
Abstract
Microbes often secrete high levels of quorum sensing (QS) autoinducers into the environment to coordinate gene expression and biofilm formation, but risk detection and subsequent predation by bacterivorous predators. With such prominent signaling molecules acting as chemoattractants that diffuse into the environment at alarmingly high concentrations, it is unclear if bacterial cells can mask their chemical trails from predator detection. Here, we describe a microbial-based anti-detection adaptation, termed as “biofilm cloak”, where the biofilm prey produced biofilm matrix exopolysaccharides that “locked” and reduced the leaching of autoinducers into the milieu, thereby concealing their trails to the detection by the bacterivorous Caenorhabditis elegans nematode. The exopolysaccharides act as common good for the non-producers to hide their autoinducers from predator detection. Deficiency in chemosensory gene odr-10 in mutant animals abrogated their ability to detect autoinducers and migrate toward their prey in a directed manner, which led to lower population growth rate of animals. Hence, restriction of bacterial communication activities to the confinements of biofilms is a novel approach for predator evasion, which plays a fundamental role in shaping ecological dynamics of microbial communities and predator-prey interactions.
Similar content being viewed by others

Introduction
Anti-predator adaptations are mechanisms that assist the prey organisms in their struggle for survival against predators, where many are evasive in nature, such as camouflage and masquerade. In this pursuit, experimental model systems using microbes or higher organisms, have served as an important bridge between nature’s complexity and simplified theoretical studies [1, 2]. We and others had recently shown novel predator-prey interactions using the bacteria and nematode, Caenorhabditis elegans as model systems [3, 4], thus revealing novel insights into predator-prey relationships in the microbial world [5]. Predators such as C. elegans, can detect the attractive odorants released by bacteria, such as isoamyl alcohol and diacetyl, and locate their prey [6, 7]. C. elegans depends on its olfactory system to detect bacterial prey, where neuron pairs present in its head and tail are important for detection of chemical cues in the environment [8].
Bacteria spend nearly their entire lives in biofilms, which are multicellular communities encased in self-produced exopolymeric matrix. To coordinate biofilm formation, bacteria employ quorum sensing (QS) to crosstalk among themselves via the release of QS autoinducers. One classical example is Pseudomonas aeruginosa, which employs both homoserine-based (las and rhl) and quinolone-based (pqs and iqs) QS systems for intercellular communication and subsequent biofilm formation. However, autoinducers could be detected easily by predators and immune cells [9, 10], thus posing a dilemma for microbes in choosing between biofilm formation and predator evasion.
Here, we showed that the biofilm matrix acted as cloak to hide QS signals and prevent predator detection. Specifically, exopolysaccharides released by P. aeruginosa biofilms was crucial in “locking” N-butanoyl-l-homoserine lactone (BHLs) within the biofilms to reduce their leaching into the environment, thereby preventing detection by C. elegans. The “biofilm cloaking” feature was a common good for non-exopolysaccharide producers and other bacterial species. Correspondingly, odr-10, a C. elegans chemosensory gene, was crucial in the detection of autoinducers released from biofilms. Hence, our work demonstrated a hitherto described biofilm cloaking phenotype which plays a fundamental role in shaping ecological dynamics of microbial communities and predator-prey interactions.
Results
C. elegans cannot detect exopolysaccharide-producing biofilms
We first tested if C. elegans would prefer the bacteria deficient in biofilm matrix production over wild-type bacteria in a typical choice assay. Since P. aeruginosa mainly produce Pel and Psl exopolysaccharides in its biofilms [11], we evaluated the conventional choice assay which allowed the animals to choose between exopolysaccharide deficient (ΔpelA, ΔpslBCD, or ΔpelAΔpslBCD) biofilms and PAO1 wild-type biofilm. We found that the exopolysaccharide mutant biofilms were preferred by the animals than wild-type PAO1 biofilm (Supplementary Fig. S1). As proof-of-concept showing if C. elegans can detect the metabolites released by the biofilms into the environment, we employed a modified assay of chemotaxis, where secreted biofilm metabolites were used as chemoattractant for C. elegans detection (Fig. 1A). Briefly, we first grew P. aeruginosa biofilms on 0.2-µm filter overlaid nematode growth media (NGM) agar at opposite ends of the petri dish for 24 h, allowing the secreted metabolites to seep into the agar. We then removed the filter (with biofilms attached) from the agar to eliminate the possibility that C. elegans could detect bacterial biofilm cells directly, followed by placement of C. elegans in the middle of agar plate. This enabled the C. elegans to detect the presence of pure biofilm-secreted metabolites which had leached into the agar plate.

