were diluted to a series of 10-fold to generate the standard curves. The qPCR assay was performed on a lightcycler 480 Real-time PCR system (Roche, Solna, Sweden) with 384-well plates, using SYBR Premix Ex Taq dye II (Takara, Kusatsu, Japan). PCR amplification condition was at 95 °C for 30 s, followed by 40 cycles of 95 °C for 10 s, 60 °C for 30 s. The copy number of target bacteria in digesta (copy/g) was quantified according to standard curves. The primer sequences of specific 16S rRNA genes [19] are listed in Table S2.
Isolation, purification and identification of L. amylovorus
The isolation, purification and identification of Lactobacillus were performed as previously described [20]. Briefly, luminal digesta was obtained from the small intestine of healthy pigs (Duroc × Landrace × Yorkshire). One gram digesta was suspended in sterile saline water (5 mL) and then serially diluted to 1:103, 1:105, and 1:107. The diluted samples were spread on MRS agar plates and cultured at 37 °C for 48 h under anoxic conditions until the isolated colonies formed. Well-isolated colonies were picked and further streaked into fresh MRS plates until a pure culture was obtained. Gram-staining, morphology, and molecular identification based on 16S rRNA gene sequencing were performed [20]. The identity of L. amylovorus S1 and gut bacteria (OTU1) in 16S rRNA gene sequence is 100% using MEGA 11 (Molecular Evolutionary Genetic Analysis) software (available at https://www.megasoftware.net). Phylogenetic tree was conducted by MEGA11 using a neighbor-joining method, with a bootstrap value of 1000.
Computational reconstruction of peptide and amino acid metabolism modules for L. amylovorus S1
The complete genome of L. amylovorus S1 was sequenced using HiSeq 4000 (Illumina Inc.) with paired-end modes and PacBio sequencing with Pacific Biosciences SMRT (PacBio RSII) at Biozeron Biotechnology Co., Ltd. (Shanghai, China). For genome assembly, the optimal results of the assembly were obtained by ABySS (version 2.0.2, http://www.bcgsc.ca/platform/bioinfo/software/abyss). PacBio-corrected long reads were then assembled by Canu (version 1.8, https://github.com/marbl/canu). Finally, GapCloser (version 1.12, https://sourceforge.net/projects/soapdenovo2/files/GapCloser/) was subsequently applied to fill up the remaining local inner gaps and correct the single base polymorphism. Gene prediction was performed using GeneMarkS (version 4.6b). The complete circle of the genome was drawn with Circos (version 0.64). The annotations of transporter proteins and amino acid metabolic enzymes were obtained against TCDB (Transporter Classification Database, http://www.tcdb.org/) and KEGG debase (http://www.genome.jp/kegg/). To gain better insights into amino acid utilization, we reconstructed the peptide and AA metabolism and transporter modules for L. amylovorus S1.
Prediction of the amino acids that can bind the oligopeptide transporter was performed using COACH protein-ligand binding prediction server [21]. 3D structure model of OptB protein was generated by I-TASSER [22] and by SPARKS-X [23]. The ligand binding site predictions were performed by TM-SITE and S-SITE by matching ligand-binding templates from the BioLip protein function database (version Apr 01, 2022) with the query structure and sequence [24]. The prediction for final ligand binding sites was generated using ConCavity method [25].
In vitro study of AA utilization by L. amylovorus S1
To investigate the utilization of AAs and peptides, we used the L. amylovorus S1 as a model microorganism for small intestinal L. amylovorus and analyzed its growth in MRS medium enriched with free amino acids or peptides in batch culture. Peptides and free amino acids were provided by casein acid hydrolysate (enriched with free amino acids; Casein-AA group) or casein enzymatic hydrolysate (enriched with peptides; Casein-PEP group) in equimolar quantities as the sole nitrogen source. Casein acid hydrolysate provided amino acid source, containing FAAs and PBAAs in 60.43 and 39.57%, respectively, while the casein enzymatic hydrolysate provided peptide source, containing PBAAs and FAAs in 71.95 and 27.80%, respectively. The AA concentrations in the medium before batch culture were also measured (Table S3). To balance the total amino acid concentration at the same level, different amounts of the casein hydrolysates were added, with the casein acid hydrolysate at 18 g/L and the casein enzymatic hydrolysate at 8.2 g/L in the medium. The solution was filter-sterilized (through 0.22 µm filter) into pre-gassed autoclaved serum bottle. The bacteria were grown anaerobically at 37 °C in a modified MRS medium, replacing the nitrogen source with casein acid hydrolysate or casein enzymatic hydrolysate. Samples of bacterial cultures were taken at 0, 6, 12, and 24 h after inoculation (n = 4 per time point per group) and stored immediately at −25 °C until use for the analysis.
Bacterial growth curve and qPCR of L. amylovorus S1 in culture medium
Portions of 200 µL culture medium were transferred in 96-well plates for plotting OD600 at 37 °C, and each sample was set up in triplicate wells. Standard growth curve of L. amylovorus S1 was produced by determination of OD value every 2 h during 16-h culture. The 16S rRNA gene copy number of L. amylovorus in culture medium (copy/mL) was determined and quantified following aforementioned procedures.
RNA extraction and quantitative reverse transcription PCR analysis of gene expression of bacterial transporters and enzymes
Total RNAs were extracted by TRIzol method using RNA extraction Kit (Aidlab, Beijing,China) and quantified using the Nanodrop 2000 (Thermo Fisher Scientific, MA, USA) and reverse transcribed into cDNAs using PrimeScrip RT Master Mix (Takara, Kusatsu, Japan) according to the manufacturer’s protocol. Specific primers (Table S2) were designed to target AA and peptide transporters (optB, potE, artJ, metN, lysXY, gluA and bcaP), and metabolic enzymes (bcat, metK, sda, dapA, dapB, ackA, and ldh). recA was used as a reference gene. The amplification was performed on QuantStudio 5 Real-Time PCR instrument (Applied Biosystems, CA, USA) using SYBR Green method. The gene expression level was determined by comparing to 0 h-Casein-AA group using 2−ΔΔCt method.
