
Abstract
Oral cholera vaccination is used to induce immune responses in the intestines to protect against cholera infection. However, oral vaccination may also affect immune responses in other mucosal tissues. To study this, tissue-specific homing potential and kinetics of B-cell responses were characterized after oral cholera vaccination. Healthy adult volunteers received two doses of Dukoral® and blood, saliva, nasal wash, and fecal samples were collected over time to detect vaccine-specific antibodies. Additionally, homing potential of lymphocytes to small intestine, colon, airways, skin, and periphery was measured by expression of Integrin β1 and β7, CCR9, CCR10, CCR7, and CLA. After vaccination, antibody responses to cholera toxin B (CTB) and Dukoral® were detected in serum and nasal wash. CTB-specific memory B cells in peripheral blood and tissue homing profiles of memory B cells peaked at day 18. IgA+ memory B cells expressed markers that enable homing to the airways and colon, while IgA− memory B cells primarily expressed small-intestine-homing markers. These data show that oral cholera vaccination has a differential effect on immune responses in various mucosal sites, including the respiratory tract.
Similar content being viewed by others

Introduction
Mucosal surfaces are frequently exposed to potentially harmful microorganisms. As a result, mucosal tissues can be infected by many different pathogens, such as Influenza A, rhinovirus, respiratory syncytial virus (RSV), and Streptococcus pneumoniae in the respiratory tract and Vibrio Cholerae, Rotavirus, Norovirus, Salmonella Typhi, Escherichia coli, and Campylobacter jejuni in the gastrointestinal tract.
Parenteral vaccination predominantly elicits systemic immune responses to the vaccine, whereas mucosal vaccination via the oral or intranasal route induces adaptive immune responses targeted to the mucosal tissues and intestinal lumen to inhibit colonization and invasion by pathogens.1,2,3 Oral vaccines used for prevention of diarrheal diseases caused by Vibrio Cholerae, Salmonella Typhi, and Rotavirus4 are commonly used for populations at risk. The only intranasal vaccine licensed for use in humans is against Influenza A and B5 and other licensed vaccines for respiratory infections are administered parenterally. In this respect, there is much interest in the potential for oral vaccination to elicit a more protective immune responses in the upper respiratory tract than parenteral vaccines.
Oral vaccines primarily interact with the immune system through the tonsils of Waldeyer’s ring in the oral cavity or via the Peyer’s patches in the small intestine. In addition, other structures in the gut associated lymphoid tissue (GALT) and isolated lymphoid follicles in the colon can also play a role in immune responses to oral vaccines. Oral vaccination is known to induce larger amounts of vaccine-specific IgA in the small intestine, colon, mammary glands, salivary glands, and nasal cavity, than in the genital tract, tonsils, and the lower respiratory tract.6,7,8,9 Conversely, vaccination in the rectum and female genital tract mainly induces local vaccine-specific IgA production.10,11,12 Part of this compartmentalization of the mucosal immune system may be explained by tissue-specific homing properties of B and T cells. Tissue-specific homing of lymphocytes is dependent on their expression pattern of homing receptors that bind addressins expressed on the endothelium of the target tissue.13 Examples are binding of Integrin α4β1 to vascular cell adhesion molecule (VCAM)-1 and Integrin α4β7 to mucosal addressin cell adhesion molecule (MAdCAM)-1.14 Furthermore, locally produced chemokines (e.g., CCL25, CCL28) attract lymphocytes via their chemokine receptors (CCR915,16 and CCR10,14 respectively) and direct them into the tissue.17,18 It is not known to what extent an oral cholera vaccine can induce a local mucosal immune response by modulating the homing profile of peripheral lymphocytes. Another factor that can determine lymphocyte trafficking and tissue homing is the location of the initial immune activation, as was recently described for plasmablasts.19,20 In addition, homing is influenced by external factors, such as stromal factors and food components, such as retinoic acid.21,22
The aim of this study was to evaluate whether oral cholera vaccination is able to induce potent IgA and IgG responses in serum, as well as at mucosal surfaces. In addition, the kinetics of these antibody responses and the tissue homing marker expression profiles of memory B cells (in contrast to plasmablasts as described recently)19 toward airways, small intestine, colon, periphery, and skin (Supplementary figure S1) was investigated. This is to our knowledge the first study that investigated the kinetics of extensive homing profiles of memory B cells upon oral vaccination.