A Experimental design for modified choice assay. B C. elegans preferred biofilm metabolites of ΔpelA, ΔpslBCD, and ΔpelAΔpslBCD mutants over PAO1 control, CI > 0.00. C C. elegans preferred biofilm metabolites of PAO1 control over ΔwspF and PAO1/plac-YedQ mutants, CI < 0.00. D C. elegans preferred biofilm metabolites of ΔwspFΔpelA, ΔwspFΔpslBCD, and ΔwspFΔpelAΔpslBCD mutants over ΔwspF control, CI > 0.00. E C. elegans preferred biofilm metabolites of ΔpelAΔpslBCD control over ΔpelAΔpslBCD possessing exogenous Pel and Psl, CI < 0.00. Means and SD from triplicate experiments are shown. ***p < 0.001, one-way ANOVA.
In this case, C. elegans were given the choice to choose between the metabolites secreted by wild-type PAO1 and the mutant deficient in producing a biofilm matrix component, namely ΔpelA, ΔpslBCD, and ΔpelAΔpslBCD in our modified chemotaxis assay. The animals preferred the metabolites secreted by ΔpelA, ΔpslBCD, and ΔpelAΔpslBCD biofilms over those from wild-type PAO1 control (Fig. 1B), implying that both Pel and Psl were important in preventing the detection by C. elegans. We eliminated the possibility that different bacterial numbers could lead to differences in metabolite released, as CFU in biofilms remained nearly constant for PAO1 wild-type and its mutants (Supplementary Fig. S2).
We further ascertained our results by boosting biofilm formation in P. aeruginosa via the induction of c-di-GMP signaling. C-di-GMP is a bacterial secondary messenger whose synthesis by diguanylate cyclases led to motility loss and biofilm formation, while breakdown by phosphodiesterases led to biofilm dispersal [12,13,14]. We employed the ΔwspF mutant which promoted the constitutive expression of wspR DGC, resulting in increased biofilm formation via the production of Pel and Psl [15]. Mutations in the wspF gene were frequently identified in biofilm-mediated P. aeruginosa infections [16, 17]. By establishing a chemotaxis assay between wild-type PAO1 and pro-biofilm-forming ΔwspF, we showed that the animals were highly attracted toward the region previously colonized by PAO1 biofilms instead of ΔwspF biofilms, indicating the absence of a certain metabolite released by ΔwspF biofilms (Fig. 1C).
To maximize the phenotypic effects of the EPS in question and ensure that our results were solely dependent on one EPS, we mutated the EPS genes in the ΔwspF mutant. The presence of wspF mutation will boost the production of the exopolysaccharide. However, the loss of Pel or Psl in ΔwspFΔpelA, ΔwspFΔpslBCD led to chemoattraction by C. elegans when placed in a chemotaxis assay with ΔwspF (Fig. 1D). Loss of both Pel and Psl in ΔwspFΔpelAΔpslBCD exacerbated the effect, where C. elegans were highly attracted to the mutant as compared to wild-type strain (Fig. 1D).
To confirm that our observations was attributed to c-di-GMP signaling per se and not due to pleiotropic effects of the wsp operon, we first expressed a plac-YedQ plasmid containing a YedQ DGC in wild-type PAO1 to constitutively elevate intracellular c-di-GMP levels and boost Pel and Psl production [17]. We found qualitatively identical results which corroborated with our observations using ΔwspF (Fig. 1C and Supplementary Fig. S3). Although cell number in the biofilm remained consistent across the wild-type and mutants (Supplementary Fig. S2), both ΔwspF and PAO1/plac-YedQ were previously shown to produce significantly higher exopolysaccharides than PAO1 wild-type [3]. We also found no significant differences concerning other mutants of known biofilm matrix components, such as adhesion proteins and eDNA, indicating that they are not involved in the “safehouse” phenotype (Supplementary Fig. S4). Moreover, other polysaccharides, such as cellulose and pectin, could not prevent the detection of BHL by C. elegans (Supplementary Fig. S4B).
It is also crucial to find out if exogenous addition of Pel or Psl to a non-exopolysaccharide mutant (ΔpelAΔpslBCD) could prevent their colonized area from being detected by the predator. Since our previous study had shown that PAO1 wild-type biofilm contained an estimated of 1 µg ml−1 Pel and Psl in the biofilm matrix, we added 1 µg ml−1 Pel or Psl to the ΔpelAΔpslBCD and grew a biofilm overnight before removing the biofilm from the agar. Interestingly, we found that C. elegans preferentially chose the secreted metabolites of ΔpelAΔpslBCD control over ΔpelAΔpslBCD + 1 µg ml−1 Pel or Psl (Fig. 1E).
