
Abstract
Alcohol use disorders are chronic debilitating diseases characterized by severe withdrawal symptoms that contribute to morbidity and relapse. GABAA receptor (GABAAR) adaptations have long been implicated in the chronic effects of alcohol and contribute to many withdrawal symptoms associated with alcohol dependence. In rodents, GABAAR hypofunction results from decreases in Gabra1 expression, although the underlying mechanism controlling Gabra1 expression after chronic ethanol exposure is still unknown. We found that chronic ethanol exposure using either ethanol gavage or two-bottle choice voluntary access paradigms decreased Gabra1 expression and increased Hdac2 and Hdac3 expression. Administration of the HDAC inhibitor trichostatin A (TSA) after chronic ethanol exposure prevents the decrease in Gabra1 expression and function as well as the increase in Hdac2 and Hdac3 expression in both the cortex and the medial prefrontal cortex (mPFC). Chronic ethanol exposure and withdrawal, but not acute ethanol exposure or acute withdrawal, cause a selective upregulation of HDAC2 and HDAC3 associated with the Gabra1 promoter that accompanies a decrease in H3 acetylation of the Gabra1 promoter and the reduction in GABAAR α1 subunit expression. TSA administration prevented each of these molecular events as well as behavioral manifestations of ethanol dependence, including tolerance to zolpidem-induced loss of righting reflex, reduced open-arm time in the elevated plus maze, reduced center-time and locomotor activity in the open-field assay, and TSA reduced voluntary ethanol consumption. The results show how chronic ethanol exposure regulates the highly prominent GABAAR α1 subunit by an epigenetic mechanism that represents a potential treatment modality for alcohol dependence.
Similar content being viewed by others

Introduction
Alcohol use disorders are chronic debilitating disorders and one of the leading causes of preventable death in the United States [1] and are characterized by severe withdrawal symptoms. Withdrawal symptoms include central nervous system hyperexcitability, tremor, anxiety, insomnia, seizures, cross-tolerance to benzodiazepines, and the severity of these symptoms is correlated with likelihood to relapse [2]. Alcohol withdrawal symptoms are known to be linked to deficits in GABAergic signaling in multiple brain regions and can be readily treated acutely with GABAergic modulators, such as benzodiazepines [3]. However, benzodiazepines have severe disadvantages for long-term treatment of alcohol use disorders, due to their high abuse potential, cross-tolerance with alcohol, high risk of mortality when taken in combination with alcohol, and, with prolonged use, decreases in GABAAR expression [3,4,5]. In rodent models, many symptoms of dependence, withdrawal, and GABAergic hypofunction are correlated with downregulation of the α1 subunit and upregulation of the α4 subunit [4, 6,7,8,9,10,11,12]. GABAARs containing the α1 subunit are the most prominent receptor in many brain regions including the cortex [13] and mediate the sedative effects of GABAAR agonists and positive allosteric modulators [9]. Knockout of the α1 subunit also causes an essential tremor similar to the phenotype observed in human patients with severe alcohol dependence [8]. Meanwhile, upregulation of the α4 subunit likely contributes to benzodiazepine cross-tolerance and increased anxiety [14]. Changes in α1 and α4 expression are known to cause changes in GABAAR function in vitro by modifying miniature inhibitory post-synaptic current characteristics, specifically decreasing decay τ1, and these changes may have relevance in vivo [15, 16]. It has been known for many years that decreases in α1 expression and increases in α4 expression occur on a transcriptional level [7], but few studies have explored how either transcript is regulated epigenetically by ethanol.
Epigenetic mechanisms are broadly defined as changes to gene expression that are not due to mutations in the gene. Changes in chromatin structure through post-translational modifications of histone tails causes both short- and long-lasting changes in gene expression in response to environmental stimulus or drug exposure. The most well-known of these post-translational modifications is acetylation of histone tails that serves to open the chromatin to allow increased transcription [17]. Acetylation/deacetylation of histones is facilitated by a large family of histone acetyltransferases and histone deacetylases (HDACs), respectively. Several studies have shown that drugs of abuse modulate gene transcription and expression through epigenetic mechanisms, including acetylation [18,19,20], and that epigenetic intervention using HDAC inhibitors can prevent molecular and behavioral changes induced by repeated exposure to drugs of abuse. Therefore, it could be possible that decreases in Gabra1 expression after chronic ethanol exposure may be prevented by epigenetic intervention. Recent studies in our laboratory screened a number of HDAC inhibitors in vitro to demonstrate that ethanol-induced changes in GABAAR gene expression can be prevented by the co-exposure with various HDAC inhibitors, including trichostatin A (TSA) [21]. TSA was selected for the current study because it was a very potent inhibitor of HDAC2 and HDAC3 in our screen of HDAC inhibitors that prevent ethanol-induced changes in GABAARs [21], and also because HDAC inhibitors only effect 2% of genes, indicating their relative selectivity [22]. Since GABAAR hypofunction occurs after chronic ethanol exposure in vivo, we evaluated the efficacy of the most potent HDAC inhibitor in our in vitro screen, TSA, to prevent changes in GABAAR expression, function, and evaluate behavioral correlates in rodent models of dependence.
In order to determine if an epigenetic mechanism regulates the effects of ethanol in vivo, we investigated the efficacy of an HDAC inhibitor preventing diminished Gabra1 or α1 subunit expression in rat cerebral cortex after chronic ethanol exposure using two different models of ethanol dependence. We focused on cerebral cortex because decreases in benzodiazepine binding occur in human cortex consistent with GABAergic dysfunction [23]. We next investigated whether regions of the cortex involved in the neurocircuitry of addiction, the prefrontal cortex (PFC), also exhibited changes in GABAAR expression after chronic ethanol, and if these changes were due to epigenetic mechanisms that could be prevented by the administration of TSA using both voluntary and involuntary alcohol exposures [24, 25]. Our results indicate that ethanol alters GABAAR function and expression via epigenetic mechanisms involving deacetylation of histones associated with the Gabra1 promoter by HDAC2 and HDAC3. Preventing these changes using an HDAC inhibitor prevents molecular, epigenetic, and functional adaptations as well as alcohol dependence-related behaviors.
Materials and Methods
Animals and treatments
Adult male Sprague Dawley rats (200–300 g, 8–9 weeks old, Envigo) were used for ethanol administration by gavage. Adult male Wistar rats were used for voluntary ethanol self-administration by two-bottle choice (200–300 g, 8–9 weeks old, Charles River Laboratories, Inc.). All procedures were carried out in compliance with guidelines specified by the Institutional Animal Care and Use Committee at the University of North Carolina at Chapel Hill.
Intragastric gavage ethanol-exposure paradigm
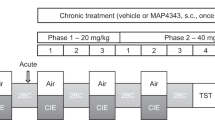
Sprague Dawley rats were housed in pairs in a temperature (23 °C) and humidity controlled vivarium under a 12 h light/dark cycle (lights on from 0700 to 1900 hours) and given ad libitum access to food and water. For chronic ethanol administration and withdrawal experiments, rats were given a single water or ethanol dose (5 g/kg, 25% v/v, i.g.) once daily for 14 days, then allowed to withdraw for 24 h. On the last 2 days of ethanol exposure and the day of withdrawal, rats were injected with either TSA (2 mg/kg, i.p.) or vehicle (10% DMSO in 0.9% saline) 2 h before gavage or sacrifice. TSA exposure time and dose were chosen based on previous studies [26, 27]. At the end of all experiments, rats were killed by decapitation and cortices were dissected and snap frozen using dry ice.
Intermittent ethanol access using two-bottle choice
Intermittent two-bottle choice (I2BC) drinking was performed as previously described [28]. Adult male Wistar rats were singly housed in a reverse dark cycle room (lights off from 1000 to 2200 hours) and were given access to 20% ethanol (EtOH) in tap water, or tap water alone during the dark cycle on Monday, Wednesday, and Friday. Volumes of ethanol and tap water were measured on Tuesday, Thursday, and Saturday at the end of the light cycle. Rodents had ad libitum access to food and water. After session 8, all rats had established a stable drinking baseline that was similar to previously reported values [28]. On session 14, 2 h before two-bottle choice, rats were given an injection of vehicle (10% DMSO in 0.9% saline, i.p.) or TSA (2 mg/kg, i.p.). After I2BC, rats were sacrificed with decapitation, and PFC was dissected.