Amino acid measurement
Intestinal digesta and bacterial culture supernatant
The contents of free amino acids (FAAs) and total amino acids (TAAs) in the jejunal and ileal digesta and bacterial culture supernatant were measured according to the method of o-phthaldialdehyde with high performance liquid chromatography (Agilent 1220 Infinity LC system; Agilent Technologies Inc., CA, USA) [26]. For TAA measurement, the samples were hydrolyzed with 6 M H2SO4 at 110 °C for 24 h before analysis. The content of peptide-bound AAs was calculated as a difference of TAAs and FAAs.
Bacterial cells
Bacterial cells were precipitated from 4 mL of bacterial culture into anaerobic phosphate-buffered saline. The cells were then hydrolyzed with 6 M H2SO4 at 110 °C for 24 h and used for TAA measurement.
Calculation
The AA disappearance amount in culture medium was calculated as follows: DAt = AA0-AAt, where DAt is the disappearance amount at t h, AA0 is the AA concentration at 0 h, AAt is the AA concentration at t h.
Measurement of short-chain fatty acids (SCFAs), lactate, and glucose in bacterial culture medium
Concentrations of SCFAs were determined with gas chromatography as described in our previous study [27]. The concentration of lactate was determined by a commercial lactate assay kit (A019-2, Nanjing Jiancheng Bioengineering Institute, Nanjing, China). The glucose concentration was determined by a commercial glucose assay kit (F006-1-1, Nanjing Jiancheng Bioengineering Institute, Nanjing, China).
Statistical analysis
Bartlett’s test was used to determine whether variance differed between groups where significance levels were p < 0.05 using Graphpad Prism 8 (La Jolla, CA, USA). Comparisons of means were performed with Student’s t test when homogeneity of variance was true, and Mann–Whitney U-test when variances were not homogeneous. p < 0.05 was considered significant. For comparisons of bacterial taxa between two groups, p values were adjusted with a false discovery rate (FDR) analysis [28], limiting the overall FDR to 5% (q < 0.05). For amino acid contents in bacterial cells, a general linear model (SPSS 20.0; SPSS Inc., Chicago, IL, USA) was performed to analyze the data of bacterial amino acid contents, using the bacteria quantity at 12 h as a covariate to correct.
Results
All pigs remained healthy during the experimental period, and no clinical signs of diarrhea or health impairment were observed. In comparison with pigs fed on Casein-intact diet, those on Casein-pep diet had a significant higher average daily gain (Fig. S1).
Sequencing quality of 16S rRNA gene sequences
After trimming of sequence reads and quality filtering, 2,821,266 high-quality sequences (representing 88.90% of the total sequences) were acquired, with an average of 44,082 sequences per sample. Sequencing depth nearly reached the total microbial species richness, with almost all the rarefaction curves approaching the saturation plateau (Fig. S2). This observation was verified by the Good’s coverage of each sample which was more than 99.9%, implying that most of microbial diversity had been sufficiently captured.
Hydrolyzed casein changed microbiome structure depending on gut locations
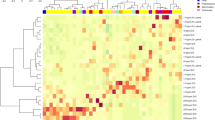
The weighted UniFrac PCoA analysis showed that there were differences of overall microbial composition between different intestinal segments (Fig. S3), which indicated a spatial heterogeneity of the microbiome across the intestine. Compared to Casein-intact, the Casein-pep significantly altered the microbiota composition in the lumen of jejunum, ileum, and colon (AMOVA p < 0.05, Fig. 1A), but not significantly in the duodenum. These observations suggest that the impact of Casein-pep, relative to Casein-intact, on the microbiota composition was different across gut locations.

A Principle-coordinate analysis by weighted UniFrac distance and the analysis of molecular variance. B The heatmap visualization of the relative abundance of top 30 genera. C Relative abundances of representative taxa. Abbreviations: Co colon, Du duodenum, Il ileum, Casein-intact intact casein group, Je jejunum, Casein-pep hydrolyzed casein group. *p < 0.05, **p < 0.01 between groups.
Hydrolyzed casein led to the dominance of Lactobacillus in the small intestine
At the taxa level, we observed the higher relative abundances of Lactobacillus (15–60%) and lower abundance of Escherichia in the jejunum in Casein-pep group than those in Casein-intact group (Fig. 1B, C). Moreover, the relative abundance of Megasphaera and Mitsuokella in the jejunum and Lactobacillus and Actinobacillus in the ileum were significantly higher in Casein-pep group than those in Casein-intact group (Fig. 1B). The relative abundance of Lactobacillus was also higher in the colon in Casein-pep group than that in Casein-intact group (Fig. 1B). Within the total Lactobacillus, the relative abundance of L. amylovorus was increased from 3.6% in Casein-intact group to 41.3% (mean value) in Casein-pep group in the jejunum (p < 0.05, Fig. 1C). Similar alterations of L. amylovorus were also observed in the ileum, while no difference for Limosilactobacillus reuteri. Regarding the strain diversity within L. amylovorus species, there are two OTUs (OTU1 and OTU42) that were close to L. amylovorus, among which, OTU1 showed 100% similarity to L. amylovorus with the relative abundance between 3.6% and 41.3% in the small intestine. OTU42 showed 99.77% similarity to L. amylovorus with relative abundance <0.001%. Casein-pep further reduced the relative abundance of Ligilactobacillus salivarius in the jejunum (Fig. 1C). Overall, it is evident that Casein-pep vs Casein-intact had the largest impact on the microbiota in the jejunum and ileum. Remarkably, the genus Lactobacillus was differentially affected along the whole intestine.
Hydrolyzed casein increased L. amylovorus and peptide-bound amino acids in the jejunal and ileal digesta compared to intact casein
To determine which group of Lactobacillus was specifically stimulated by the Casein-pep diet, qPCR was conducted to quantify the Lactobacillus, L. amylovorus, L. paracasei, L. salivarius, L. reuteri and L. johnsonii. Our results showed that compared with Casein-intact diet, Casein-pep diet significantly increased the quantity of Lactobacillus in the jejunum (p = 0.024, Fig. 2A). In the jejunum and ileum, the quantity of L. amylovorus increased by nearly 15-fold (p = 0.023) and 27-fold (p = 0.021) respectively, by Casein-pep diet compared to the Casein-intact diet, suggesting that Casein-pep diet could stimulate the growth of L. amylovorus relative to Casein-intact diet (Fig. 2A, B). Furthermore, L. amylovorus had the highest quantity among Lactobacillus spp. There was no difference in quantities of L. salivarius, L. reuteri and L. johnsonii between groups (p > 0.05); this may be due to the large individual variation in Casein-pep samples.