C. elegans detects BHL QS autoinducer in distance-dependent manner
We next asked what biofilm metabolite was detected by C. elegans. Since there were high levels of homoserine-based autoinducers in the biofilms, we hypothesized that the autoinducers could leach from biofilms into the environment, resulting in detection by any lurking predator. To test if C. elegans could detect the OdDHL and BHL leached from biofilms into the agar, we first placed pure OdDHL and BHL in the agar for seeping and diffusion across the agar, allowing the animals to detect the metabolites. We found that C. elegans could detect both OdDHL (C12-chain autoinducer for las QS) and BHL (C4-chain autoinducer for rhl QS) (Fig. 2A), which corroborated with a previous finding that C. elegans could detect autoinducers [9]. This could be attributed to the smaller molecular weight of BHL than OdDHL, where the concentration of BHL and OdDHL decreased over distance in the agar (Fig. 2B). We tested other known metabolites (pqs, rhamnolipid, pyoverdine and pyocyanin) and secreted protein (lasB), where they were not detected by C. elegans (Supplementary Fig. 5).

A C. elegans preferred pure autoinducers over saline control, CI > 0.00. B Pure OdDHL and BHL concentrations in the agar across distances. C BHL concentration in biofilms. D Biofilm-secreted BHL concentrations in agar across distances. E Increasing concentrations of Pel or Psl can trap 10 µM BHL and prevent predator detection. Means and SD from triplicate experiments are shown. F Cellulase degradation of exopolysaccharides results in C. elegans detection of BHL, where C. elegans preferred cellulase-degraded pure exopolysaccharide with released 10 µM BHL over pure exopolysaccharide with trapped 10 µM BHL control, CI > 0.00. *p < 0.05, ***p < 0.001, ns not significant, one-way ANOVA.
Exopolysaccharides accumulate BHL within biofilms
Since C. elegans could detect OdDHL and BHL in the agar, it is highly likely that our prior observations in Fig. 1 could be attributed to wild-type PAO1 and mutant biofilms leaching different concentrations of OdDHL and BHL. We measured the concentration of BHL within the wild-type and mutant biofilms and found no significant differences among the biofilms (Fig. 2C), indicating that there was no deficiency in BHL production in the biofilms. Interestingly, once outside the biofilm in the agar, there were higher concentrations of BHL in ΔpelA and ΔpelAΔpslBCD mutants, as compared to wild-type PAO1 and ΔpslBCD (Fig. 2D). We found qualitatively similar results for the exopolysaccharide mutants expressed with the ΔwspF background, albeit at lower levels than those under the wild-type background (Fig. 2D), indicating that overproduction of exopolysaccharides would prevent BHL from “leaking” into the environment.
To confirm our observations that Pel exopolysaccharide could trap BHL and prevent leaching into the agar, we tested if C. elegans could detect the autoinducers leached in the agar after using purified Pel and Psl to trap the autoinducers on the filter. As control, purified Pel and Psl themselves were neither attractive nor aversive cues to the animals (Fig. S6). We found that both purified exopolysaccharides at increasing concentrations could trap BHL and reduce BHL entry onto the agar, thereby preventing the detection by C. elegans (Fig. 2E). This could be attributed to the “stickiness” of the biofilm matrix which allowed it to trap small metabolites in a general manner [18]. By breaking down the exopolysaccharides in the biofilm matrix with a known anti-biofilm glycosyl hydrolase, cellulase [19], we showed that cellulase-induced degradation of biofilm matrix resulted in the detection of BHL by the animals (Fig. 2F), emphasizing the importance of biofilm matrix in “cloaking” BHL from predators.
Public good for non-producers and other microbial species
Since most microbial communities in the environment are polymicrobial in nature, some microbial species are not producers of QS or exopolysaccharides. To address if non-QS producers and other microbial species could similarly be protected by the Pel exopolysaccharide from detection by C. elegans as a form of public good, we added different concentrations of Pel or Psl and AHL or BHL exogenously in various mixtures to a non-QS producer (ΔlasIΔrhlI mutant) and another bacterial species (Escherichia coli OP50).
For non-QS producer which could not produce AHL and BHL (ΔlasIΔrhlI mutant), C. elegans could not detect the ΔlasIΔrhlI mutant in the choice assay with PAO1 control (Fig. 3A). Addition of pure AHL or BHL to ΔlasIΔrhlI mutant could restore the phenotype for C. elegans detection, but with the presence of purified Pel or Psl added to the biofilms, C. elegans could no longer detect the metabolites anymore (Fig. 3B).

A C. elegans preferentially selected PAO1 control strain as compared to ΔlasIΔrhlI mutant, CI < 0.00. B Exogenous addition of Pel or Psl protected ΔlasIΔrhlI mutant from predator detection in the presence of BHL. C Exogenous addition of Pel or Psl protected E. coli OP50 from predator detection. D Exogenous addition of Pel or Psl protected E. coli OP50 from predator detection even in the presence of BHL. Means and SD from triplicate experiments are shown. ***p < 0.001, one-way ANOVA.
For another bacterial species (E. coli OP50) which produce its own QS molecules such as AI-2 [20] (Fig. 3C). Exogenous addition of pure BHL to OP50 could restore the phenotype for C. elegans detection, but with the presence of purified Pel or Psl added to the biofilms, C. elegans could no longer detect the metabolites anymore (Fig. 3D). This indicated that Pel and Psl could be used as a public good for bacteria to hide their “signatures” from predator detection.