RNA isolation and qPCR
Cortices were rinsed with ice-cold PBS then homogenized in Trizol (Ambion) and RNA was extracted and purified according to manufacturer’s instructions. RNA was quantified and quality controlled using a Nanodrop spectrophotometer (all 260/280 and 230/260 values ≥1.8, Fisher Scientific). Purified RNA (2 µg) was reverse transcribed to a cDNA library using high capacity RNA to DNA kit (Applied Biosystems) following manufacturer’s instructions. DNA (10 ng per reaction) was then subjected to quantitative PCR (qPCR) analysis using TaqMan gene expression probes and Taq Gene Expression MasterMix (Supplementary Table 2, Applied Biosystems). Reactions were run in duplicate on a StepOnePlus RT-PCR system (Applied Biosystems) using glyceraldehdyde-3-phosphate dehydrogenase (Gapdh) as a loading control [29]. Data was analyzed using the ΔΔCT method and expressed as fold control.
Western blot analysis
Western blots were performed as previously described, with some modifications [29]. Cortices were rinsed with ice-cold PBS, then subjected to subcellular fractionation using the Subcellular Protein Fractionation Kit for Tissues following manufacturer’s instructions (ThermoScientific). Primary antibodies (Supplementary Table 1) were added to the blot and incubated overnight at 4 °C. Bands were visualized using the Odyssey Classic Imaging System (LI-COR) and band density was determined using ImageStudio Lite (LI-COR). Results were normalized to β-actin, GAPDH, or histone 3 depending on the cellular fraction and expressed as percent control values.
Locomotor activity assay
General locomotor activity was assessed following 14 days of ethanol administration (light cycle, 1400 hours), 24 h into withdrawal, and 2 h after TSA/vehicle injections by placing rats in Plexiglas open-field chambers (43.2 × 43.2 cm; Med Associates). Horizontal distance traveled (cm) was determined from the number of photobeam breaks in 2 min time intervals using Activity Monitor software (Med Associates). Time spent in the center (21.6 × 21.6 cm) of the open field was expressed as the % of total locomotor activity. This ratio was chosen to account for differences in total locomotor activity between treatment groups. Groups were counterbalanced to different chambers.
Zolpidem-induced loss of righting reflex
Zolpidem-induced hypnosis was assessed 24 h after the final ethanol gavage on day 14 and 2 h after TSA injections (light cycle, 1400 hours). Rats were administered zolpidem (60 mg/kg i.p., dissolved in 45% (w/v) β-cyclodextrin in 0.9% saline, Tocris) and placed in the supine position in custom built trellises. Duration of loss of righting reflex (LORR) was defined as the time required to regain the ability to right three times in 1 min.
Elevated plus maze
Elevated plus maze (EPM) was performed on the 18th session of I2BC as previously described [30]. After the beginning of the dark cycle, Wistar rats were brought into the testing room and allowed to acclimate to room environment for 30 min. Rats were then placed in EPM apparatus (Stoelting Co.) and allowed to freely explore for 5 min. Locomotor activity was recorded using an overhead camera and data was processed using ANY-Maze™ (Stoelting Co., v. 4.82). Analysis of behavior was performed as % time spent in open arms over total time.
Chromatin immunoprecipitation
Chromatin immunoprecipitations (ChIP) were performed as previously described [21]. Cortices were rinsed with ice-cold PBS, homogenized, then cross-linked in 1% formaldehyde (ThermoScientific) for 10 min at room temperature, and quenched with 125 mM glycine for 5 min at room temperature. Samples were lysed using ChIP lysis buffer (10 mM Tris-HCl, ph 8.0, 10 mM NaCl, 0.2% v/v NP-40, and 1 mM phenylmethylsulfonyl fluoride) on ice for 30 min. Nuclei were lysed in nuclear lysis buffer (10 mM Tris-HCl, ph 8.0, 50 mM EDTA, 1% v/v sodium dodecyl sulfate (SDS), and protease inhibitors) on ice for 10 min, then sonicated to shear chromatin. Following sonication, chromatin samples were clarified using centrifugation and DNA concentrations were determined using a Nanodrop (Fisher Scientific). Equal amounts of chromatin were incubated overnight at 4 °C with antibody (Supplementary Table 1), and an aliquot was set aside to determine input and to ensure equal loading. PureProteome™ Protein G Magnetic beads (Millipore) were added to chromatin samples for 1 h at 4 °C, then complexes were washed, eluted in 1% (w/v) SDS, 0.75% (w/v) sodium bicarbonate buffer, and crosslinks were reversed overnight at 65 °C. DNA was purified using QIAquick PCR Purification Kit (Qiagen) and analyzed using SYBR® Green Real-Time PCR Master Mixes (ThermoScientific) following manufacturer’s instructions. Primers for qPCR analysis are included in Supplementary Table 2.
Medial prefrontal cortex slice electrophysiology
Adult male Sprague Dawley rats were killed via rapid decapitation and their brains extracted 24 h after the last water or ethanol treatment and ~300 µm thick coronal brain slices containing the mPFC were prepared. Individual slices were placed into the recording chamber and continuously perfused with artificial cerebrospinal fluid (aCSF) composed of the following in mM: [NaCl (125), KCl (2.5), NaH2PO4 (1.5), MgCl2 (1.3), CaCl2 (2.6), Ascorbic Acid (0.4) Glucose (10), NaHCO3 (25), osmolarity adjusted to ~310 mOsm with sucrose] supplemented with DL-2-Amino-5-Phosphonovaleric acid (50 µM), 6-cyano-7-nitroquinoxaline-2,3-dione (10 µM), and 3-[[[(3,4-Dichlorophenyl)methyl]amino]propyl](diethoxymethyl) phosphinic acid (10 µM) (Abcam) at a flow rate of 2 ml/min. Electrodes containing an internal solution (in mM): CsCl (140), MgCl2 (2), EGTA (1), HEPES (10), Na-ATP (2), Na-GTP (0.3), phosphocreatine (5), QX-314 (3), pH adjusted to 7.2 with CsOH and osmolarity adjusted to ~290 with sucrose were used to patch deep-layer pyramidal neurons after obtaining a >1 GΩ seal. All cells were held at a −70 mV holding potential for the duration of recording. Pharmacologically isolated GABAAR-mediated currents were acquired using an Axon Instruments MultiClamp 700A amplifier (Molecular Devices) and digitized by an Axon Instruments Digidata 1440A digitizer (Molecular Devices) controlled by pClamp10 software (Molecular Devices). Recordings were sampled at 10 kHz and filtered at 2 kHz. Access resistance was monitored throughout the duration of the recording and the recording was terminated if access resistance exceeded 25 MΩ or changed by >30%. For kinetic analyses, a minimum of 25 visually inspected spontaneous inhibitory post-synaptic currents (sIPSCs) exhibiting a stable baseline, sharp falling phase, and single peak were selected for analysis using MiniAnalysis software (v6.0.7, Synaptosoft). Captured events were then used to construct a scaled average composite trace to which a double exponential curve was fit to determine decay time constants.
Statistics
All groups were randomly assigned. Two-way analysis of variances (ANOVAs) were performed to determine significance for all experiments that contained more than three groups. One-way ANOVAs were performed to determine significance for experiments containing three groups. Bonferroni’s post hoc test was used to perform multiple comparisons between groups in order to determine significance after either two-way or one-way ANOVAs. Student’s unpaired two-tailed t test was used to compare two groups. Values were excluded from analysis (decided a priori) if they were two standard deviations from the mean. Significance was set at p < 0.05.