Quantitative real-time PCR measurement of bacterial quantities in the lumen of jejunum (A) and ileum (B). Concentrations of peptide-bound amino acids (PBAA) and free amino acids (FAA) in the jejunal (C) or ileal digesta (D). *P < 0.05 between groups.
Since Casein-pep and Casein-intact diets provide important nitrogen substrate for bacterial growth, we further determined the FAA and PBAA concentrations in the intestinal digesta. In the jejunum, the level of PBAA was significantly higher in Casein-pep group compared with Casein-intact group (p = 0.046, Fig. 2C), and in the ileum, the content of PBAA showed the increased tendency in Casein-pep group compared with Casein-intact group (p = 0.072, Fig. 2D). The level of FAA showed no difference between groups in both jejunum (p = 0.570) and ileum (p = 0.956). Taken together, these data demonstrated that Casein-pep diet increased the concentration of PBAAs and the quantity of L. amylovorus in the small intestinal digesta, indicating that peptide-rich substrate may lead to the growth predominance of L. amylovorus.
We further measured short-chain fatty acids as a reflection of microbial fermentation. In the jejunum, Casein-pep group had higher concentrations of butyrate than the Casein-intact group (p = 0.045, Fig. S4). No differences in acetate and propionate were observed between the two groups.
Identification of L. amylovorus S1 and its global amino acid utilization inferred from genomic features
The next step was to isolate L. amylovorus from the jejunum of the small intestine in order to determine how it was affected by the Casein-pep diet. By picking a single colony, we isolated a L. amylovorus strain S1 (Fig. 3A). The 16S rRNA gene sequence of L. amylovorus S1 were 100% identical to the representative 16S rRNA gene sequence of L. amylovorus from microbiome sequencing (Fig. 3B). To better understand the functional potential involved in AA and peptide utilization, whole genome sequencing was conducted. Nearly 3.05 GB raw data of L. amylovorus S1 sequence were generated by the HiSeq sequencing platform. The Q30 value of raw reads was 93.83%. After data QC and filtering, nearly 2.90 GB clean bases were obtained for genome assembly. Additionally, 464,347 nanopore reads (N50 = 11,193, totally 1.87 GB bases) were obtained. The complete genome sequence of L. amylovorus S1 consists of a circular chromosome (2,091,784 bp), and 2 circular plasmids (67,799 bp and 39,117 bp, respectively). The genome of L. amylovorus S1 contains 2232 protein-coding sequences and GC content is 38.33%, and harbors 65 tRNA genes and 15 rRNA operons. The genome-based annotation on metabolism and transport of AAs and peptides of L. amylovorus S1 were depicted (Fig. 3C). The annotation of enzymes involved in amino acid metabolism showed that L. amylovorus S1 strain possessed a number of enzymes involved in metabolism of methionine, aspartate, serine, glutamate, glycine, threonine, valine, isoleucine and leucine, such as oligopeptide ABC transporter permease protein OptB and branched-chain amino acid aminotransferase Bcat.

A Morphology of L. amylovorus S1. B Phylogenetic tree of L. amylovorus S1 inferred from the 16S rRNA gene sequences by MEGA 11. C Computational reconstruction of the peptide and AA transporters, and AA metabolic enzyme modules for L. amylovorus S1 based on genomic sequencing. The enzyme, substrate and product information in enzyme reactions were available by KEGG debase (Kyoto Encyclopedia of Genes and Genomes), and the transporter proteins of AA and peptide were obtained through TCDB (Transporter Classification Database).
Peptide-rich substrate increased the growth of L. amylovorus and the disappearance rate of PBAAs compared to AA-rich substrate
To study the growth of L. amylovorus S1 cultured in medium containing Casein-AA (FAA-rich) and Casein-PEP (PBAA-rich) as nitrogen sources, a growth curve was generated (Fig. 4A). Growth at both substrates followed the same exponential phase until 6 h and that stationary phase was reached at 6 h for Casein-AA and at 12 h for Casein-PEP. Based on the growth curve of L. amylovorus S1 in PBAAs-rich substrate, the time point of the sample collection were selected at 0 h (initial phase), 6 h (mid-exponential phase), and 12 h (stationary transition phase). The quantity of L. amylovorus S1 was also increased by 22.96% in PBAA-rich substrate as compared to that in FAA-rich substrate at 12 h (p < 0.001, Fig. 4B).

A Growth curve. B Quantitative measurement of L. amylovorus S1. C The disappearance amounts of free amino acids (FAA) and peptide-bound amino acids (PBAA) in the amino acid-rich (Casein-AA group) and peptide-rich (Casein-PEP group) substrates at 12 h after cultivation. D The cellular AA concentration in L. amylovorus S1 at 12 h. Asterisk indicates significance between groups. E The relative expression of optB (oligopeptide ABC transporter substrate binding protein). F The protein-ligand binding site prediction for OptB protein of L. amylovorus S1, showing the ligand binding sites for threonine (T58, T291), phenylalanine (F63, F423), valine (V60, V439), serine (S443), glycine (G441) and lysine (K511), which were obtained by COACH protein-ligand binding prediction server. G The disappearance rate of peptide-bound AAs in the cultivation at 6 h and 12 h. H A proposed model shows that the upregulation of OptB peptide transport accompanies with increased utilization of peptide-bound AAs that can bind OptB. *p < 0.05, **p < 0.01, ***p < 0.001 between groups.
To investigate the substrate utilization of L. amylovorus S1 for nitrogen sources, the bacterial utilization of PBAAs and FAAs were tested after in vitro cultivation (Fig. 4C). The 12-h disappearance amount of PBAAs was higher than that of FAAs in Casein-PEP group (p < 0.001), whereas the disappearance amounts of PBAAs and FAAs showed no obvious difference in Casein-AA group (p = 0.719). These results indicate that the growth of L. amylovorus S1 performs better with peptides than free AAs.