Results
Oral vaccination with Dukoral® induces vaccine-specific IgA and IgG antibodies
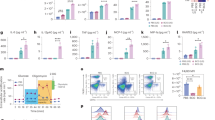
To measure the humoral response to oral cholera vaccination, vaccine-specific IgA and IgG (Dukoral® and cholera toxin B (CTB)- specific antibodies) were measured in serum and mucosal fluids. The median values and ranges are depicted in Fig. 1. Vaccine-specific antibody levels in serum increased over time, with a maximum at day 28 post vaccination. Levels of both Dukoral®-specific IgA (Fig. 1a) and cholera toxin B (CTB)-specific IgA (AU/ml) (Fig. 1b) were significantly increased at days 18 and 28. Vaccine-specific IgG (Fig. 1c, d) in serum was increased significantly at day 28. Dukoral®-specific IgA in fecal extracts and vaccine-specific IgA in nasal wash were just above the detection limit (0.12 AU/ml) and did not increase over time. Dukoral®-specific IgG was not detectable in fecal extracts. The concentration of vaccine specific IgG in nasal wash was significantly increased at day 28. Saliva samples were not available for all donors at all time points. Low levels of CTB-specific IgA were detected in saliva and showed no significant vaccination response. CTB-specific IgG in saliva was below the detection limit. In conclusion, vaccine-specific antibodies were induced in serum (IgA and IgG) and nasal wash (IgG) following oral vaccination

Serum of healthy volunteers (n = 14) was measured on days 0, 7, 14, 18, and day 28. Dukoral-specific IgA (a), CTB-specific IgA (b), Dukoral-specific IgG (c), and CTB-specific IgG (d) were measured over time. Data are shown as dot plots with mean ± SEM in AU/ml. Serum IgA was diluted 75×, serum IgG 300×. Repeated measures ANOVA was performed after 10 log transformation of the data. Stars indicate significant differences at time points compared to day 0. *p < 0.05; **p < 0.01; ***p < 0.001
Cholera toxin B-specific memory B cells in blood increase after oral vaccination
In addition to measuring the levels of specific antibodies in serum, the induction of CTB-specific B cells in peripheral blood was investigated. The presence of IgA+ and IgG+ CTB-specific B cells in peripheral blood mononuclear cells (PBMCs) was analyzed by flow cytometry (Fig. 2a). The frequency of CTB-specific IgA+ as well as IgG+ memory B cells significantly increased after oral vaccination and peaked at day 18 (Fig. 2b, c).

Kinetics of cholera toxin B (CTB)-specific memory B cells in peripheral blood. PBMCs of healthy volunteers (n = 14) were stained and measured on days 0, 14, 18, and 28. CTB-specific IgA+ and IgG+ memory B cells were analyzed by flow cytometry (a) and are shown as #CTB-specific IgA+ memory B cells/1 × 106 memory B cells at days 0, 14, 18, and 28 (b, c). Data are shown in Whisker plots with a median, 25 and 75% quartile, and 5–95% confidence interval. Repeated measures ANOVA was performed *p < 0.05; **p < 0.01; ***p < 0.001
The B cells identified using this gating strategy are isotype-switched CD19+ memory B cells. To compare the effect of oral vaccination on memory B cells with the effect on plasmablasts, we devised an alternative gating strategy to study plasmablasts (Supplementary Figure S2). Using this strategy, the kinetics of IgG+ as well as IgA+ and IgG−IgA− (IgM) plasmablasts were analyzed. In contrast to the results shown for IgA+ and IgG+ memory B cells, the numbers of IgA+ and IgG+ plasmablasts in peripheral blood did not increase significantly after vaccination on the days studied. The number of IgA−IgG− plasmablasts seemed to decrease after vaccination, but this decrease was also not significant.
Based on the fact that we only see a clear vaccination response in the memory B cells all further analyses were performed on the memory B cells. Oral vaccination induces markers associated with homing to the colon on IgA+ memory B cells and to the small intestine on IgA− memory B cells.