Results
TSA prevents changes in GABAARs and global histone acetylation caused by chronic ethanol exposure and withdrawal
Chronic ethanol exposure by inhalation, liquid diet, or gavage causes a decrease in GABAAR α1 expression and increase in α4 expression in the cerebral cortex and hippocampus that are associated with behavioral and pharmacological maladaptations found in alcohol dependence in both Sprague Dawley and Wistar rats [4, 6, 7, 10, 31]. We previously found that TSA co-exposure in vitro would prevent changes in GABAAR subunit expression in cultured cortical neurons following ethanol exposure [21]. Here, we investigated whether TSA administration would prevent decreases in GABAAR expression in the cerebral cortex after chronic ethanol administration for 14 days and 24 h of withdrawal. The HDAC inhibitor TSA was administered for the last 2 days of ethanol exposure and the day of withdrawal (Fig. 1a). In agreement with previous studies using a liquid diet drinking paradigm [6, 7], 14-day chronic ethanol (5 g/kg, i.g. daily) administration caused a decrease in both Gabra1 transcript (Fig. 1b, 0.35 ± 0.07 fold control) and α1 subunit membrane expression (Fig. 1c, d, 67.37 ± 2.46% control). Further, TSA (2 mg/kg, i.p.) prevented the effect of ethanol (Fig. 1b, Gabra1: 0.99 ± 0.10 fold control, p < 0.0001; Fig. 1c, d; α1: 88.83 ± 2.30% control, p < 0.05). We also observed an increase in Gabra4 transcript (Fig. 1b, 2.14 ± 0.13 fold control) and α4 subunit membrane expression (Fig. 1c, d, 129.70 ± 3.45% control), which was also prevented by the administration of TSA (Fig. 1b, Gabra4: 0.93 ± 0.08 fold control, p < 0.0001; Fig. 1c, d, α4: 96.48 ± 7.44% control, p < 0.01). We did not observe any changes in other GABAAR subunit transcripts or subunits after ethanol administration, nor did TSA have any direct effect on GABAAR subunit transcript or membrane expression (Fig. 1b and Fig. 1c, d, respectively). Since TSA is an HDAC inhibitor, we next assessed whether ethanol altered histone acetylation after chronic exposure (Fig. 1e, f). Western blot analysis using a pan-histone acetylation antibody revealed a decrease in H3 acetylation (H3Ac) after chronic ethanol exposure (Fig. 1f, 83.04 ± 5.76% control) that was prevented by TSA (223.80 ± 12.55% control, p < 0.0001), while TSA injection alone increased H3Ac (347.80 ± 18.37% control, p < 0.0001). No treatment altered total H3 protein levels (Fig. 1e). We next assessed H3Ac marks associated with active promoters [17], H3Ac at lysine 9 (H3K9Ac), and H3Ac at lysine 14 (H3K14Ac). Chronic ethanol administration caused a decrease in both H3K9Ac (16.84 ± 11.49% control, p < 0.01) and H3K14Ac (12.07 ± 5.13% control), which was prevented by TSA (H3K9Ac, 103.60 ± 19.85% control, p < 0.01; H3K14Ac 92.31% ± 22.32% control, p < 0.001). TSA alone increased H3K14Ac and H3pAc, but TSA did not increase H3K9Ac (Fig. 1e, f). We next evaluated methylation marks, as histone post-translational marks are known to be co-regulated [17]. Interestingly, we found no change in permissive histone methylation marks lysine 4 dimethylation (H3K4me2), histone lysine 4 trimethylation (H3K4me3), or in the repressive mark histone lysine 9 trimethylation (Fig. 1e, f, H3K9me3).

Chronic ethanol exposure-induced changes in GABAAR expression and H3 acetylation in cerebral cortex are prevented by TSA. a Schematic outlining 14-day gavage procedure (5 g/kg, 25% EtOH) with 24 h withdrawal. TSA (2 mg/kg, i.p.) was administered on the last 3 days of the procedure. b qPCR analysis reveals that chronic ethanol exposure causes decreases in Gabra1 and increases in Gabra4 mRNA transcripts that are prevented by the administration of TSA. A significant interaction between ethanol and TSA exposure was observed for Gabra1 (F1,20 = 19.42, p = 0.0003, n = 6) and Gabra4 (F1,20 = 30.11, p < 0.0001, n = 6), using two-way ANOVA. Values are normalized to Gapdh, and expressed as fold control. c Representative western blots of GABAA subunit membrane expression following chronic 14-day ethanol administration with 24 h withdrawal ± TSA treatment. d Quantification of western blots for GABAA subunit membrane expression shown in c. A significant interaction between ethanol and TSA exposure for α1 subunit expression was found (F1,19 = 10.53, p = 0.0043, n = 6, 6, 6, 5) using two-way ANOVA. A significant main effect of ethanol exposure on α4 subunit expression (F1,18 = 10.09, p = 0.0052, n = 6, 5, 5, 6) was found using two-way ANOVA. Values shown represent % control, normalized to β-actin. e Representative blots of H3 PTMs in the chromatin fraction after chronic ethanol exposure ± TSA treatment. f Quantification of blots shown in e. TSA prevents chronic ethanol-induced reductions in global H3 acetylation and H3K9Ac. A significant interaction was found between ethanol exposure and TSA exposure for pH3Ac (F1,20 = 18.38, p = 0.0004, n = 6), and H3K9Ac (F1,20 = 8.212, p = 0.0096, n = 6). There was a significant main effect of TSA exposure for H3K14Ac (F1,18 = 24.41, p = 0.0001, n = 6, 5, 5, 6) found using two-way ANOVA. Two-way ANOVA’s were followed by Bonferroni’s post hoc tests, *p < 0.05, ***p < 0.001, ****p < 0.0001. Data are represented as mean ± SEM
Chronic ethanol exposure increases HDAC2 and HDAC3 associated with chromatin and the Gabra1 promoter and decreases acetylated H3 at the Gabra1 promoter
We next evaluated if decreases in histone acetylation were due to altered HDAC expression. Analysis by qPCR revealed that ethanol exposure upregulated Hdac2 (1.27 ± 0.07 fold control) in the cortex, which trended toward a reversal by the administration of TSA (Fig. 2a, 1.03 ± 0.10 fold control). Hdac3 was also significantly upregulated following chronic ethanol exposure (2.02 ± 0.13 fold control), and this effect was prevented by the administration of TSA (1.32 ± 0.18 fold control, p < 0.01) (Fig. 2a). No changes in other Hdac isoforms (Hdac1, Hdac4-11) were found. Observed changes in transcript levels correlated well with HDAC2 and HDAC3 protein expression in the chromatin fraction (HDAC2 135.4 ± 5.6% control, p < 0.001; HDAC3 178.9 ± 15.2% control, p < 0.01) after chronic ethanol exposure (Fig. 2b). However, HDAC1, another class I HDAC [32] did not exhibit increased transcript or increased expression in the chromatin fraction after chronic ethanol exposure (Fig. 2a, b).

Chronic ethanol exposure increases HDAC2 and HDAC3 both in the chromatin fraction and at the Gabra1 promoter likely facilitating decreases in H3 acetylation at the Gabra1 promoter. a qPCR analysis of the 11 Hdac isoforms and the effect of TSA. Chronic ethanol administration selectively upregulates Hdac2 and Hdac3 transcript levels. A significant main effect of ethanol exposure was found for Hdac2 (F1,20 = 13.10, p = 0.0017, n = 7, 7, 5, 5) while a significant interaction was found for Hdac3 and TSA (F1,18 = 6.233, p = 0.0225, n = 7, 5, 5, 5) using two-way ANOVA. b Chronic ethanol exposure increases HDACs in the chromatin fraction. HDAC2 (t10 = 5.733, p = 0.0002) and HDAC3 (t10 = 3.605, p = 0.0048) were significantly increased using Student’s unpaired two-tailed t test. c TSA prevents changes in histone acetylation associated with the Gabra1 promoter and TSS. A significant main effect of TSA on acetylation at the Gabra1 promoter (F1,16 = 8.840, p = 0.0090, n = 4, 5, 5, 6) and a significant interaction between ethanol exposure and TSA exposure at the Gabra1 TSS (F1,15 = 7.570, p = 0.0148, n = 4, 5, 5, 6) was found using two-way ANOVAs. d Chronic ethanol administration and withdrawal does not change association of the H3K9me3 with the Gabra1 promoter. e ChIP assays reveal that chronic ethanol exposure increases HDAC2 association with the Gabra1 promoter, but not the Gabra4 or Gabrg2 promoters; this effect is blocked by TSA administration. There was a significant interaction between ethanol and TSA exposure for HDAC2 associated with the Gabra1 promoter (F1,16 = 7.353, p = 0.0154, n = 5) using two-way ANOVA. f Chronic ethanol exposure increases HDAC3 enrichment at the Gabra1 promoter, but not Gabra4 or Gabrg2 promoters, and this effect is blocked by the administration of TSA. There was a significant interaction between ethanol and TSA exposure (F1,20 = 46.05, p < 0.0001, n = 6) found using two-way ANOVA. Two-way ANOVAs were followed by Bonferroni’s post hoc tests, *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001. Data are represented as mean ± SEM.