Peptide-rich substrate increased cellular AA profiles in L. amylovorus
Cellular AA profiles reflect the portion of AAs incorporated into bacterial cells. At 12 h, the contents of cellular phenylalanine (p = 0.008), arginine (p = 0.010), glycine (p = 0.020), isoleucine (p = 0.017), leucine (p = 0.003), lysine (p = 0.003), methionine (p = 0.003) and tyrosine (p = 0.026) were higher in peptide-rich substrate compared with AA-rich substrate. The contents of alanine (p = 0.113), aspartate (p = 0.087), serine (p = 0.133), threonine (p = 0.082), histidine (p = 0.175), glutamate (p = 0.205) and valine (p = 0.866) showed no difference between groups. These results indicate the increased incorporation of amino acids into bacterial cells when growing in PBAA-rich substrate.
Peptide-rich substrate increased bacterial peptide utilization
Oligopeptide ABC transporter-binding protein (Opt) family is crucial for peptides intake into bacterial cells [29]. The gene expression of oligopeptide ABC transporter optB was significantly upregulated in Casein-PEP group at 6 h (p < 0.001) and 12 h (p = 0.023) after culture (Fig. 4E). To investigate the AAs that may bind to OptB, we performed the protein-ligand binding site prediction. The structure of OptB was predicted by SPARKS-X with template 7ofwA, z-score of 22.820 (cutoff = 6.9). The prediction showed that OptB harbors ligand binding sites for threonine (T58, T291), phenylalanine (F63, F423), valine (V60, V439), serine (S443), glycine (G441) and lysine (K511) (Fig. 4F) with a confidence score of 0.80, indicating that these AAs may bind to OptB with high-affinity for transport.
To study if OptB-mediated transport of these AAs was affected by the substrate, we determined the disappearance rates of these AAs in peptide-bound form (i.e. PBAAs). The 6 h-disappearance rates of serine (p = 0.003) and valine (p = 0.003) were higher in Casein-PEP group, while that of the phenylalanine was significantly lower than in Casein-AA group (p = 0.036, Fig. 4G). As the growth at Casein-PEP was still in exponential phase, the 12 h-disappearance rates of threonine (p = 0.001), serine (p = 0.002), valine (p < 0.001) and lysine (p = 0.017) were higher in Casein-PEP group than in Casein-AA group, while glycine (p = 0.221) and phenylalanine (p = 0.207) showed no differences between groups. Thus, these results indicate that the upregulation of optB gene expression was accompanied with increased utilization of threonine, serine, valine and lysine in peptide-bound form that could bind OptB under peptide-rich than AA-rich substrates (Fig. 4H).
Peptide-rich substrate further increased bacterial AA utilization
At 12 h, the growth was in exponential phase in Casein-PEP while stationary phase in Casein-AA group (Fig. 4A). To study if AA utilization was altered, we further measured the 0–12 h-disappearance rate of FAAs. The disappearance rates of valine, leucine, methionine, serine, arginine, and lysine were significantly increased in Casein-PEP group compared with Casein-AA group (p < 0.05), while the aspartate (p = 0.072) and glutamate (p = 0.850) showed no significant difference between groups (Fig. 5A). The AA utilization depends on their transporters and metabolic enzymes. The expression of arginine transporter (artJ, p = 0.009), methionine transporter (metN, p = 0.002), lysine transporter (lysXY, p < 0.001) and glutamate transporter (gluA, p = 0.025) genes increased in Casein-PEP group at 12 h (Fig. 5A). For the AA metabolism enzymes, we found the upregulation of genes encoding AA metabolic enzymes toward branch-chain amino acids (bcat, p = 0.001), methionine (metK, p < 0.001), serine (sda, p = 0.050), aspartate (dapA, p < 0.001 and dapB, p < 0.001) in Casein-PEP group compared with Casein-AA group (Fig. 5B).

A The 12 h-disappearance rates of free AAs, and the gene expression of their corresponding transporters and metabolic enzymes. B The proposed model shows that the transport of Met and Arg are mainly through their specific transporters, while both OptB and LysXY are responsible for Lys transport, resulting in the increased concentrations of Met, Arg, and Lys in bacterial cells of L. amylovorus S1. *p < 0.05, **p < 0.01, ***p < 0.001 between groups.
Peptide- and AA-rich substrate led to distinct fermentation patterns
To characterize the metabolic end products under AA-rich and peptide-rich substrates, we measured the short-chain fatty acids and lactate. The lactate production was higher in Casein-PEP at 24 h after culture (p < 0.001, Fig. 6A), accompanied with the upregulation of lactate dehydrogenase (ldh) gene expression compared with FAA-rich substrate at 6 h (p < 0.001), 12 h (p = 0.010) and 24 h (p = 0.026) (Fig. 6B). Corresponding to the increase of lactate, the pH value was lower in Casein-PEP at 24 h after culture (p = 0.018, Fig. 6C). As the microbial fermentation product, acetate accounts for the high proportion among microbial-derived SCFAs, and our data showed that acetate was the only bacterial metabolite detected among SCFAs during 24 h-culture. The concentration of acetate was lower in Casein-PEP at 24 h after culture (p = 0.009, Fig. 6D), with the downregulation of acetate kinase (ackA) gene expression in Casein-PEP compared with Casein-AA substrate at 24 h (p = 0.046) (Fig. 6E).

A Lactate concentrations. B Relative gene expressions of lactate dehydrogenase (ldh). C pH measurement. D Acetate concentrations. E Relative gene expressions of acetate kinase (ackA). F Glucose concentrations. G Summary of the alterations in metabolite patterns. *p < 0.05, **p < 0.01, ***p < 0.001 between groups.
In addition to amino acids, glucose concentrations also affect fermentation patterns. To determine if glucose may be a confounding factor, we further measured the glucose concentration and found that the glucose concentration showed no obvious difference (Fig. 6F), indicating the difference in production dominance of lactate and acetate may be mainly driven by the amino acid and peptide content in different substrates (Fig. 6G).