Loss of function of odr-10 gene in C. elegans eliminates biofilm prey detection
We next sought to examine how the autoinducer detection of biofilm prey by C. elegans are impacted when we limit the ability of C. elegans to detect autoinducers. We employed worms with a mutation in the odr-10 gene, which causes loss of odorant detection to volatile compounds [21, 22]. ODR-10 is a specific G-protein-coupled olfactory receptor that binds to diacetyl, which is a volatile odorant produced by various microbial species including lactic acid bacteria [7, 21, 22]. We showed that the exploratory behavior of odr-10 mutants was comparable to wild-type N2 animals (Supplementary Fig. S7), but the mutant worms could not detect pure autoinducers (Fig. 4A), even with Pel or Psl (Supplementary Fig. S8), and differentiate between bacterial mutants (Fig. 4B) in the choice assays.

A C. elegans odr-10 mutant could not detect pure autoinducers. B C. elegans odr-10 mutant could not differentiate mutant biofilm mutants from wild-type PAO1. C Experimental design for studying population growth of C. elegans after choice assay. D Higher population growth of wild-type N2 C. elegans which localized on metabolite spot left by ΔpelA, ΔpslBCD, and ΔpelAΔpslBCD mutants relative to odr-10 mutant. Means and SD from triplicate experiments are shown. **p < 0.01, ***p < 0.001, ns not significant, one-way ANOVA.
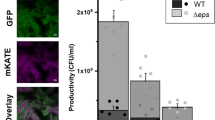
Lastly, we examined if the loss of autoinducer detection by C. elegans had a significant impact on bacterial prey survival. After performing the choice assay, we fed the worms localized at their respective corners with fresh bacteria and enumerate the bacterial populations (Fig. 4C). Since the animals preferred the metabolites secreted by ΔpelA, ΔpslBCD and ΔpelAΔpslBCD biofilms over those from wild-type PAO1 control, we observed the feeding-attributed reduction of ΔpelA, ΔpslBCD and ΔpelAΔpslBCD bacterial populations as compared to wild-type PAO1 control (Fig. 4D). This indicated that the ability to sense autoinducers could result in the predator’s preference toward its prey, which had significant impact on bacterial prey survival.
Discussion
To ensure survival from bacterivorous predation, bacteria produce different types of virulence factors of protein or toxin in nature, which can effectively eliminate their predators [23,24,25]. However, this requires hours to days in duration to kill C. elegans, as reflected by the quick paralytic killing and slow killing assays, respectively [26]. Unable to react immediately to their rapidly changing plight, biofilms need to passively protect themselves from predator, where the biofilm matrix could either slow down the motility of predators to reduce predation [3], or act as barrier to prevent direct feeding.
However, given the plethora of proteins and metabolites being secreted into the environment by bacteria, it appeared to be inevitable that biofilms will attract predators. This is especially true when there are high concentrations of QS autoinducers secreted into the environment at high cell densities or in biofilms, where autoinducers were known chemoattractants to C. elegans [9, 10]. This raised a longstanding fundamental question into how bacteria could avoid predator detection under such circumstances.
Here, we reveal a novel “stealth” approach adopted by biofilms to “hide” their biosignatures and prevent the release of QS autoinducers into the environment for predator detection (Fig. 5). This was reflected in the previously undescribed role of biofilm exopolysaccharides, in the form of Pel and Psl, to trap BHL autoinducers within the biofilms and prevent them from leaching into the surrounding agar in a distance-dependent manner, thereby preventing C. elegans detection via ODR-10 olfactory receptor. However, the exact mechanisms of how ODR-10 mediates attraction of OdDHL and BHL remain unclear. Since ODR-10 could homo-oligomerise, and even form heteromers with STR-112, another related nematode receptor [27], we speculate that such complexity could possibly help in the detection of larger chemoattractants, including OdDHL and BHL. Similar to a police and criminal-like scenario [28], the criminals (bacterial cells) could conduct their illicit meeting (QS) in the safehouse (biofilms) and prevent leaks of their communication signals (autoinducers), where the police (C. elegans) could not hijack the signals for arrest (olfactory detection and predation).

The exopolysaccharides in the biofilm matrix reduce the leaching of autoinducers into the environment. However, the absence of exopolysaccharides will enable the diffusion of autoinducers into the environment, resulting in the detection by the predator.