We next evaluated whether ethanol altered histone acetylation associated with the Gabra1 and Gabra4 promoters using chromatin immunoprecipitation. Chronic ethanol gavage caused a decrease in H3Ac associated with the promoter region of Gabra1 (Fig. 2c, 0.05 ± 0.01% input) as well as the transcription start site (TSS) of Gabra1 (0.17 ± 0.04% input, p < 0.05). TSA administration prevented decreases in the acetylation after chronic ethanol exposure at both the Gabra1 promoter (0.25 ± 0.05%, p < 0.05) and the Gabra1 TSS (1.13 ± 0.34% input, p < 0.0001). We did not observe a change in H3Ac at the Gabra1 distal exon (exon 5). Further, we did not observe changes in histone acetylation at the Gabra4 or Gabrg2 promoters (Fig. 2c). No change in the repressive methylation mark H3K9me3 was observed at the Gabra1 promoter after chronic ethanol exposure or TSA administration (Fig. 2d).
We next evaluated if HDAC2 or HDAC3 association with the Gabra1 promoter was altered after 14-day chronic ethanol exposure and withdrawal. ChIP assays revealed that chronic ethanol exposure increases HDAC2 association with the Gabra1 promoter (0.25 ± 0.01% input, p < 0.0001) that was prevented by TSA (0.15 ± 0.01% input, p < 0.001, Fig. 2e). Similarly, HDAC3 association with the Gabra1 promoter was also increased after chronic ethanol exposure (0.18 ± 0.02% input, p < 0.0001) and this effect was prevented by TSA (0.07 ± 0.03% input, p < 0.0001, Fig. 2f). There was no change in HDAC2 or HDAC3 association with either the Gabra4 promoter or the Gabrg2 promoter after chronic ethanol exposure or TSA administration (Fig. 2e, f).
TSA administration prevents changes in gene expression of Gabra1, Hdac2, and Hdac3 in the mPFC, and prevents changes in GABAAR function measured via slice electrophysiology
Since TSA prevented changes in Gabra1 expression in whole cortex, we next evaluated whether these changes occurred in the mPFC, a region of cortex that is dysregulated in alcohol dependence [24, 25]. Sprague Dawley rats were treated for 14 days with 5 g/kg ethanol (i.g.) and the mPFC was microdissected and processed for qPCR analysis (Fig. 3a). The results were similar to those found in whole cerebral cortex, as Gabra1 expression was decreased (Fig. 3b, 0.47 ± 0.05 fold control) and this was prevented by the administration of TSA (Fig. 3b, 0.87 ± 0.06, p < 0.01). We next evaluated if Hdac2 and Hdac3 were upregulated in mPFC. Hdac2 was increased after chronic ethanol gavage (Fig. 3c, 1.8 ± 0.09 fold control, p < 0.01), but TSA treatment did not prevent this upregulation. Hdac3 was also increased after chronic ethanol gavage (Fig. 3d, 1.84 ± 0.10 fold control, p < 0.0001) and this change was prevented by the administration of TSA (Fig. 3d, 1.10 ± 0.05 fold control, p < 0.0001). We next performed slice electrophysiology to determine if there were cell specific effects in pyramidal cells in the prelimbic mPFC (layer V and VI), including changes in GABAergic transmission as determined by sIPSCs. We found a significant decrease in τ1 decay time (Fig. 3f, 8.60 ± 0.370 ms, p < 0.05), indicative of decreased post-synaptic inhibitory activity, that was prevented by the administration of TSA (Fig. 3f, 11.98 ± 0.59 ms, p < 0.01). We did not find any other significant changes in sIPSC characteristics for any treatment group after either ethanol exposure or TSA administration (Supplementary Table 3).

Chronic ethanol exposure-induced changes in Gabra1 expression and GABAA-R function in prelimbic medial PFC are prevented by TSA. a Schematic showing experimental paradigm of ethanol exposure and TSA administration, followed by PFC microdissection or mPFC slice electrophysiology. b qPCR analysis reveals that changes in Gabra1 expression caused by chronic ethanol gavage are prevented by the administration of TSA. There was a significant interaction between ethanol and TSA exposure for Gabra1 expression (F1,20 = 12.67, p = 0.0020, n = 6). c qPCR analysis reveals that increased Hdac2 expression caused by chronic ethanol gavage is prevented by the administration of TSA. There was a significant interaction between ethanol and TSA exposure for Hdac2 expression (F1,19 = 9.096, p = 0.0071, n = 6, 6, 6, 5). d qPCR analysis reveals that increased Hdac3 expression caused by chronic ethanol gavage is prevented by the administration of TSA. There was a significant interaction between ethanol and TSA exposure for Hdac3 expression (F1,19 = 30.88, p < 0.0001, n = 6, 6, 6, 5). e Representative composite traces for sIPSCs measured in layer V and VI in prelimbic mPFC slices. f Changes in sIPSC decay τ1 recorded in the mPFC caused by chronic ethanol exposure are prevented by the administration of TSA. There was a significant effect of TSA on decay τ1 (F1,48 = 12.68, p = 0.0008, n = 14, 13, 16, 9). Two-way ANOVAs were followed by Bonferroni’s post hoc tests, *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001. Data are represented as mean ± SEM.
TSA decreases voluntary ethanol consumption, and prevents effects of voluntary drinking on Gabra1, Hdac2, and Hdac3 expression in the PFC
Since TSA prevented involuntary ethanol-induced changes in gene expression, HDAC activity and behavior following ethanol administration, we examined whether voluntary ethanol consumption would produce similar changes using an intermittent two-bottle choice (I2BC) paradigm using Wistar rats (Fig. 4a) [28]. We injected TSA (2 mg/kg, i.p.) 2 h before drinking on the 14th drinking session. We then monitored drinking for another three sessions. In three of the four sessions following TSA administration, there was a significant decrease in ethanol consumption (Fig. 4b), but only the 14th session showed a significant decrease in alcohol preference (data not shown). There was no significant difference in water consumption on any day between the two groups (data not shown).

Voluntary ethanol consumption alters Gabra1, Hdac2, and Hdac3 in the mPFC, and TSA prevents ethanol-induced changes in voluntary consumption. a Schematic demonstrating two-bottle intermittent ethanol access paradigm. Wistar rats were given free access to bottles of EtOH (15% w/v) and water for 24 h on Monday, Wednesday, and Friday in a reverse light-cycle room and allowed to reach baseline drinking levels (approximately eight sessions). Before session 14, rats received either a vehicle or TSA injection; on session 18, rats were given access to EPM and evaluated for anxiety-like behavior. b TSA administration decreased the amount of ethanol consumed. Each session was analyzed individually using a Student’s t test. n = 13, 14. c qPCR analysis reveals that voluntary ethanol consumption decreases Gabra1 expression and TSA administration prevents this effect. One-way ANOVA analysis found that there was a significant difference between ethanol/vehicle- and ethanol/TSA-treated animals for Gabra1 expression (F2,18 = 43.24, p < 0.0001, n = 7). d qPCR analysis reveals that increased Hdac2 expression caused by voluntary ethanol consumption is prevented by the administration of TSA. One-way ANOVA analysis showed a significant difference between ethanol/vehicle- and ethanol/TSA-treated animals for Hdac2 expression. (F2,15 = 1264, p < 0.0001, n = 6). e qPCR analysis reveals that increased Hdac3 expression caused by voluntary ethanol consumption is prevented by the administration of TSA. One-way ANOVA analysis found that there was a significant difference between ethanol/vehicle- and ethanol/TSA-treated animals for Hdac3 expression (F2,15 = 454.1, p < 0.0001, n = 6). *p < 0.05, ****p < 0.0001. Data are represented as mean ± SEM.