Discussion
In the present study, we found that peptide-rich hydrolyzed casein could lead to growth dominance of L. amylovorus as well as increased peptide-bound AAs in the small intestine. Our in vitro study using L. amylovorus S1 as a model microorganism further validated that L. amylovorus had nutrient preference toward PBAA-rich substrate by enhancing peptide and amino acid utilization (Fig. 7). These findings support the hypothesis that the bacterial capability of resource or nutrient utilization could serve as a deterministic factor that drives the dominance of specific bacteria in the intestine. As an abundant species accounting for about 20% in the small intestine of piglets, L. amylovorus has been proved to exert carbohydrate metabolic functions [7] and favorable effect by modulating gut immunity and inflammation [5]. This study brings new insights into the dominance of L. amylovorus driven by AA utilization in microenvironment and highlights the important role of peptide-rich diet in the enrichment of lactic acid bacteria, such as L. amylovorus in the small intestine of pigs. These findings may provide guidelines for dietary manipulations toward gut health especially small intestine health.

A greater capability to utilize peptides than amino acids may contribute to the growth preference and dominance of L. amylovorus in peptide-rich substrate relative to amino acid-rich substrate.
Peptide-rich diet enriched Lactobacillus species in the small intestine
In the present study, the genus Lactobacillus was dominant in the small intestine of control animals with L. amylovorus as most dominant representative species under physiological condition. Dietary protein undergoes enzymatic digestion and subsequently releases peptides to the small intestine for bacterial and host metabolism. Given that AA and peptide utilization could be a survive strategy for bacteria in the intestine [30, 31], it is reasonable to speculate that the colonization dominance of Lactobacillus (particularly L. amylovorus) in the small intestine may be due to its nutrient requirement and substrate preference for peptides than AAs.
Interestingly, Lactobacillus, as well as the specific taxon L. amylovorus, was further boosted in the small intestine after casein hydrolysate inclusion in the diet, as well as the concentration of PBAA. Relative to the intact casein, the casein hydrolysate contains more readily available peptides, which provides a peptide-rich environment that mimics the physiological condition. Therefore, it is likely that a peptide-rich environment may be related to the growth dominance of L. amylovorus. Lactobacillus species could preferentially utilize the peptides than intact protein [32,33,34], due to the kinetic advantage of peptide-uptake system [33]. Two independent studies reported that extensively hydrolyzed protein could promote the growth of Lactobacillus species in vitro as compared to the intact protein [13, 32].
In the present study, with the enhancement of Lactobacillus after casein hydrolysate feeding, the concentration of lactate and the abundance of Megasphaera were also increased. Species of Megasphaera have been described to use lactate resulting in the production of butyrate and therefore, it could convert the lactate produced by Lactobacillus to butyrate. A cross-feeding has been demonstrated between Megasphaera and Lactobacillus in the hindgut of pigs [35]. Therefore, it is reasonable to speculate that there is a cross-feeding relationship between Lactobacillus and Megasphaera in the small intestine of pigs in response to peptide-rich hydrolyzed casein.
Another evident finding is that peptide-rich substrates exert greater impact in the small intestine than in the colon. Intestinal locations vary in nutrient availability and physiological conditions [36, 37]. Jejunum and ileum are major sites for the digestion and absorption of protein and amino acids, yet less well studied so far. These results highlight the relevance of small intestinal microbiota in protein nutrition.
After hydrolyzed casein feeding, Lactobacillus abundance and lactate concentration increased in the small intestine, which enhanced the ghrelin signal and development of interstitial cells of Cajal, accompanied by the increased ileal flows of nutrients and higher carbohydrate availability to support the bacterial activity [12]. This phenotype indicates the metabolic interaction between host and gut microbes after hydrolyzed casein feeding. In the present study, hydrolyzed casein also increased butyrate concentration in the small intestine. Butyrate is well known as an inhibitor of histone deacetylase that could affect the intestinal macrophage function [38]. Whether an increase in butyrate may exert epigenetic modification on host functions remains unknown, which can be investigated in future study.
A stronger capacity of utilizing peptide-bound amino acids contributes to the dominance of Lactobacillus amylovorus
We observed that the L. amylovorus S1 strain prefers the utilization of peptides for bacterial protein synthesis. Most of bacterial cellular AAs in L. amylovorus S1 were higher in peptide-rich than AA-rich substrate after 12 h-culture, together with increased utilization of peptide-bound amino acids, supporting the growth dominance of L. amylovorus S1 under peptide-rich substrate both in vivo and in vitro. Consistent with our findings, a previous study revealed that the small intestinal bacteria of pigs could utilize peptides preferentially for their bacterial protein synthesis compared with free amino acids in vitro [11]. Studies in ruminants also suggest that the ruminal bacteria prefer to assimilate more nitrogen source in the peptide-rich substrate for growth than free amino acid [39,40,41]. The utilization preference toward peptides may be an important performance trait of bacteria to survive in the intestine.
The growth curve of the isolated strain L. amylovorus S1 in peptide-rich substrate showed the longer exponential phase that achieved a higher OD value compared with AA-rich substrate. This difference may be due to the different strategies for bacterial AA and peptide utilization. AAs and peptides can be directly incorporated into bacterial cells as protein building blocks, and their transport systems are independent [42, 43]. However, peptide transport system was reported to be more kinetic advantages in comparison with those for free amino acids [33]; this in the present study allows the rapid uptake of peptides and protein synthesis by L. amylovorus S1 for proliferation, which contributed to the extended exponential phase under peptide-rich substrate.
An increase in the transport of peptides may account for the preference for peptide-rich substrate. Based on the genomic annotation, L. amylovorus S1 is equipped with peptide uptake system, allowing it to exert transport and utilization of peptides. Furthermore, the gene expression of peptide transporter optB was upregulated in peptide-rich substrate. The expression of bacterial peptide transporters could be controlled by the peptide concentration in medium [44, 45]. In Lactococcus lactis, peptide transporter optB gene expression was also upregulated in response to an increase in peptide levels [46]. In addition, serine and lysine could also be transported by OptB. The utilization of serine and lysine in the peptide form was also enhanced by peptide-rich substrate. Taken together, both the in vivo and in vitro results suggest that the peptide utilization by OptB may serve as an important strategy that allows the growth dominance of L. amylovorus.