In addition to the presence of CTB-specific memory B cells, the kinetics of memory B cells with homing marker profiles consistent for homing to different organs was analyzed with panels of antibodies for relevant homing markers (Supplementary Figure S1). In Fig. 3, the overall gut homing potential of IgA+ and IgA− memory B cells (CD19+CLA− Integrin β7+), based on integrin β7+ expression alone, was compared with chemokine receptor expression of CCR9 alone(CD19+CLA−CCR9+) and CCR10 alone (CD19+CLA−CCR10+). Furthermore, homing potential toward the colon (CD19+CLA-Integrinβ7+CCR10+CCR9−) and small intestine (CD19+CLA−Integrinβ7+CCR10−CCR9+) was compared by combining the expression of integrin β7 and chemokine receptors. Upon vaccination, the percentage of IgA+ memory B cells with overall homing potential to the gut, colon, and small intestine, and also CCR9 or CCR10 expression alone increased. Overall, the gut homing potential of IgA+ memory B cells, based on Integrin β7+ expression alone, was significantly increased, most profoundly at day 14. The clearest vaccination response for IgA+ memory B cells was observed, when combining Integrin β7+ expression with CCR10 as markers for colon homing potential, with a peak at day 18 (p < 0.001, Fig. 3b), which corresponded with a peak in the number of CTB-specific IgA+ memory B cells. The same kinetics were observed when IgA+ memory B cells were only gated for the expression of CCR10. When combining Integrin β7+ expression of IgA+ memory B cells with CCR9, a significant increase in homing potential to the small intestine was observed at day 14, while this increase was smaller for CCR9 expression alone. For IgA− memory B cells, the kinetics and tissue selectivity of the vaccination-induced homing response were different. Homing potential of IgA− memory B cells towards the colon was only significantly increased at day 18 and a similar trend was observed for single CCR10+ IgA− memory B cells. Homing potential toward the small intestine was much more profound for IgA− memory B cells as compared to IgA+ memory B cells, with a peak at day 18, a trend also observed for single CCR9+ IgA− memory B cells.

Kinetics of IgA+ and IgA− memory B cells with a gut, colon, and small intestine homing potential or expression of CCR9 alone and CCR10 alone. PBMCs of healthy volunteers (n = 14) were analyzed by flow cytometry on days 0, 14, 18, and 28. a Gating strategy used to differentiate between IgA+ (b) and IgA− (c) memory B cells with a gut (CD19+CLA−Integrinβ7+), colon (CD19+CLA−Integrinβ7+CCR10+CCR9−), small intestine (CD19+CLA−Integrinβ7+CCR10−CCR9+), CCR9+ alone (CD19+CLA−CCR9+) and CCR10+ alone(CD19+ CLA−CCR10+) homing profile. Data are shown in Whisker plots with a median, 25 and 75% quartile, and 5–95% confidence interval. Repeated measures ANOVA was performed *p < 0.05; **p < 0.01; ***p < 0.001
The total number of IgA+ and IgA− memory B cells with distinct homing profiles on day 18 was higher compared to the number of CTB-specific memory B cells. This can be explained by the fact that the response to CTB represents only part of the total response to the vaccine that also contains formalin- and heat-killed Vibrio cholerae bacteria.
In conclusion, the homing potential of memory B cells toward the colon and small intestine increased upon oral vaccination with Dukoral®, with a distinct preferential increase of IgA+ memory B cells expressing a colon homing profile and of IgA− memory B cells expressing a small intestine homing profile.
Oral vaccination induces homing to airways with different kinetics for IgA + and IgA − memory B cells
As oral vaccines are in contact with the oral mucosa and tonsils in the Waldeyer’s ring, it was evaluated whether oral cholera vaccination also induced homing of IgA+ and IgA− memory B cells to the airway mucosa. Oral vaccination induced an increased airway homing potential of peripheral IgA+ memory B cells, (CD19+CLA−Integrinβ1+β7−CCR10+) already at day 14, with a peak at day 18 (Fig. 4). In IgA− memory B cells, the strongest increase of cells with airway homing potential was observed on day 14.

Kinetics of IgA+ and IgA− memory B cells with an airways homing potential. PBMCs of healthy volunteers (n = 14) were analyzed on days 0, 14, 18, and day 28, using flow cytometry. Gating strategy (a) and kinetics (b) of IgA+ and IgA− memory B cells with homing potential to airways (CD19+CLA−Integrinβ1+β7-CCR10+). Data are shown in Whisker plots with a median, 25 and 75% quartile and 5–95% confidence interval. Repeated measures ANOVA was performed *p < 0.05; **p < 0.01; ***p < 0.001
Distribution of homing potential of lymphocytes at days 0 and 18
Additionally, we analyzed the effect of the cholera vaccine on distribution of homing potential to the periphery (lymph nodes (LN) and spleen, CLA− IgA+/IgG+CCR7+) (Supplementary Figure S3) and skin (CLA+IgA+/−Integrin β1+ integrin β7−CCR10+) (Supplementary Figure S4). Homing potential of memory B cells to the LN and spleen was significantly increased at day 18 for IgA+ memory B cells and at day 28 for both IgA+ and IgG+ memory B cells. The number of IgA− memory B cells with skin homing potential did not significantly change over time (Supplementary Figure S4). This was in contrast to IgA+ memory B cells, which had a significantly increased skin homing potential from day 14 onwards.