Our findings also had implications in infectious diseases, where nearly 80% of bacterial infections were mediated by biofilms. While it is commonly known that biofilms could evade immune detection where its biofilm matrix acts as barrier against phagocytic clearance and detection by inflammatory factors [29], the exact mechanism of preventing detection by immune cells remain poorly understood. Although our lab-based approach used to study C. elegans detection of biofilm metabolites was artificial, our findings offered insight that the biofilms reduce the likelihood of immune detection by trapping its biosignatures in its matrix exopolysaccharides. Moreover, the biofilm matrix offers a multitude of functions in infections, with recently discovered examples of offering protection of tumor cells from anticancer chemotherapy and efficient pathogen transfer across food [30, 31], thereby complicating the role of biofilms in infections. Hence, use of anti-biofilm agents to disrupt biofilms [32] could be a viable way of exposing the biofilm bacteria to immune detection and improve clearance by the host immune system.
Materials and methods
Bacterial strains and growth conditions
The bacterial strains and plasmids used in this study are listed in Supplementary Table S1. E. coli DH5a strain was used for standard DNA manipulation, while E. coli OP50 was used for growth and maintenance of C. elegans. Luria-Bertani (LB) medium was used to cultivate E. coli and P. aeruginosa strains. For plasmid maintenance in E. coli, the medium was supplemented with 100 μg ml−1 ampicillin and 15 μg ml−1 gentamicin. For marker selection in P. aeruginosa, 30 μg ml−1 gentamicin, 30 μg ml−1 tetracycline or 200 μg ml−1 carbenicillin were used, as appropriate.
Growth and maintenance of C. elegans
As previously described [33], the bacterial lawn, such as E. coli OP50 and P. aeruginosa strains, were cultivated on NGM agar plates at 37 °C for 16 h. The laboratory C. elegans N2 nematode was transferred to the bacterial lawn on the NGM agar plates and cultivated at room temperature for 72 h for population expansion. The CX32 strain possessing the odr-10 mutation was procured from the Caenorhabditis Genetics Center for experiments.
C. elegans choice assay of biofilm
P. aeruginosa wild-type PAO1 and its respective mutants were first inoculated and grew in 2 ml of LB medium at 37 °C with shaking at 200 rpm for 16 h. After washing the overnight culture with 0.9% NaCl and adjustment of OD600 to 0.1, the 30 µl of bacterial cultures (control strain and test strain) were transferred directly to opposite ends of NGM petri dish. The cultures were spread carefully on the agar with a bacteriological spreader. The culture was incubated at 37 °C for 16 h to allow biofilm growth on the agar. At least 50 individuals of adult C. elegans from 3 independent trials were transferred from the feeding plate to the center of the petri dish with wire picker. Live animals localized at each biofilm spot were enumerated by 6 h.
C. elegans modified choice assay of biofilm metabolites
P. aeruginosa wild-type PAO1 and its respective mutants were first inoculated and grew in 2 ml of LB medium at 37 °C with shaking at 200 rpm for 16 h. After washing the overnight culture with 0.9% NaCl and adjustment of OD600 to 0.1, the 30 µl of bacterial cultures (control strain and test strain) were transferred to 0.2-µm filters (Nanopore) which were placed on opposite ends of NGM petri dish. The cultures were spread carefully on the filter with a bacteriological spreader, where the cultures should not be directly in touch with the NGM agar. The culture was incubated at 37 °C for 16 h to allow biofilm growth and diffusion of secreted metabolites through the filter into the NGM agar.
For chemoattractant test, the pure chemical (OdDHL, BHL, PQS, rhamnolipid, pyocyanin and pyoverdine) at 10 µM and purified polymers (pectin, cellulose and P. aeruginosa genomic DNA) at 10 µg ml−1 was placed on the 0.2-µm filter one end of NGM plate, while its saline control (0.9% NaCl) was placed on the opposite end of the petri dish, followed by 16 h incubation at 37 °C.
The filters (containing biofilms or chemicals) on opposite ends of petri dish were carefully peeled off from the NGM agar, leaving behind the secreted biofilm metabolites in the agar. At least 50 individuals of adult C. elegans from three independent trials were transferred from the feeding plate to the center of the petri dish with wire picker. The animals were allowed to detect the secreted biofilm metabolites and migrate to the spot previously occupied by either control strain or test strain, where live animals localized at each spot were enumerated by 6 h.
Tabulation of choice index (CI)
The CI was tabulated in Eq. (1):
CI > 0.00 indicated animal preference for test strain or compounds, where CI < 0.00 indicated animal preference for control strain or compounds. CI thresholds for non-preference were set as a range of −0.1 < CI < 0.1.
Quantification of secreted autoinducers in biofilms and on agar
As previously described [13], the biofilms grown on filter on agar were scrapped and transferred to 1 ml saline (0.9% NaCl). The biofilms were then homogenized to single cells using water sonication in ice bath for 15 min and 15-s vortexing, to release the secreted autoinducers into the solution. The cells were removed via centrifugation at 13,000 × g for 3 min, and the supernatants containing autoinducers were filtered through 0.2-μm filters.