Utilization of free amino acids was enhanced simultaneously with peptide utilization in peptide-rich substrate
In addition to the peptide transport, expressions of free amino acid transporters and their metabolic enzymes were also upregulated in peptide-rich substrate. L. amylovorus S1 harbors specific transporter MetN, ArtJ and LysXY, which play an important role in AA transport [47,48,49]. metN, encoding components of an ABC-type methionine transporter, is reported to mediate the high-affinity uptake of methionine [50]. Escherichia coli lacking MetN is unable to transport methionine, resulting in the decreased viability [51]. The ABC transporter encoded by artJ could specially bind arginine with high affinity [52]. The overproduction of artJ stimulated the arginine uptake by E. coli [47]. lysXY encodes a putative lysine uptake system that was reported as a new lysine transporter in Bacillus subtilis [49]. In this study, the upregulation of metN, artJ and lysXY, with the increased disappearance rates of methionine, arginine and lysine in peptide-rich substrate, may lead to the increased contents of these AAs in bacterial cells.
Enzymes responsible for amino acid metabolism were also upregulated. L. amylovorus S1 harbored genes encoding S-adenosylmethionine synthetase (MetK), serine dehydratase (Sda) and branch-chain amino acid aminotransferase (Bcat). Yvon and colleagues demonstrated that bcat transcription could be repressed by free AAs in L. lactis [53]. The present results showed that the increased utilizations of methionine, serine, valine and leucine were accompanied with the upregulation of metK, sda and bcat expression in peptide-rich compared to AA-rich substrates.
Distinct metabolic patterns of L. amylovorus in AA-rich and peptide-rich substrates
Metabolic patterns reflect the microbial physiology under specific conditions. In the present study, L. amylovorus S1 showed a typical heterolactic fermentation, with lactate and acetate as major products. However, the peptide-rich substrate enhanced lactate production while the AA-rich substrate enhanced acetate production at the end of fermentation. The mechanism behind is still unclear, but might be involved in the expression of enzymes, such as the upregulation of ldh (lactate production) and downregulation of ackA (acetate production) in peptide-rich substrate relative to AA-rich substrate. Another reason may be that the growth in peptide-rich substrate had longer exponential period than that in AA-rich substrate, which could lead to the accumulation of lactate. Consistent with the in vitro findings, an earlier in vivo study has demonstrated that lactate level was increased by the diet supplemented with hydrolyzed casein as compared with intact casein in pig small intestine [12]. Our result is similar to an in vitro study revealed that soybean oligopeptide can be preferentially utilized by L. acidophilus, but there was less acetate produced from soybean oligopeptide in comparison with pre-digested soybean oligopeptide by L. acidophilus at 48-h cultivation [54]. In the present study, the major substrates for L. amylovorus S1 metabolism are glucose and AA/peptide-rich substrate. Since there is no difference in glucose concentration, we suppose that the AA/peptide substrate was probably a major factor affecting the metabolism of L. amylovorus S1.
The animal gut harbors complex microbiota. The present study demonstrated that some of the in vivo observations could be explained by studying one L. amylovorus isolate from the small intestine. However, it has to be realized that using the single strain setup in vitro may potentially underestimate the intestinal complexity. Approaches using co-cultures with for example L. amylovorus and a Megaspaera strain or synthetic cultures with multiple strains may be used in future studies to consider the intestinal complexity. Further, it is of great interest for future studies using a hologenomic approach [55, 56] to characterize the host and microbial activity, and integrating metagenomics, transcriptomics and metabolomics platform as this would gain deeper insights into the microbe-host metabolic interaction in vivo. Furthermore, a longitudinal study (e.g. cannulated pig model) could be a great follow-up of the presented cross-sectional study in future to monitor the temporal dynamics of microbiota in the small intestine, allowing a better understanding of the microbiota turnover and compositionality changes over time due to the hydrolyzed casein feeding.
Conclusion
In summary, we conclude that a greater capability to utilize peptides than amino acids allows the dominance of Lactobacillus amylovorus in the small intestine. This feature could be reflected by their metabolic capability toward peptides rather than amino acids. Thus, these evidences support a long-standing ecological theory elucidating the nutrient utilization as an important deterministic factor in shaping the microbial distribution. Additionally, the present study provides a strategic approach to explain the performance of key taxa in the small intestine, which could be further applied to other microbes and investigated in future. Considering that a peptide-rich intestinal microenvironment provides competitive advantage for Lactobacillus and Lactobacillus amylovorus, these evidences implicate the relevance of nutritional manipulation to regulate intestinal microenvironment by peptide-rich diet.
Data availability
The 16S rRNA gene sequence of L. amylovorus S1 has been submitted to NCBI under the accession number MT525371. The high-throughput sequencing data are available under PRJNA796201 within NCBI Sequence Read Archive. The complete sequence of L. amylovorus S1 is available at GenBank under the accession number: CP090603 (chromosome), CP090604 (plasmid 1) and CP090605 (plasmid 2).
References
Chen YJ, Leung PM, Wood JL, Bay SK, Hugenholtz P, Kessler AJ, et al. Metabolic flexibility allows bacterial habitat generalists to become dominant in a frequently disturbed ecosystem. ISME J. 2021;15:2986–3004.
Mu C, Yang Y, Su Y, Zoetendal EG, Zhu W. Differences in microbiota membership along the gastrointestinal tract of piglets and their differential alterations following an early-life antibiotic intervention. Front Microbiol. 2017;8:797.
Li D, Chen H, Mao B, Yang Q, Zhao J, Gu Z, et al. Microbial biogeography and core microbiota of the rat digestive tract. Sci Rep. 2017;8:45840.
Yasuda K, Oh K, Ren B, Tickle TL, Franzosa EA, Wachtman LM, et al. Biogeography of the intestinal mucosal and lumenal microbiome in the rhesus macaque. Cell Host Microbe. 2015;17:385–91.
Hynonen U, Kant R, Lahteinen T, Pietila TE, Beganovic J, Smidt H, et al. Functional characterization of probiotic surface layer protein-carrying Lactobacillus amylovorus strains. BMC Microbiol. 2014;14:199.
Marti R, Dabert P, Ziebal C, Pourcher A-M. Evaluation of Lactobacillus sobrius/L. amylovorus as a new microbial marker of pig manure. Appl Environ Microbiol. 2010;76:1456–61.