Both the number of CTB-specific memory B cells and specific homing profiles peaked at day 18. For each homing profile, the number of memory B cells was plotted at days 0 and 18 as relative percentage (r%) to compare the relative contribution per homing profile (Fig. 5). The total percentage (t%) of IgA+ memory B cells with a distinct homing profile at day 0 was 1.77 t%, while at day 18 this increased to 2.17 t%, resulting in a 1.22-fold increase (Fig. 5a). IgA− memory B cells had a lower percentage of cells with a homing potential at day 0 (1.53 t%) while this was 2-fold increased at day 18 to 3.06 t%. The majority of the memory B cells had a homing potential toward the periphery at both days 0 and 18 (84–99 r%) (Fig. 5a, b). In the mucosal tissues (small intestine, colon, and airways), the relative number of IgA+ memory B cells with homing potential toward these organs were increased 3.8-fold (Fig. 5c). The largest relative increase of IgA+ memory B cells was found for homing potential toward airways (from 36 to 50 r%), followed by colon (from 38 to 40 r%), whereas small intestine showed a decrease (from 26 to 10 r%). As the total frequency of IgA+ memory B cells was increased, the number of IgA+ memory B cells with a homing potential toward small intestine, colon, and airways per million of memory B cells was calculated for days 0 and 18. Indeed, Fig. 5e shows that there was a large increase in number of IgA+ memory B cells with homing potential toward airways (5.3-fold), followed by colon (4.0-fold), and skin (3.5-fold), whereas a small increase was observed for small-intestine-homing potential (1.5-fold) and peripheral homing (1.19-fold).

Distribution of homing potential of IgA+ and IgA− memory B cells at days 0 and 18. In the piecharts (a–d) relative percentages (r%) of homing potential of periphery, small intestine, colon, airways, and skin (together 100%) are depicted for IgA+ memory B cells (a) and IgA− memory B cells (b). c, d show the distribution in relative percentage (r%) of homing potential toward small intestine, colon, and airways only (together 100%) at day 0 vs. day 18. The increase in total number of cells at day 18 is shown as ×1.22–3.84 B cells under the piechart. In e and f, the number of cells with a homing potential toward small intestine, colon, airways, periphery and skin is given (#homing potential cells/106 memory B cells) at days 0 and 18
Remarkably, IgA− memory B cells showed a completely different profile. Only 10 r% of the IgA− memory B cells had an airway homing potential both at days 0 and 18 (Fig. 5d). Colon homing potential decreased from 43 r to 30 r%. Small-intestine-homing potential strongly increased to 60 r% at day 18 compared to 47 r% at day 0. Of all IgA− memory B cells expressing homing receptors, 90 r% had a gut homing potential on days 0 and 18, while this was only around 60 r% for IgA+ memory B cells. At day 18 after vaccination, the number of IgA− memory B cells per million of memory B cells with specific homing potential for mucosal sites increased 2-fold for the small intestine, 1.7-fold for the airway, 1.6-fold for the skin, and 1.2-fold for the colon (Fig. 5f).
Oral cholera vaccination does not increase vaccine-specific T-cell proliferation
To investigate whether vaccine-specific T-cell responses were increased in response to oral cholera vaccination PBMCs were stimulated for 7 days in the presence of the vaccine antigens CTB and Dukoral®, control antigen ovalbumin (OVA) or recall antigen tetanus toxoid (TT). A recall antigen is used as control to elicit a specific immune response in previously (Tetanus) vaccinated subjects. After 7 days, T-cell proliferation was measured by staining the cells for expression of the proliferation marker Ki67 (Fig. 6). After stimulation with CTB no increased T-cell proliferation was observed compared to medium. When Dukoral® and TT-induced proliferation was compared to medium controls, there was a highly significant difference at all time points (p < 0.001). This was already the case at day 0, also for Dukoral®, which is caused by cross reactivity as the patients were not vaccinated yet. When Dukoral-specific proliferation was followed over time and compared to Dukoral stimulation at day 0, no significant differences between the time points were observed, albeit day 14 seems to have increased T-cell proliferation (p = 0.2).