As quantifying autoinducers in the agar, the agar was cut at different distances (0, 1.5, and 3 cm) from the center of biofilm on filter, and transferred to 1 ml saline (0.9% NaCl). The agar was mashed into smaller pieces using a microtube pestle and placed at 4 °C overnight for metabolites to diffuse into the saline. The agar pieces were removed via centrifugation at 13,000 × g for 3 min, and the supernatants containing autoinducers were filtered through 0.2-μm filters.
Overnight culture of the reporter strain ΔlasIΔrhlI/plasB-gfp or ΔlasIΔrhlI/prhlA-gfp for quantifying OdDHL and BHL respectively, was adjusted to OD600 = 0.1 using 2-fold ABTGC medium. In total, 100 μl of filtrate was added to 100 μl of reporter strain in a 96-well plate (SPL, South Korea). As ΔlasIΔrhlI does not produce OdDHL and BHL, plasB-gfp or prhlA-gfp was induced by the addition of biofilm filtrates containing OdDHL or BHL respectively. GFP fluorescence from prhlA-gfp expression (expressed in relative fluorescence units, RFU) was measured for each well using a microplate reader (Tecan Infinite 2000) and was normalized to the OD600 of each well. To quantify autoinducer concentration, the standard curves for OdDHL and BHL (using 0, 0.1, 1, 5, 10, 20, and 30 µM) was generated using the similar assay for comparison to test samples. Experiments were performed in triplicate, and results are shown as the mean ± SD.
Colony-forming unit (CFU) assay
As previously described [34], the biofilms were quantified using the CFU assay. The biofilms grown on filter on agar were scrapped and transferred to 1 ml saline (0.9% NaCl). The biofilms were then homogenized to single cells using water sonication in ice bath for 15 min and 15-s vortexing. Bacterial cells were serially diluted, plated on LB agar and incubated at 37 °C for 16 h. CFU ml−1 was calculated as average number of colonies by the dilution factor and dividing by the volume. Experiments were performed with three replicates, and the results are shown as the mean ±SD.
Extraction of polysaccharides
As previously described [3], Pel and Psl were extracted by growing ΔpslBCD/plac-YedQ, and ΔpelA/plac-YedQ static biofilms in 15 ml ABTGC in standard Petri dishes at 37 °C for 16 h. The biofilms were collected and separated from the supernatant by centrifugation at 10,000 × g for 5 min. The cell pellet was re-suspended in 0.9% NaCl and homogenized with mild water sonication on ice (Elmasonic P120H, Power = 50%, Frequency = 37 KHz) for 5 min to dislodge the surface-associated matrix from bacterial cells. The cells were then separated from the matrix by centrifugation at 13,000 × g for 3 min, leaving behind the crude matrix extract.
The crude extract was then further treated by removal of eDNA by precipitation with 25% ethanol and 0.1 M CaCl2. Extracellular proteins were then removed from the extract with 0.5 mg ml−1 proteinase K at 60 °C for 1 h and inactivation at 80 °C for 30 min. The extract was then filtered with centrifugal filter (<3 kDa) at 15,000 × g for 5 min to remove the bacterial metabolites. The extract was then lyophilized and re-suspended in 1 ml sterile ddH2O.
C. elegans choice assay of purified exopolysaccharides and autoinducers
Varying concentrations (0, 0.5, 1, 5, and 10 µg ml−1) of Pel or Psl were mixed with 10 µM OdDHL or BHL, and the solutions were added to the 0.2-µm filter at one end of NGM plate, while its control was placed on the opposite end of the petri dish, followed by 16 h incubation at 37 °C. This allowed the OdDHL or BHL to diffuse through the filter, while Pel or Psl were retained on the filter. The filters (containing biofilms or chemicals) on opposite ends of petri dish were carefully peeled off from the NGM agar, leaving behind the diffused autoinducers in the agar. At least 50 individuals of adult C. elegans from three independent trials were transferred from the feeding plate to the center of the petri dish with wire picker. The animals were allowed to detect the secreted biofilm metabolites and migrate to the spot previously occupied by either control strain or test strain, where live animals localized at each spot were enumerated by 6 h.
A modified choice assay involving cellulase treatment to degrade the exopolysaccharides was also used, where Pel or Psl were pre-treated with 5 µg ml−1 of cellulase for 24 h at 37 °C. The cellulase was heat-inactivated for 1 h at 65 °C, where remaining Pel and Psl were purified as described in previous section, before use for choice assay.
Exogenous addition of exopolysaccharides and autoinducers to non-producing strains
The P. aeruginosa ΔpelAΔpslBCD or E. coli OP50 cells from growth culture (described in the previous section on growth conditions) were washed with 0.9% NaCl and centrifuged at 13,000 × g for 3 min. Varying concentrations (0, 0.5, 1, 5, and 10 µg ml−1) of Pel or Psl were mixed with 10 µM OdDHL or BHL, and the solutions were re-suspended in the cell pellet. The various combinations were used for the modified choice assays.