Konstantinov SR, Awati AA, Williams BA, Miller BG, Jones P, Stokes CR, et al. Post-natal development of the porcine microbiota composition and activities. Environ Microbiol. 2006;8:1191–9.
Dai Z, Zhang J, Wu G, Zhu W. Utilization of amino acids by bacteria from the pig small intestine. Amino Acids. 2010;39:1201–15.
Wallace RJ. Ruminal microbial metabolism of peptides and amino acids. J Nutr. 1996;126:1326S–34S.
Yang YX, Dai ZL, Zhu WY. Important impacts of intestinal bacteria on utilization of dietary amino acids in pigs. Amino Acids. 2014;46:2489–501.
Liu J, Mu C, Yu K, Zhu W. Effect of two different casein hydrolysates on small intestinal bacteria of growing pigs. Acta Microbiol Sin. 2018;58:63–72.
Shen J, Mu C, Wang H, Huang Z, Yu K, Zoetendal EG, et al. Stimulation of gastric transit function driven by hydrolyzed casein increases small intestinal carbohydrate availability and its microbial metabolism. Mol Nutr Food Res. 2020;64:2000250.
Hsieh CM, Yang F-C, Iannotti EL. The effect of soy protein hydrolyzates on fermentation by Lactobacillus amylovorus. Process Biochem. 1999;34:173–9.
Visser J, Bos N, Harthoorn L, Stellaard F, Beijer‐Liefers S, Rozing J, et al. Potential mechanisms explaining why hydrolyzed casein‐based diets outclass single amino acid‐based diets in the prevention of autoimmune diabetes in diabetes‐prone BB rats. Diabetes/Metab Res Rev. 2012;28:505–13.
Singh KM, Jisha TK, Reddy B, Parmar N, Patel A, Patel AK, et al. Microbial profiles of liquid and solid fraction associated biomaterial in buffalo rumen fed green and dry roughage diets by tagged 16S rRNA gene pyrosequencing. Mol Biol Rep. 2015;42:95–103.
Fadrosh DW, Ma B, Gajer P, Sengamalay N, Ott S, Brotman RM, et al. An improved dual-indexing approach for multiplexed 16S rRNA gene sequencing on the Illumina MiSeq platform. Microbiome. 2014;2:6.
Segata N, Izard J, Waldron L, Gevers D, Miropolsky L, Garrett WS, et al. Metagenomic biomarker discovery and explanation. Genome Biol. 2011;12:R60.
Schloss PD, Westcott SL, Ryabin T, Hall JR, Hartmann M, Hollister EB, et al. Introducing mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl Environ Microbiol. 2009;75:7537–41.
Kim E, Yang S-M, Lim B, Park SH, Rackerby B, Kim H-Y. Design of PCR assays to specifically detect and identify 37 Lactobacillus species in a single 96 well plate. BMC Microbiol. 2020;20:96.
Chen L, Luo Y, Wang H, Liu S, Shen Y, Wang M. Effects of glucose and starch on lactate production by newly isolated Streptococcus bovis S1 from Saanen Goats. Appl Environ Microbiol. 2016;82:5982–9.
Yang J, Roy A, Zhang Y. Protein–ligand binding site recognition using complementary binding-specific substructure comparison and sequence profile alignment. Bioinformatics. 2013;29:2588–95.
Zheng W, Zhang C, Li Y, Pearce R, Bell EW, Zhang Y. Folding non-homologous proteins by coupling deep-learning contact maps with I-TASSER assembly simulations. Cell Rep Methods 2021;1:100014.
Yang Y, Faraggi E, Zhao H, Zhou Y. Improving protein fold recognition and template-based modeling by employing probabilistic-based matching between predicted one-dimensional structural properties of query and corresponding native properties of templates. Bioinformatics. 2011;27:2076–82.
Yang J, Roy A, Zhang Y. BioLiP: a semi-manually curated database for biologically relevant ligand-protein interactions. Nucleic Acids Res. 2013;41:D1096–103.
Capra JA, Laskowski RA, Thornton JM, Singh M, Funkhouser TA. Predicting protein ligand binding sites by combining evolutionary sequence conservation and 3D structure. PLoS Comput Biol. 2009;5:e1000585.
Dai Z, Wu Z, Jia S, Wu G. Analysis of amino acid composition in proteins of animal tissues and foods as pre-column o-phthaldialdehyde derivatives by HPLC with fluorescence detection. J Chromatogr B Anal Technol Biomed Life Sci. 2014;964:116–27.
Zhang C, Yu M, Yang Y, Mu C, Su Y, Zhu W. Differential effect of early antibiotic intervention on bacterial fermentation patterns and mucosal gene expression in the colon of pigs under diets with different protein levels. Appl Microbiol Biotechnol. 2017;101:2493–505.
Benjamini Y, Hochberg Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J R Stat Soc. 1995;57:289–300.
Lamarque M, Charbonnel P, Aubel D, Piard J-C, Atlan D, Juillard V. A multifunction ABC transporter (Opt) contributes to diversity of peptide uptake specificity within the genus Lactococcus. J Bacteriol. 2004;186:6492–500.
Dai Z, Wu G, Zhu W. Amino acid metabolism in intestinal bacteria: links between gut ecology and host health. Front Biosci. 2011;16:1768–86.
Morowitz MJ, Carlisle EM, Alverdy JC. Contributions of intestinal bacteria to nutrition and metabolism in the critically ill. Surgical Clin. 2011;91:771–85.
Zhang Q, Ren J, Zhao H, Zhao M, Xu J, Zhao Q. Influence of casein hydrolysates on the growth and lactic acid production of Lactobacillus delbrueckii subsp. bulgaricus and Streptococcus thermophilus. Int J Food Sci Technol. 2011;46:1014–20.
Davila A-M, Blachier F, Gotteland M, Andriamihaja M, Benetti P-H, Sanz Y, et al. Re-print of “Intestinal luminal nitrogen metabolism: Role of the gut microbiota and consequences for the host”. Pharmacol Res. 2013;69:114–26.
Pridmore RD, Berger B, Desiere F, Vilanova D, Barretto C, Pittet AC, et al. The genome sequence of the probiotic intestinal bacterium Lactobacillus johnsonii NCC 533. Proc Natl Acad Sci USA 2004;101:2512–7.