Antigen-specific T-cell proliferation. PBMCs of healthy volunteers (n = 14) were collected on days 0 and 14 before receiving Dukoral vaccination, and at day 28. PBMCs were stimulated for 7 days with a cholera toxin B (10 μg/ml) or Dukoral (108 bacteria/ml) or b IMDM medium, ovalbumin (OVA) (10 μg/ml) or tetanus toxoid (TT) (5 lf/ml). Proliferating T cells (CD3+ Ki67+) were analyzed by flow cytometry. Data are shown as mean with SEM. Repeated measures ANOVA was performed per stimulation, no significant differences found. For the comparisons between stimulations and medium control per day a univariate ANOVA (GLM) was used, *p < 0.05; **p < 0.01; ***p < 0.001
Additionally no significant increase in cytokine production (IFN-y, IL-5, IL-13, IL-10, and IL-17) was measured in vaccine antigen stimulated T cells compared to the medium and OVA controls (data not shown). Thus, we conclude that oral cholera vaccination had no vaccine-specific effects on T-cell proliferation and cytokine production at the time points analyzed.
Discussion
To our knowledge, this is the first study that documents the kinetics and detailed tissue homing preference of memory B cells following oral cholera vaccination. Oral vaccination induced Dukoral®- and CTB-specific IgG and IgA responses in serum, and enhanced the number of circulating memory B cells expressing marker profiles associated with homing to small intestine, colon, airways, skin, and periphery. The number of circulating CTB-specific memory B cells as well as memory B cells with distinct tissue homing profiles peaked on day 18. Interestingly, IgA+ memory B cells preferentially expressed markers that enable homing to the airways and to the colon, while IgA− (mainly IgG+) memory B cells primarily expressed markers associated with homing to the small intestine.
The amounts of Dukoral- and CTB-specific serum IgA and IgG increased until day 28 post vaccination. Other studies with this vaccine reported an earlier peak in the vaccine-specific antibody responses (days 18–21).7,23,24 This is not caused by a difference in age range or gender ratio of the study groups.25,26
In contrast to serum IgG and IgA levels, the number of circulating CTB-specific memory B cells in blood peaked at day 18, which is 4 days after the second oral vaccination, and decreased at day 28. This decrease in circulating cholera-specific memory B cells at day 28 may indicate that these cells have entered peripheral mucosal tissues, whilst still contributing to the serum IgA and IgG levels by producing these antibodies locally.
Rudin et al.7 and Leach et al.24 observed similar kinetics of vaccine-specific antibody producing B cells in peripheral blood, but Rahman et al.27 observed a peak at day 7 that was absent at day 21. While the first two studies were performed in Sweden, the study by Rahman et al. was performed in Bangladesh. As cholera is endemic in Bangladesh but not in Europe, the kinetics in the study by Rahman might thus reflect a memory response rather than a primary response, explaining the different kinetics seen between these studies.
In nasal washes, saliva and fecal extracts, the amount of vaccine-specific IgA antibody was low and no significant increase was detected after vaccination. The levels of vaccine-specific IgG in nasal washes were low but significantly increased at day 28. The finding that larger amounts of vaccine-specific IgG were measured in the serum and nasal washes than vaccine-specific IgA is in line with previous reports.7,23,24,28 We speculate that this might be due to uptake of the oral vaccine in the lymphoid tissue of the palatine tonsils, resulting in activation of larger numbers of IgG plasma cells (55–72%) than IgA plasma cells (13–18%), which will preferentially home toward the respiratory tract.29 This will lead to more vaccine-specific IgG in the nasal cavity compared to vaccine-specific IgA. Besides, it is known that the nasal mucosa harbors the largest number of IgG plasma cells compared to other mucosal tissues, even though in steady state more IgA plasma cells are present locally compared to IgG plasma cells.14
In addition to studying the effect of oral vaccination on vaccine-specific memory B cells and antibody production, we investigated whether oral vaccination would also affect tissue homing potential of B cells. Tissue homing-related molecules such as selectins, integrins, chemokines and chemokine receptors can be expressed on several different cell types as well as in different tissues. For this reason a combination of several homing markers rather than single markers is needed to define distinct tissue homing properties. In this study we focussed on integrin β1, integrin β7, CCR9, CCR10, and CLA to define homing to gut, airways, skin, small intestine, and colon.15,16,17,18,30,31,32,33,34,35,36