C. elegans exploration assay
The exploratory behavior of wild-type (N2) nematodes and odr-10 (CX32) mutant was compared, where animals were transferred to NGM plates containing OP50 bacterial lawn. The trails of nematodes were monitored every 12 h till 48 h and recorded by enumerating the grids (5 mm × 5 mm size) covered by the trails.
Bacterial population survival after C. elegans feeding
To observe if nematodes attracted to the biofilm metabolite spot could reproduce and expand their population after the choice assay, the worms localized at their respective spots were transferred to fresh NGM plates containing OP50 lawn. The fresh NGM plates were further incubated at room temperature for 48 h to allow C. elegans feeding of bacterial prey. Bacteria on the lawns after 48 h of C. elegans L1 progeny growth were harvested for CFU assay, as described in earlier section.
Statistical analysis
The results were expressed as means ± standard deviation. Data groups were compared using the one-way ANOVA and Student’s t test to evaluate associations between independent variables, and the p values were obtained. Three independent trials were conducted in triplicates for each experiment.
References
Jessup CM, Forde SE, Bohannan BJM. Microbial experimental systems in ecology. In: Desharnais RA, editor. Advances in ecological research, Vol. 37. Elsevier, USA: Academic Press; 2005. p. 273–307.
Brockmann D, Hufnagel L, Geisel T. The scaling laws of human travel. Nature. 2006;439:462–5.
Chan SY, Liu SY, Seng Z, Chua SL. Biofilm matrix disrupts nematode motility and predatory behavior. ISME J. 2021;15:260–9.
Thutupalli S, Uppaluri S, Constable GWA, Levin SA, Stone HA, Tarnita CE, et al. Farming and public goods production in Caenorhabditis elegans populations. Proc Natl Acad Sci USA. 2017;114:2289–94.
Otto G. Arresting predators. Nat Rev Microbiol. 2020;18:675.
Worthy SE, Haynes L, Chambers M, Bethune D, Kan E, Chung K, et al. Identification of attractive odorants released by preferred bacterial food found in the natural habitats of C. elegans. PLoS ONE. 2018;13:e0201158.
Choi JI, Yoon K-H, Subbammal Kalichamy S, Yoon S-S, Il Lee J. A natural odor attraction between lactic acid bacteria and the nematode Caenorhabditis elegans. ISME J. 2016;10:558–67.
Reilly DK, Srinivasan J. Caenorhabditis elegans olfaction. Oxford Research Encyclopedia of Neuroscience: Oxford University Press; 2017.
Beale E, Li G, Tan M-W, Rumbaugh KP. Caenorhabditis elegans senses bacterial autoinducers. Appl Environ Microbiol. 2006;72:5135–7.
Werner KM, Perez LJ, Ghosh R, Semmelhack MF, Bassler BL. Caenorhabditis elegans recognizes a bacterial quorum-sensing signal molecule through the AWCON neuron. J Biol Chem. 2014;289:26566–73.
Wei Q, Ma LZ. Biofilm matrix and its regulation in Pseudomonas aeruginosa. Int J Mol Sci. 2013;14:20983–1005.
Tal R, Wong HC, Calhoon R, Gelfand D, Fear AL, Volman G, et al. Three cdg operons control cellular turnover of cyclic di-GMP in Acetobacter xylinum: genetic organization and occurrence of conserved domains in isoenzymes. J Bacteriol. 1998;180:4416–25.
Chua SL, Liu Y, Li Y, Jun Ting H, Kohli GS, Cai Z, et al. Reduced Intracellular c-di-GMP content increases expression of quorum sensing-regulated genes in Pseudomonas aeruginosa. Front. Cell. Infect. Microbiol. 2017;7:451.
Hengge R. Principles of c-di-GMP signalling in bacteria. Nat Rev Microbiol. 2009;7:263–73.
Hickman JW, Tifrea DF, Harwood CS. A chemosensory system that regulates biofilm formation through modulation of cyclic diguanylate levels. Proc Natl Acad Sci USA. 2005;102:14422–7.
Smith EE, Buckley DG, Wu Z, Saenphimmachak C, Hoffman LR, D’Argenio DA, et al. Genetic adaptation by Pseudomonas aeruginosa to the airways of cystic fibrosis patients. Proc Natl Acad Sci USA. 2006;103:8487–92.
Chua SL, Ding Y, Liu Y, Cai Z, Zhou J, Swarup S, et al. Reactive oxygen species drive evolution of pro-biofilm variants in pathogens by modulating cyclic-di-GMP levels. Open Biol. 2016;6:160162.
Seviour T, Hansen SH, Yang L, Yau YH, Wang VB, Stenvang MR, et al. Functional amyloids keep quorum-sensing molecules in check. J Biol Chem. 2015;290:6457–69.