We next analyzed whether I2BC would also cause a decrease in Gabra1 expression in the mPFC with qPCR and found that voluntary drinking decreased Gabra1 expression (Fig. 4c, 0.64 ± 0.02 fold control) and this was prevented by the administration of TSA (1.015 ± 0.02 fold control, p < 0.0001). We then analyzed Hdac2 and Hdac3 expression. We found that voluntary drinking increased Hdac2 expression in the PFC (Fig. 4d, 2.90 ± 0.06 fold control) and this was prevented by the administration of TSA (Fig. 4d, 0.45 ± 0.01, p < 0.0001). Hdac3 expression was also increased in the PFC after voluntary drinking (Fig. 4e, 2.25 ± 0.05 fold control) and prevented by the administration of TSA (Fig. 4e, 0.93 ± 0.02 fold control, p < 0.0001).
TSA administration prevents chronic ethanol-induced cross-tolerance to zolpidem and changes in open-field behavior
To determine if the chronic ethanol exposures had behavioral sequelae that were indicative of changes in GABAAR expression and alcohol dependence that involved the mechanisms elucidated above, we assessed whether TSA would prevent changes in functional activity of the GABAAR α1 receptor subtype by examination of zolpidem-induced LORR [9, 33] as well as a more general behavior disrupted by alcohol dependence (open-field locomotor activity) [34, 35] (Fig. 5a). Finally, we used the elevated plus maze to examine potential changes in anxiety-like behavior and the effect of TSA, since anxiety is a prominent symptom of alcohol withdrawal that is observed in rodent models of alcohol dependence [19, 30]. Zolpidem hypnosis (60 mg/kg, i.p.) was decreased following 14-day ethanol exposure indicating cross-tolerance (Fig. 5b, 52 ± 6.3% control, p < 0.01) and this effect was prevented by the administration of TSA (Fig. 5b, 89.64 ± 9.11% control, p < 0.05). We next assessed locomotor activity and center-time behavior using open-field chambers (Fig. 5c, d). Ethanol administration for 13 days with 24 h withdrawal decreased both the percentage of center-time (Fig. 5c, 0.98 ± 0.14%) and total locomotor activity (Fig. 5d, 1979 ± 166 cm), and both effects were prevented by the administration of TSA (Fig. 5c, 3.00 ± 0.73%, p < 0.05; Fig. 5d, 2762 ± 246 cm, p < 0.01). Following voluntary ethanol consumption (Fig. 5e), we found a decrease in the total open-arm time (Fig. 5f, 4.96 ± 1.07% total time, p < 0.05), and this change was prevented by the administration of TSA (Fig. 5f, 26.34 ± 7.05% total time, p < 0.01) in the elevated plus maze. We did not observe any decrease in total distance traveled in the elevated plus maze after I2BC (Fig. 5g).

TSA prevents chronic alcohol-induced changes in zolpidem hypnosis and open-field center time, locomotor behavior, and elevated plus maze. a Schematic showing the experimental timeline of zolpidem LORR and open-field assay during gavage paradigm. b Zolpidem-induced loss of righting reflex, expressed as % control. A significant interaction between ethanol and TSA exposure (F1,25 = 6.740, p = 0.0156, n = 8, 7, 7, 7) was found using two-way ANOVA. c Center time in open-field assay, expressed as % total locomotor activity time. A significant interaction was found between ethanol and TSA exposure (F1,34 = 12.41, p = 0.0012, n = 10, 9, 9, 10) using two-way ANOVA. d Total locomotor activity in open-field assay. A significant interaction was found between ethanol and TSA exposure (F1,34 = 11.05, p = 0.0021, n = 10, 9, 9, 10) using two-way ANOVA. e Schematic demonstrating two-bottle intermittent ethanol access paradigm and elevated plus maze testing-day. f On session 18, rats were tested on the elevated plus maze in lieu of ethanol access. Open-arm time was reduced in rats that consumed ethanol for 17 sessions compared to rats that never had ethanol access. TSA administration prevented decreases in open-arm time in the EPM in rats that had alcohol access. Significance was determined using one-way ANOVA (F2,24 = 7.022, p = 0.0042, n = 9, 9, 8). Results are expressed as % open-arm time out of total time. g There was no change in total locomotor activity in elevated plus maze for any exposure group. One and two-way ANOVAs were followed by Bonferroni’s post hoc tests, *p < 0.05, **p < 0.01, ****p < 0.0001. Data are represented as mean ± SEM.
Further investigation of the effects of acute ethanol exposure and acute ethanol withdrawal suggest that transcriptional regulation of Gabra1 is dependent upon chronic exposure to alcohol since acute exposure and acute withdrawal did not cause changes Gabra1 expression, H3Ac, or HDAC1-3 expression in the cerebral cortex (Supplementary Figure 1).
Discussion
In this study, we evaluated whether an epigenetic mechanism regulated GABAAR α1 and α4 expression in two different rodent models of ethanol dependence and determined if these changes could be prevented by the administration of the HDAC inhibitor TSA. Our results indicate that chronic ethanol exposure and withdrawal causes a decrease in α1 expression by histone deacetylation adjacent to the Gabra1 gene promoter region and transcription start site, which is facilitated by the recruitment of HDAC2 and HDAC3 and that these changes can be prevented by the administration of TSA. Evaluation of cortical circuitry involved in addiction (the mPFC) revealed similar changes in both Gabra1 expression and Hdac2 and Hdac3 expression, and also functional changes in GABAergic neurotransmission as evidenced by changes in sIPSC decay-tau, all of which were reversed after TSA administration. Cross-tolerance to GABAergic drugs is a known symptom of alcohol dependence in humans and rodents, and acute TSA administration prevented this effect as evidenced by the reversal of decreases in zolpidem-induced LORR caused by chronic ethanol exposure. In addition, several behavioral phenotypes of alcohol dependence were prevented by acute administration of TSA, including open-field activity, anxiety-like behavior in the elevated plus maze, and ethanol consumption in a two-bottle choice paradigm.
Epigenetic changes in GABAAR α1 expression are important in alcohol dependence
Chronic ethanol exposure has long been known to cause decreased α1 subunit expression using several different modes of ethanol exposure including gavage [4] or liquid diet [6] in rodent models that recapitulate symptoms observed in human patients with alcohol use disorders. The GABAAR α1 subunit is important in the chronic effects of ethanol, as chronic ethanol-induced decreases in α1 expression are associated with cross-tolerance to benzodiazepines and non-benzodiazepine hypnotics such as zolpidem, increased seizure susceptibility, decreased LORR reflex induced by GABAergic hypnotics and ethanol, and tolerance to the sedative effects of ethanol [4, 6, 9, 13, 36, 37] and restoring GABAergic signaling in alcohol dependence has long been recognized as a relevant treatment strategy for withdrawal symptoms. Previous studies have largely focused on restoring GABAAR α1 surface expression as an alternative to modulating GABAergic function with benzodiazepines, via modulating protein kinase interactions with α1-containing GABAARs to prevent receptor internalization [15, 38,39,40]. The current study suggests a previously unexplored avenue that GABAAR adaptations occur through an epigenetic mechanism via histone deacetylation of the Gabra1 promoter and transcriptional start site by HDAC2 and HDAC3. A behavioral correlate of these finding is that we found that TSA prevented changes in zolpidem-induced loss of righting reflex caused by chronic ethanol exposure.