Michalak L, Gaby JC, Lagos L, La Rosa SL, Hvidsten TR, Tétard-Jones C, et al. Microbiota-directed fibre activates both targeted and secondary metabolic shifts in the distal gut. Nat Commun. 2020;11:5773.
Pereira FC, Berry D. Microbial nutrient niches in the gut. Environ Microbiol. 2017;19:1366–78.
Zhang L, Wu W, Lee YK, Xie J, Zhang H. Spatial heterogeneity and co-occurrence of mucosal and luminal microbiome across swine intestinal tract. Front Microbiol. 2018;9:48.
Chang PV, Hao L, Offermanns S, Medzhitov R. The microbial metabolite butyrate regulates intestinal macrophage function via histone deacetylase inhibition. Proc Natl Acad Sci USA. 2014;111:2247–52.
Argyle J, Baldwin R. Effects of amino acids and peptides on rumen microbial growth yields. J Dairy Sci. 1989;72:2017–27.
Chen G, Strobel H, Russell J, Sniffen C. Effect of hydrophobicity of utilization of peptides by ruminal bacteria in vitro. Appl Environ Microbiol. 1987;53:2021–5.
Soto RC, Muhammed SA, Newbold C, Stewart C, Wallace R. Influence of peptides, amino acids and urea on microbial activity in the rumen of sheep receiving grass hay and on the growth of rumen bacteria in vitro. Anim Feed Sci Technol. 1994;49:151–61.
Chalova VI, Sirsat SA, O’Bryan CA, Crandall PG, Ricke SC. Escherichia coli, an intestinal microorganism, as a biosensor for quantification of amino acid bioavailability. Sensors. 2009;9:7038–57.
Matthews D, Payne J. Peptides in the nutrition of microorganisms and peptides in relation to animal nutrition. In: Peptide transport in protein nutrition, New York, US: North-Holland Publishing Company; 1975. p. 1–60. vol. 37.
Sanz Y, Lanfermeijer FC, Renault P, Bolotin A, Konings WN, Poolman B. Genetic and functional characterization of dpp genes encoding a dipeptide transport system in Lactococcus lactis. Arch Microbiol. 2001;175:334–43.
Hartmann T, Cairns TC, Olbermann P, Morschhäuser J, Bignell EM, Krappmann S. Oligopeptide transport and regulation of extracellular proteolysis are required for growth of Aspergillus fumigatus on complex substrates but not for virulence. Mol Microbiol. 2011;82:917–35.
Dressaire C, Redon E, Gitton C, Loubière P, Monnet V, Cocaign-Bousquet M. Investigation of the adaptation of Lactococcus lactis to isoleucine starvation integrating dynamic transcriptome and proteome information. Microb Cell Factories. 2011;10:S18.
Wissenbach U, Six S, Bongaerts J, Ternes D, Steinwachs S, Unden G. A third periplasmic transport system for l‐arginine in Escherichia coli: molecular characterization of the artPIQMJ genes, arginine binding and transport. Mol Microbiol. 1995;17:675–86.
Merlin C, Gardiner G, Durand S, Masters M. The Escherichia coli metD locus encodes an ABC transporter which includes Abc (MetN), YaeE (MetI), and YaeC (MetQ). J Bacteriol. 2002;184:5513–7.
Rodionov DA, Vitreschak AG, Mironov AA, Gelfand MS. Regulation of lysine biosynthesis and transport genes in bacteria: yet another RNA riboswitch? Nucleic acids Res. 2003;31:6748–57.
Nguyen PT, Lai JY, Lee AT, Kaiser JT, Rees DC. Noncanonical role for the binding protein in substrate uptake by the MetNI methionine ATP Binding Cassette (ABC) transporter. Proc Natl Acad Sci USA. 2018;115:E10596–604.
Flatley J, Barrett J, Pullan ST, Hughes MN, Green J, Poole RK. Transcriptional responses of Escherichia coli to S-nitrosoglutathione under defined chemostat conditions reveal major changes in methionine biosynthesis. J Biol Chem. 2005;280:10065–72.
Caldara M, Charlier D, Cunin R. The arginine regulon of Escherichia coli: whole-system transcriptome analysis discovers new genes and provides an integrated view of arginine regulation. Microbiology. 2006;152:3343–54.
Yvon M, Chambellon E, Bolotin A, Roudot-Algaron F. Characterization and role of the branched-chain aminotransferase (BcaT) isolated from Lactococcus lactis subsp. cremoris NCDO 763. Appl Environ Microbiol. 2000;66:571–7.
Li W, Zhang Y, Li H, Zhang C, Zhang J, Uddin J, et al. Effect of soybean oligopeptide on the growth and metabolism of Lactobacillus acidophilus JCM 1132. RSC Adv. 2020;10:16737–48.
Alberdi A, Andersen SB, Limborg MT, Dunn RR, Gilbert MTP. Disentangling host–microbiota complexity through hologenomics. Nat Rev Genet. 2022;23:281–97.
Bordenstein SR, Theis KR. Host biology in light of the microbiome: ten principles of holobionts and hologenomes. PLoS Biol. 2015;13:e1002226.
Acknowledgements
This work was supported by National Natural Science Foundation of China (32030104, 31902166).
Author information
Authors and Affiliations
Contributions
WZ conceived, designed, supervised the study and proposed the manuscript strategy. YJ conducted the experimental work and performed statistical tests with guidance from CM and EGZ. CM wrote the manuscript draft with revisions and edits by YJ, EGZ and WZ. WZ and CM finalized the manuscript. HW and JS conducted the animal study. All the authors have read and approved the final manuscript for publication.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
About this article

Cite this article
Jing, Y., Mu, C., Wang, H. et al. Amino acid utilization allows intestinal dominance of Lactobacillus amylovorus. ISME J 16, 2491–2502 (2022). https://doi.org/10.1038/s41396-022-01287-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41396-022-01287-8
This article is cited by
-
Ensilage using Leuconostoc lactis and Weissella confusa reduces microbial risk and enhances hygienic quality of whole-crop corn
Chemical and Biological Technologies in Agriculture (2024)
-
Regulation of serotonin production by specific microbes from piglet gut
Journal of Animal Science and Biotechnology (2023)