Ma L, Conover M, Lu H, Parsek MR, Bayles K, Wozniak DJ. Assembly and development of the Pseudomonas aeruginosa biofilm matrix. PLoS Pathog. 2009;5:e1000354.
Whitehead NA, Barnard AML, Slater H, Simpson NJL, Salmond GPC. Quorum-sensing in Gram-negative bacteria. FEMS Microbiol Rev. 2001;25:365–404.
Zhang Y, Chou JH, Bradley J, Bargmann CI, Zinn K. The Caenorhabditis elegans seven-transmembrane protein ODR-10 functions as an odorant receptor in mammalian cells. Proc Natl Acad Sci USA. 1997;94:12162–7.
Sengupta P, Chou JH, Bargmann CI. odr-10 encodes a seven transmembrane domain olfactory receptor required for responses to the odorant diacetyl. Cell. 1996;84:899–909.
Cezairliyan B, Vinayavekhin N, Grenfell-Lee D, Yuen GJ, Saghatelian A, Ausubel FM. Identification of Pseudomonas aeruginosa phenazines that kill Caenorhabditis elegans. PLoS Pathog. 2013;9:e1003101.
Gallagher LA, Manoil C. Pseudomonas aeruginosa PAO1 kills Caenorhabditis elegans by cyanide poisoning. J Bacteriol. 2001;183:6207–14.
Lewenza S, Charron-Mazenod L, Giroux L, Zamponi AD. Feeding behaviour of Caenorhabditis elegans is an indicator of Pseudomonas aeruginosa PAO1 virulence. PeerJ. 2014;2:e521–e.
Tan MW, Mahajan-Miklos S, Ausubel FM. Killing of Caenorhabditis elegans by Pseudomonas aeruginosa used to model mammalian bacterial pathogenesis. Proc Natl Acad Sci USA. 1999;96:715–20.
Tehseen M, Liao C, Dacres H, Dumancic M, Trowell S, Anderson A. Oligomerisation of C. elegans olfactory receptors, ODR-10 and STR-112, in yeast. PLoS ONE. 2014;9:e108680.
Sooknanan J, Bhatt B, Comissiong DMG. A modified predator-prey model for the interaction of police and gangs. R Soc Open Sci. 2016;3:160083.
Arciola CR, Campoccia D, Montanaro L. Implant infections: adhesion, biofilm formation and immune evasion. Nat Rev Microbiol. 2018;16:397–409.
Deng Y, Liu SY, Chua SL, Khoo BL. The effects of biofilms on tumor progression in a 3D cancer-biofilm microfluidic model. Biosens Bioelectron. 2021;180:113113.
Kwok T-Y, Ma Y, Chua SL. Biofilm dispersal induced by mechanical cutting leads to heightened foodborne pathogen dissemination. Food Microbiol. 2022;102:103914.
Yu M, Chua SL. Demolishing the great wall of biofilms in gram-negative bacteria: to disrupt or disperse? Medicinal Res Rev. 2020;40:1103–16.
Chua SL, Liu Y, Yam JKH, Chen Y, Vejborg RM, Tan BGC, et al. Dispersed cells represent a distinct stage in the transition from bacterial biofilm to planktonic lifestyles. Nat Commun. 2014;5:4462.
Liu SY, Leung MM-L, Fang JK-H, Chua SL. Engineering a microbial ‘trap and release’ mechanism for microplastics removal. Chem Eng J. 2021;404:127079.
Acknowledgements
This research is supported by The Hong Kong Polytechnic University, Department of Applied Biology and Chemical Technology Startup Grant (BE2B), Departmental General Research Fund (UALB), One-line account (ZVVV), Environmental and Conservation Fund (ECF-48/2019), Health and Medical Research Fund (HMRF-201903032), Pneumoconiosis Compensation Fund Board (PCFB-ZJN2), and State Key Laboratory of Chemical Biology and Drug Discovery Fund (1-BBX8). Some C. elegans strains were provided by the Carnorhabditis Genetics Center (CGC), University of Minnesota, which is funded by NIH Office of Research Infrastructure Programs (P40 OD010440).
Author information
Authors and Affiliations
Contributions
SLC designed methods and experiments. SYC, SYL, and SL carried out laboratory experiments, analyzed the data and interpreted the results. SLC, SL, SYL, and SYC wrote the paper. All authors have contributed to, seen and approved the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
About this article

Cite this article
Li, S., Liu, S.Y., Chan, S.Y. et al. Biofilm matrix cloaks bacterial quorum sensing chemoattractants from predator detection. ISME J 16, 1388–1396 (2022). https://doi.org/10.1038/s41396-022-01190-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41396-022-01190-2
This article is cited by
-
Distinct bacterial population dynamics and disease dissemination after biofilm dispersal and disassembly
The ISME Journal (2023)
-
Biofilm control strategies in the light of biofilm-forming microorganisms
World Journal of Microbiology and Biotechnology (2023)