An important point that supports the generalizability of these findings is that two rat strains with different genetic backgrounds (Sprague Dawley and Wistar) showed decreased α1 expression and increased Hdac2 and Hdac3 expression. Wistar rats are considered a high drinking strain and therefore were used for the I2BC, while Sprague Dawleys are considered a low drinking strain and were used for involuntary ethanol exposure via gavage. Previous studies in humans have suggested that ~50% of the risk of alcohol dependence is genetic and the remaining 50% is due to other factors, and our current study suggests that the epigenetic modifications that we observed in both the cortex and PFC are specific to ethanol exposure and not due to differences in genetic background or innate alcohol preference. Observations that ethanol reduces GABAAR function and expression through multiple molecular mechanisms and in two different rat strains in two different ethanol-exposure paradigms underscore the significance of these receptors in ethanol dependence.
Chronic ethanol changes HDAC expression in both cortex and neurocircuitry implicated in addiction
The PFC is known to be a critical brain region involved in alcohol dependence [24, 41, 42]. The current study identified that decreases in Gabra1 and increases in Hdac2 and Hdac3 also occur in the PFC. The PFC controls executive function and disinhibition of the PFC, via alterations in neurotransmission caused by an imbalance of GABAergic and glutamatergic systems, is thought to contribute to the development of alcohol dependence in rodent models [42] and human patients with alcohol use disorders [43]. Increased activity in the PFC, possibly through decreased GABAergic signaling, contributes to anxiety in humans [44] and dysregulation in projections from the PFC contributes to anxiety-like behavior in rodents [45]. Previous studies have demonstrated that chronic ethanol exposure causes decreases in GABAergic inhibition in the mPFC [46], hippocampus [4], and amygdala [47] that appear to involve loss of α1 expression. We observed changes in synaptic transmission via decreases in decay-tau that are potentially due to decreased α1 expression and/or increased α4 expression [16, 48, 49]. Decreases in α1 expression and increases in α4 expression are thought to mediate changes in benzodiazepine sensitivity, seizure susceptibility, and increased anxiety-like behavior [9, 14, 50, 51]. The fact that we found both hypoactivity and decreased center-time activity following chronic ethanol exposure may suggest that anxiety-like behavior was not observed. To mitigate this possibility, we expressed the center time as a fraction of total locomotion, thereby normalizing the data to account for reduced general locomotion. The data suggest that chronic ethanol exposure reduces both total locomotion and the relative proportion of center time, consistent with evidence for anxiety-like behavior as well as reduced exploration. This data may indicate another dependence phenotype that involves decreased locomotor behavior similar to what has been reported by other groups [34, 35].
In addition, we identified upregulation of Hdac2 and Hdac3 mRNA transcripts in the PFC. Changes in Hdac2 expression causes changes in behavioral outputs in other brain regions [18, 52]. However, these results show that changes in Hdac3 also occur in the PFC, suggesting that there are brain region specific changes relative to the previously reported changes HDAC2 expression in the amygdala. These results contribute to a growing body of work that suggests TSA is a useful treatment modality that can prevent ethanol-induced changes in circuitry involved in alcohol dependence.
Histone deacetylases inhibitors as a useful treatment for alcohol dependence
Chronic alcohol exposure causes numerous changes in the transcriptome in human alcoholics [53] and rodents [54] suggesting that TSA or other HDAC inhibitors target multiple different pathways involved in alcohol use disorders [19, 55,56,57,58]. Our data indicate that ethanol induces specific changes in H3 acetylation at Gabra1 promoter, but not the Gabra4 and Gabrg2 promoters, suggesting that histone deacetylation is specific to only certain genes. The demonstration of epigenetic modulation of the Gabra1 promoter by ethanol in vivo agrees with our previously published data showing that targeted increases in histone acetylation associated with the Gabra1 promoter using a CRISPR Cas9 strategy prevent ethanol-induced decreases in Gabra1 expression in vitro [21] suggesting that more targeted interventions can prevent changes in Gabra1 expression. Moreover, we and several other groups have demonstrated that chronic ethanol exposure causes decreases in global histone acetylation and these changes are thought to facilitate changes in gene expression, which likely contribute to alcohol use disorders [19, 57,58,59,60], suggesting that histone deacetylation may be a ubiquitous mechanism for ethanol effects. Improved selectivity of drugs targeting aberrant gene expression caused by alcohol abuse and dependence may be improved by our understanding of the epigenetic mechanisms that are dysregulated. Previous studies have implicated HDAC2 in alcohol dependence in rodents [52, 60, 61] and alcohol use disorders in humans [61, 62]. The current study found changes in HDAC2, but also found changes in HDAC3, suggesting that HDAC3 may be a previously unrecognized target for the treatment of alcohol dependence. HDAC2 and HDAC3 are differently expressed in the cortex, with HDAC3 being the more highly expressed of the two isoforms and the most highly expressed isoform in the brain [32]. HDAC2 and HDAC3 are not commonly found together in the same signaling complex, suggesting that there may be two different pathways recruited by chronic ethanol exposure. Future studies will need to address this possibility.
Our results suggest that HDAC inhibitors in the hydroxamate class (e.g., TSA and the FDA approved Vorinostat) may be useful in the treatment of alcohol dependence. Our results indicate that even acute TSA treatment can normalize HDAC2/3 mRNA expression suggesting that this may be the mechanism of action of TSA to prevent changes in global H3 acetylation and H3 acetylation associated with the Gabra1 promoter. RNA-sequencing studies of GABAAR α1 knockout mice find that there are considerable changes to gene regulatory networks [63] suggesting that decreases in GABAAR α1 expression are also important for changes beyond just the GABAergic system and implicating the need for a broad-based HDAC inhibitor, such as TSA, to prevent global changes in gene transcription induced by downregulation of GABAAR α1 receptors. Therefore, HDAC inhibitors may have therapeutic value in other disease states where GABAAR α1 expression is dysregulated, including epilepsy [64], autism [65], schizophrenia [66], Alzheimer’s disease [67], and depression [68]. However, long-term treatment with HDAC inhibitors in alcohol use disorders and other disease states still need to be evaluated.
The present paper demonstrates that acute TSA can be used to prevent drinking behaviors in an I2BC paradigm, withdrawal behaviors, and the loss of GABAergic inhibition in the mPFC and decreases in α1 expression in the PFC and whole cortex. The results presented here may inform the development of better therapies utilizing epigenetic pathways for the treatment of alcohol use disorders.
References
Stahre M, Roeber J, Kanny D, Brewer RD, Zhang X. Contribution of excessive alcohol consumption to deaths and years of potential life lost in the United States. Prev Chronic Dis. 2014;11:E109.
Shaw GK, Waller S, Latham CJ, Dunn G, Thomson AD. The detoxification experience of alcoholic in-patients and predictors of outcome. Alcohol Alcohol. 1998;33:291–303.
Miller NS, Gold MS. Management of withdrawal syndromes and relapse prevention in drug and alcohol dependence. Am Fam Physician. 1998;58:139–46.
Cagetti E, Liang J, Spigelman I, Olsen RW. Withdrawal from chronic intermittent ethanol treatment changes subunit composition, reduces synaptic function, and decreases behavioral responses to positive allosteric modulators of GABAA receptors. Mol Pharmacol. 2003;63:53–64.
Tietz EI, Huang X, Weng X, Rosenberg HC, Chiu TH. Expression ofa1, a5 and g2 GABAA receptor subunit mRNAs measured In Situ in rat hippocampus and cortex following chronic flurazepam administration. J Mol Neurosci. 1993;4:277–92.
Devaud LL, Fritschy JM, Sieghart W, Morrow AL. Bidirectional alterations of GABAA receptor subunit peptide levels in rat cortex during chronic ethanol consumption and withdrawal. J Neurochem. 1997;69:126–30.
Devaud LL, Smith FD, Grayson DR, Morrow AL. Chronic ethanol consumption differentially alters the expression of γ-aminobutyric acid type A receptor subunit mRNAs in rat cerebral cortex: competitive, quantitative reverse transcriptase-polymerase chain reaction analysis. Mol Pharmacol. 1995;48:861–8.
Kralic JE, Criswell HE, Osterman JL, O’Buckley TK, Wilkie ME, Matthews DB, et al. Genetic essential tremor in gamma-aminobutyric acidA receptor α1 subunit knockout mice. J Clin Invest. 2005;115:774–9.