CCR10 can bind to CCL28 that is expressed in the respiratory tract together with VCAM-134,35, but also in the colon, tonsils, and lactating mammary glands.14,16 In addition, CCR10 can also bind to CCL27 produced in the skin.17,33,37,38 Homing to the gastrointestinal tract can be specified to colon vs. small intestine on the basis of expression of CCR10 and CCR9. The ligand of CCR9, CCL25 is expressed both in the small intestine as well as in the thymus.39 Therefore, expression of CCR10 and CCR9 cannot be used to define tissue homing preference of lymphocytes without studying additional markers. For differentiating between markers related to homing to the skin, small intestine, colon, and airways, expression of Integrin β1 and integrin β7, and CLA were also measured. Integrin β1 and integrin β7 represent the expression of the heterodimer integrin α4β140,41 and α4β720,42 leading respectively to homing potential to non-intestinal (α4β7−, α4β1+) and intestinal tissues (α4β7+). The homing profiles of B cells that can home to airways and skin are quite comparable, and are based on expression of CCR10 in combination with α4β1 without α4β7. They are differentiated by expression of the skin homing marker CLA. CLA can bind E-selectin, which is expressed in the skin, as wells as on oral mucosa and inflamed tissues.18,31,32 However, even in the presence of CLA, homing to airways cannot be excluded. As recently shown by Seong et al, CLA+ plasmablasts are able to migrate into the oral cavity under inflammatory conditions, when E-selectin expression is upregulated. Furthermore, patients suffering from upper respiratory tract infections show a strong increase in CLA and CD62L expression.19 As the present study was performed in healthy donors we have made the assumption that CLA expression in this study does not reflect homing to the oral cavity.
To date, no markers are known for B cells to distinguish between homing to the lower and upper respiratory tract, even though CCR3 has been described as a marker for differential T-cell homing to the nasal mucosa.36 In addition to the markers we used, the use of additional markers like CCR3 and GPR1519,36 in even more extensive homing panels will help to further specifying tissue homing potential of circulating lymphocytes in future studies.
The tissue homing potential of the memory B-cell populations was characterized using the homing marker combinations designed for airways, small intestine, colon, periphery, and skin (Supplementary Figure S1). Cholera vaccination enhanced the number of circulating memory B cells for all tissue homing marker profiles with kinetics following the kinetics as seen in the CTB-specific memory B cells, suggesting a vaccination induced expression of distinct homing profiles.
Interestingly, IgA+ memory B cells preferentially expressed a homing marker profile associated with homing to the airways (CLA−Integrinβ1+ Integrinβ7− CCR10+), colon (CLA−Integrinβ7+ CCR10+CCR9−), and even to the skin (CLA+ Integrinβ1+ Integrinβ7− CCR10+) (Fig. 5). All of these homing profiles are linked to expression of CCR10. In contrast to IgA+ memory B cells, IgA−memory B cells showed homing marker profiles consistent with homing toward the small intestine (CLA−Integrin β7+ CCR10−CCR9+; Fig. 5). It should be noted however, that this conclusion is based on the expression of tissue homing markers on peripheral blood lymphocytes, and do not formally demonstrate homing of these cells to these peripheral tissues. As stromal factors, such as retinoic acid,43 determine tissue homing properties of locally activated lymphocytes, it has been suggested that antigens taken up in the tonsils of Waldeyer’s ring will induce the formation of especially IgG-antibody producing memory B cells with homing capacity to lungs, upper airways, and peripheral blood.29 In contrast, IgA+ B cells induced in the Peyer’s patches are thought to preferentially home to the intestine, specifically the small intestine.14,44 This hypothesis is in line with the antibody production detected in nasal wash (mainly IgG) and fecal water (mainly IgA). However, this hypothesis does not explain the observation of IgA+ memory B cells homing preferences to airways or colon, and of IgA− memory B cells homing to the small intestine, respectively. Although we have no evidence on where the B cells were activated after vaccination, a possible explanation could be that the IgA− memory B cells we detect in blood were induced in the PP and are instructed to home to the small intestine under the influence of stromal factors, and that the IgA+ memory B cells that can home to the airways or colon were induced elsewhere, either in Waldeyer’s ring or in the isolated follicles of the colon. If the vaccine induces a proinflammatory response in PP, it may induce IgG there, whereas a more regulatory-biased IgA response to the vaccine is induced in Waldeyers ring or colon.
CD19 is maintained on peripheral B cells, including memory B cells, but also on plasmablasts and subsets of plasma cells. Based on CD19 expression only, memory B-cells will also include a small number of CD19+ plasmablasts.45 Subsets of plasmablasts and plasma cells are further defined by the level of expression of CD38 and CD138. The CD38hi subset of peripheral B-cells comprises both newly generated plasmablasts and plasma cells that recirculate from the bone marrow.