Kralic JE, O’Buckley TK, Khisti RT, Hodge CW, Homanics GE, Morrow AL. GABAA receptor α1 subunit deletion alters receptor subtype assembly, pharmacological and behavioral responses to benzodiazepines and zolpidem. Neuropharmacology. 2002;43:685–94.
Liang J, Suryanarayanan A, Abriam A, Snyder B, Olsen RW, Spigelman I. Mechanisms of reversible GABAA receptor plasticity after ethanol intoxication. J Neurosci. 2007;27:12367–77.
Sanna E, Serra M, Cossu A, Colombo G, Follesa P, Cuccheddu T, et al. Chronic ethanol intoxication induces differential effects on GABAA and NMDA receptor function in the rat brain. Alcohol Clin Exp Res. 1993;17:115–23.
Ticku MK, Burch T. Alterations in g-aminobutyric acid receptor sensitivity following acute and chronic ethanol treatments. J Neurochem. 1980;34:417–23.
Kralic JE, Korpi ER, O’Buckley TK, Homanics GE, Morrow AL. Molecular and pharmacological characterization of GABAA receptor α1 subunit knockout mice. J Pharmacol Exp Ther. 2002;302:1037–45.
Gulinello M, Gong QH, Smith SS. Progesterone withdrawal increases the alpha4 subunit of the GABA(A) receptor in male rats in association with anxiety and altered pharmacology - a comparison with female rats. Neuropharmacology. 2002;43:701–14.
Carlson SL, Kumar S, Werner DF, Comerford CE, Morrow AL. Ethanol activation of protein kinase A regulates GABAA alpha1 receptor function and trafficking in cultured cerebral cortical neurons. J Pharmacol Exp Ther. 2013;345:317–25.
Werner DF, Kumar S, Criswell HE, Suryanarayanan A, Fetzer JA, Comerford CE, et al. PKCgamma is required for ethanol-induced increases in GABAA receptor α4 subunit expression in cultured cerebral cortical neurons. J Neurochem. 2011;116:554–63.
Wang Z, Zang C, Rosenfeld JA, Schones DE, Barski A, Cuddapah S, et al. Combinatorial patterns of histone acetylations and methylations in the human genome. Nat Genet. 2008;40:897–903.
Kennedy PJ, Feng J, Robison AJ, Maze I, Badimon A, Mouzon E, et al. Class I HDAC inhibition blocks cocaine-induced plasticity by targeted changes in histone methylation. Nat Neurosci. 2013;16:434–40.
Pandey SC, Ugale R, Zhang H, Tang L, Prakash A. Brain chromatin remodeling: a novel mechanism of alcoholism. J Neurosci. 2008;28:3729–37.
Renthal W, Kumar A, Xiao G, Wilkinson M, Covington HE, Maze I, et al. Genome-wide analysis of chromatin regulation by cocaine reveals a role for sirtuins. Neuron. 2009;62:335–48.
Bohnsack JP, Patel VK, Morrow AL. Ethanol exposure regulates Gabra1 expression via histone deacetylation at the promoter in cultured cortical neurons. J Pharm Exp Ther. 2017;363:1–11.
Van Lint C, Emiliani S, Verdin E. The expression of a small fraction of cellular genes is changed in response to histone hyperacetylation. Gene Expr. 1996;5:245–53.
Freund G, Ballinger WEJ. Decrease of benzodiazepine receptors in frontal cortex of alcoholics. Alcohol. 1988;5:275–82.
Abernathy K, Chandler LJ, Woodward JJ. Alcohol and the prefrontal cortex. Int Rev Neurobiol. 2010;91:289–320.
Tu Y, Kroener S, Abernathy K, Lapish C, Seamans J, Chandler LJ, et al. Ethanol inhibits persistent activity in prefrontal cortical neurons. J Neurosci. 2007;27:4765–75.
Pandey SC, Zhang H, Ugale R, Prakash A, Xu T, Misra K. Effector immediate-early gene arc in the amygdala plays a critical role in alcoholism. J Neurosci. 2008;28:2589–2600.
Sakharkar AJ, Zhang H, Tang L, Shi G, Pandey SC. Histone deacetylases (HDAC)-induced histone modifications in the amygdala: a role in rapid tolerance to the anxiolytic effects of ethanol. Alcohol Clin Exp Res. 2012;36:61–71.
Simms JA, Steensland P, Medina B, Abernathy KE, Chandler LJ, Wise R, et al. Intermittent access to 20% ethanol induces high ethanol consumption in Long-Evans and Wistar rats. Alcohol Clin Exp Res. 2008;32:1816–23.
Bohnsack JP, Carlson SL, Morrow AL. Differential regulation of synaptic and extrasynaptic α4 GABAA receptor populations by protein kinase A and protein kinase C in cultured cortical neurons. Neuropharmacology. 2016;105:124–32.
Walf AA, Frye CA. The use of the elevated plus maze as an assay of anxiety-related behavior in rodents. Nat Protoc. 2007;2:322–8.
Kumar S, Porcu P, Werner DF, Matthews DB, Diaz-Granados JL, Helfand RS, et al. The role of GABAA receptors in the acute and chronic effects of ethanol: a decade of progress. Psychopharmacology. 2009;205:529–64.
Broide RS, Redwine JM, Aftahi N, Young W, Bloom FE, Winrow CJ. Distribution of histone deacetylases 1-11 in the rat brain. J Mol Neurosci. 2007;31:47–58.
Mikolajczak P. Zolpidem involvement on memory and hypnotic effect of ethanol in chronically ethanol-treated rats. Alcohol Alcohol. 1999;34:511–9.
Devaud LL, Bartoo G, Malthankar G. Altered responses to dizocilpine maleate administration in ethanol-withdrawn male and female rats. Alcohol. 2002;28:83–93.
Li J, Bian WL, Xie GQ, Cui SZ, Wu ML, Li YH, et al. Chronic ethanol intake-induced changes in open-field behavior and cal-cium/calmodulin-dependent protein kinase IV expression in nucleus accumbens of rats: naloxone reversal. Acta Pharmacol Sin. 2008;29:646–52.
Blednov YA, Jung S, Alva H, Wallace D, Rosahl T, Whiting PJ, et al. Deletion of the a1 or b2 subunit of GABAA receptors reduces actions of alcohol and other drugs. J Pharmacol Exp Ther. 2003;304:30–6.
Liang J, Cagetti E, Olsen RW, Spigelman I. Altered pharmacology of synaptic and extrasynaptic GABAA receptors on CA1 hippocampal neurons is consistent with subunit changes in a model of alcohol withdrawal and dependence. J Pharmacol Exp Ther. 2004;310:1234–45.
Hodge CW, Mehmert KK, Kelley SP, McMahon T, Haywood A, Olive MF, et al. Supersensitivity to allosteric GABAA receptor modulators and alcohol in mice lacking PKCe. Nat Neurosci. 1999;2:997–1002.
Kumar S, Sieghart W, Morrow AL. Association of protein kinase C with GABAA receptors containing a1 and a4 subunits in the cerebral cortex: selective effects of chronic ethanol consumption. J Neurochem. 2002;82:110–7.
Kumar S, Suryanarayanan A, Boyd KN, Comerford CE, Lai MA, Ren Q, et al. Ethanol reduces GABAA α1 subunit receptor surface expression by a protein kinase C gamma-dependent mechanism in cultured cerebral cortical neurons. Mol Pharmacol. 2010;77:793–803.
Heilig M, Barbier E, Johnstone AL, Tapocik J, Meinhardt MW, Pfarr S, et al. Reprogramming of mPFC transcriptome and function in alcohol dependence. Genes Brain Behav. 2017;16:86–100.
Woodward JJ, Pava MJ. Effects of ethanol on persistent activity and up-states in excitatory and inhibitory neurons in prefrontal cortex. Alcohol Clin Exp Res. 2009;33:2134–40.