The B cells in the present study are primarily CD19+ memory B cells, as shown in Supplementary Figure S2. The data are thus complementary to the homing potential of CD19- plasmablasts that were described recently.19 Our data on memory B cells also differs from earlier work in relation to the kinetics of CTB-specific plasmablasts.46 In this study CTB-specific spot forming cells were studied after oral vaccination against cholera. The number of CTB-specific cells peaked at days 7 and 21, and were almost at background levels at days 14 and 28. This is in contrast to our results where we have a peak at a similar moment, but at days 14 and 28 the number CTB-specific IgA and IgG memory B cells is clearly increased compared to day 0. This would imply that CTB-specific memory B cells remain in circulation for a prolonged period compared to plasma cells. Likewise, the expression of homing marker integrin β7 also seems to differ between memory B cells. After oral vaccination with Salmonelly typhi Ty21a 100% of the IgA, IgG, and IgM plasmablasts express α4β7.20 This is very much in contrast with our findings after cholera vaccination, where only 2.5% (% of all memory B cells) of the IgA+ memory B cells are positive for β7 expression and 65% (% of all memory B cells) of the IgA− memory B cells expressed β7 after oral cholera vaccination. This difference is not caused by β7 expression alone, as β7 expression was compared to α4β7 expression in this study and all the differences in β7 expression was due to α4β7 expression.20 A possible explanation is the differences in the vaccine used. Salmonella typhi Ty21a may be a more potent inducer of gut homing than the cholera vaccine.
In addition to the paper of Kantele et al. we measured CCR9 and CCR10 in combination of α4β7 to be able to distinguish potential homing towards the colon and small intestines for IgA+ and IgA− memory B cells. That the combination of the markers β7, CCR10, and CCR9 is able to distinguish between colon and small intestine homing potential is confirmed by another study.47 In this study both IgA+ and IgA− plasmablasts in healthy donors had a higher level of CCR9 expression compared to CCR10 expression. However, during severe Ulcerative Colitis CCR10 is upregulated in the α4β7+ IgA+ and IgA− PBs. This is in constrast with our finding that upon oral cholera vaccination IgA+ memory B cells have a higher α4β7 CCR10+ (colon) expression and IgA− memory B cells have a α4β7 CCR9+ (small intestine) expression.
Despite consistent induction of humoral immune responses in blood and mucosal fluids, we were not able to detect significant increases in vaccine-specific T-cell proliferation and cytokine production after oral vaccination as reported by others.48 However, one study reported the induction of T-cell proliferation following oral cholera vaccination.49 One possible explanation is that the T-cell proliferation takes place in the draining lymph nodes, and that the recirculation of the T cells in peripheral blood peaks at different time points than the selected study days (days 0, 14, and 28). As tissue homing of non-B cells (mainly T cells) followed comparable kinetics compared to B cells (data not shown), the peak of recirculating vaccine-specific T cells may have been at day 18 rather than at day 14 or 28.
Taken together, this study demonstrates that oral cholera vaccination not only induces humoral immunity in the gastrointestinal tract, but also in other mucosal locations, such as the respiratory tract. Therefore, oral vaccination could be considered as an option for vaccination against respiratory pathogens. This may be an obvious route of vaccination, as food components such as 1,25-dihydroxyvitamin D3 can induce expression of CCR10 in human B cells and thereby direct B cells to mucosal sites, including lungs.38 Furthermore, retinoic acid (RA), a vitamin A metabolite, is known to induce immune cells to express CCR9, thereby promoting homing to the small intestine.21,22 The immunomodulating properties of vitamin A and D could be part of the overall gut–lung axis framework.50 In this ‘gut–lung axis’ theory, the interplay between diet, gut microbiota, and improved respiratory health by migrating immune cells is linked together, although the exact mechanisms are not yet fully understood.51,52,53,54 Oral cholera vaccination may therefore also be a useful model to study the immunomodulatory capacity of (food) components.
Materials and methods
Volunteers, specimens, and sample collection
Healthy adult volunteers (total n = 14, range 20–46 years, 25.07 ± 6.53 years, 10 females, 4 males) gave informed consent to participate in this study. The study was approved by the Research Ethics Committee of Wageningen University and registered at www.clinicaltrials.gov with identification-number: NCT02238548. Volunteers, who had not previously been vaccinated with Vibrio Cholera or E. coli, were vaccinated with the oral cholera vaccine Dukoral® (EU/1/03/263/002, Crucell, Leiden, The Netherlands). The vaccine consisted of a total of 1.25 × 1011 formalin- and heat-killed Vibrio cholerae bacteria (strains O1 Inaba and O1 Ogawa) and 1 mg recombinant cholera toxin B-subunit (rCTB). Each individual received Dukoral® in the sodium carbonate buffer supplied with the vaccine.