Volkow ND, Wang GJ, Hitzemann R, Fowler JS, Wolf AP, Pappas N, et al. Decreased cerebral response to inhibitory neurotransmission in alcoholics. Am J Psychiatry. 1993;150:417–22.
Martin EI, Ressler KJ, Binder E, Nemeroff CB. The neurobiology of anxiety disorders: brain imaging, genetics, and psychoneuroendocrinology. Psychiatr Clin North Am. 2009;32:549–75.
Calhoon GG, Tye KM. Resolving the neural circuits of anxiety. Nat Neurosci. 2015;18:1394–404.
Kroener S, Mulholland PJ, New NN, Gass JT, Becker HC, Chandler LJ. Chronic alcohol exposure alters behavioral and synaptic plasticity of the rodent prefrontal cortex. PLoS ONE. 2012;7:e37541.
Herman MA, Roberto M. Cell-type-specific tonic GABA signaling in the rat central amygdala is selectively altered by acute and chronic ethanol. Addict Biol. 2016;21:72–86.
Liang J, Zhang N, Cagetti E, Houser CR, Olsen RW, Spigelman I. Chronic intermittent ethanol-induced switch of ethanol actions from extrasynaptic to synaptic hippocampal GABAA receptors. J Neurosci. 2006;26:1749–58.
Carlson SL, Bohnsack JP, Morrow AL. Ethanol regulation of synaptic GABAA alpha4 receptors is prevented by protein kinase A activation. J Pharmacol Exp Ther. 2016;357:10–6.
Kralic JE, Wheeler M, Renzi K, Ferguson C, O’Buckley TK, Grobin AC, et al. Deletion of GABAA receptor α1 subunit-containing receptors alters responses to ethanol and other anesthetics. J Pharmacol Exp Ther. 2003;305:600–7.
Smith SS, Gong QH, Hsu F-C, Markowitz RS, Ffrench-Mullen JMH, Li X. GABAA receptor α4 subunit suppression prevents withdrawal properties of an endogenous steroid. Nature. 1998;392:926–30.
Sakharkar AJ, Zhang H, Tang L, Baxstrom K, Shi G, Moonat S, et al. Effects of histone deacetylase inhibitors on amygdaloid histone acetylation and neuropeptide Y expression: a role in anxiety-like and alcohol-drinking behaviours. Int J Neuropsychopharmacol. 2014;17:1207–20.
Farris SP, Arasappan D, Hunicke-Smith S, Harris RA, Mayfield RD. Transcriptome organization for chronic alcohol abuse in human brain. Mol Psychiatry. 2014;20:1438–47.
Hashimoto JG, Forquer MR, Tanchuck MA, Finn DA, Wiren KM. Importance of genetic background for risk of relapse shown in altered prefrontal cortex gene expression during abstinence following chronic alcohol intoxication. Neuroscience. 2011;173:57–75.
Moonat S, Sakharkar AJ, Zhang H, Tang L, Pandey SC. Aberrant histonedeacetylase2–mediated histone modifications and synaptic plasticity in the amygdala predisposes to anxiety and alcoholism. Biol Psychiatry. 2013;73:763–73.
Sakharkar AJ, Vetreno RP, Zhang H, Kokare DM, Crews FT, Pandey SC. A role for histone acetylation mechanisms in adolescent alcohol exposure-induced deficits in hippocampal brain-derived neurotrophic factor expression and neurogenesis markers in adulthood. Brain Struct Funct. 2016;221:4691–703.
Simon-O’Brien E, Alaux-Cantin S, Warnault V, Buttolo R, Naassila M, Vilpoux C. The histone deacetylase inhibitor sodium butyrate decreases excessive ethanol intake in dependent animals. Addict Biol. 2015;20:676–89.
Warnault V, Darcq E, Levine A, Barak S, Ron D. Chromatin remodeling--a novel strategy to control excessive alcohol drinking. Transl Psychiatry. 2013;3:e231.
D’Addario C, Caputi FF, Ekstrom TJ, Di Benedetto M, Maccarrone M, Romualdi P, et al. Ethanol induces epigenetic modulation of prodynorphin and pronociceptin gene expression in the rat amygdala complex. J Mol Neurosci. 2013;49:312–9.
Arora DS, Nimitvilai S, Teppen TL, McElvain MA, Sakharkar AJ, You C, et al. Hyposensitivity to gamma-aminobutyric acid in the ventral tegmental area during alcohol withdrawal: reversal by histone deacetylase inhibitors. Neuropsychopharmacology. 2013;38:1674–84.
López-Moreno JA, Marcos M, Calleja-Conde J, Echeverry-Alzate V, Bühler KM, Costa-Alba P, et al. Histone deacetylase gene expression following binge alcohol consumption in rats and humans. Alcohol Clin Exp Res. 2015;39:1939–50.
Zhou Z, Yuan Q, Mash DC, Goldman D. Substance-specific and shared transcription and epigenetic changes in the human hippocampus chronically exposed to cocaine and alcohol. Proc Natl Acad Sci USA. 2011;108:6626–31.
Ponomarev I, Maiya R, Harnett MT, Schafer GL, Ryabinin AE, Blednov YA, et al. Transcriptional signatures of cellular plasticity in mice lacking the alpha1 subunit of GABAA receptors. J Neurosci. 2006;26:5673–83.
Jansen LA, Peugh LD, Roden WH, Ojemann JG. Impaired maturation of cortical GABAA receptor expression in pediatric epilepsy. Epilepsia. 2010;51:1456–67.
Fatemi SH, Reutiman TJ, Folsom TD, Thuras PD. GABAA receptor downregulation in brains of subjects with autism. J Autism Dev Disord. 2008;39:223–30.
Hoftman GD, Volk DW, Bazmi HH, Li S, Sampson AR, Lewis DA. Altered cortical expression of GABA-related genes in schizophrenia: illness progression vs developmental disturbance. Schizophr Bull. 2013;41:180–91.
Limon A, Reyes-Ruiz JM, Miledi R. Loss of functional GABA(A) receptors in the Alzheimer diseased brain. Proc Natl Acad Sci USA. 2012;109:10071–6.
Poulter MO, Du L, Weaver ICG, Palkovits M, Faludi G, Merali Z, et al. GABAA receptor promoter hypermethylation in suicide brain: implications for the involvement of epigenetic processes. Biol Psychiatry. 2008;64:645–52.
Acknowledgements
We thank Drs. Abigail Agoglia and Anel Jaramillo in the UNC Bowles Center for Alcohol Studies for assistance with open-field locomotor assays, and Dr. Sara Faccidomo in the UNC Bowles Center for Alcohol Studies with for assistance with elevated plus maze. We also thank Dr. Melissa Herman in the UNC Bowles Center for Alcohol Studies for help with PFC dissections. This work was supported by the National Institute of Alcohol Abuse and Alcoholism Grant AA11605 (ALM) and the Bowles Center for Alcohol Studies.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Electronic supplementary material
41386_2018_34_MOESM2_ESM.pdf
Supplementary Figure 1. Acute ethanol intoxication and acute alcohol withdrawal do not induce changes in GABAAR expression, H3 PTMs, or HDAC association with chromatin
Rights and permissions
About this article

Cite this article
Bohnsack, J.P., Hughes, B.A., O’Buckley, T.K. et al. Histone deacetylases mediate GABAA receptor expression, physiology, and behavioral maladaptations in rat models of alcohol dependence. Neuropsychopharmacol 43, 1518–1529 (2018). https://doi.org/10.1038/s41386-018-0034-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41386-018-0034-8
This article is cited by
-
Epigenetic drugs and psychedelics as emerging therapies for alcohol use disorder: insights from preclinical studies
Journal of Neural Transmission (2024)
-
HDAC1-mediated regulation of GABA signaling within the lateral septum facilitates long-lasting social fear extinction in male mice
Translational Psychiatry (2023)
-
Sex- and subtype-specific adaptations in excitatory signaling onto deep-layer prelimbic cortical pyramidal neurons after chronic alcohol exposure
Neuropsychopharmacology (2021)
-
Studying alcohol use disorder using Drosophila melanogaster in the era of ‘Big Data’
Behavioral and Brain Functions (2019)