Blood, nasal washes, saliva, and fecal samples were collected at baseline on study day 0, and at days 7, 14, 18, and 28 after the first vaccination dose. On the vaccination days (days 0 and 14) biological samples were collected before the vaccine was given. Blood was collected in K2-EDTA tubes (367525 BD, Franklin Lakes, NJ, USA) for PBMC isolation and in serum tubes (BD 367895, Plymouth, United Kingdom) for serum collection. Serum was stored at −80 °C. PBMCs were isolated within 24 h using 50 ml Leucosep tubes (227290, Greiner Bio-One, Alphen a/d Rijn, The Netherlands) filled with Ficoll plaque plus (17–1440–02, GE Healthcare Life Sciences, Uppsala, Sweden) according to the manufacturer’s protocol. Remaining PBMCs were cryopreserved and stored in liquid nitrogen.
For collection of nasal wash, volunteers were instructed not to use any nasal sprays from the evening before until the collection. For the nasal wash 3 ml pre-warmed saline (32 °C) was injected in each nostril and collected into a container. Containers were put on ice and processed within 2 h after collection. Nasal wash samples were centrifuged at 3000×g for 10 min at 4 °C. Supernatant was stored at −80 °C.
For saliva collection, volunteers were not allowed to ingest anything else than water for 1 h preceding sampling. Ten minutes after rinsing the mouth with water, saliva was collected by passive drooling for 6 min. Saliva was collected, vortexed, and stored at −80 °C.
Fecal samples were collected using a fecotainer (AT Medical BV, Enschede, The Netherlands) and stored at home at −20 °C and transported on dry ice to the research facility at the end of the study. Fecal extracts were prepared by adding 3.6 ml of PBS and 40 μl Protease inhibitor (Sigma-Aldrich, Zwijndrecht, The Netherlands P8340) to 0.4 g of feces, vortexing at least 30 s, and centrifuged at 3000×g for 10 min at 4 °C. Fecal extracts were stored at −80 °C until measurement.
T-cell proliferation
To assess antigen-specific T-cell proliferation (day 0, day 14, day 28), PBMCs were freshly isolated from blood, plated in a 24 wells culture plate (Costar 3526, Sigma-Aldrich, Zwijndrecht, The Netherlands; 1 × 106 PBMCs/well) and stimulated with different stimuli for 7 days in IMDM medium (Gibco 31980–022, Grand Island, New York, USA) supplemented with 5% Human AB serum, 1% Pen/Strep, 0,2% Normocin (#ant-nr-1 Invivogen, San Diego, California, USA). The stimuli were CTB 10 μg/ml (Sigma-Aldrich, C9903, Zwijndrecht, The Netherlands), TT 5 lf/ml (Statens Serum Institut, Copenhagen, Denmark, kindly provided by W. van de Veen, SIAF Switzerland), OVA 10 μg/ml (Invivogen 9006–59–1, Toulouse, France) and Dukoral® vaccine 108 bacteria/mland 0,8 μg CTB/ml. After 7 days, supernatants were collected and stored at −80 °C for cytokine analysis using BD™ Cytometric Bead Array (CBA) Human Th1/Th2/Th17 Kit, (BD Bioscience 560484, Franklin Lakes, New Jersey, USA). Cells were collected and analyzed on FACS CantoII for proliferation by staining cells for the following markers: anti-CD3-Pe-Cy5 (BD Pharmingen 555334, Franklin Lakes, New Jersey, USA), anti-CD19 BV510 (BD Horizon 562947, Franklin Lakes, New Jersey, USA), anti-Ki67-PE or Ki67 isotype control (BD Pharmingen 556027, Franklin Lakes, New Jersey, USA) and fixable viability dye V450 (Ebioscience 65–0863–18, Waltham, Massachusetts, USA).
Flow cytometric analysis of peripheral B cells
Freshly isolated PBMCs were stained with antibody panels and/or fluorescently labeled CTB to detect CTB-specific memory B cells, and analyzed by using multi-color flow cytometry. The selection of tissue specific homing marker profiles was done based on literature research and three antibody panels were composed (see Table 1). For every panel, 3 × 106 freshly isolated PBMCs were stained in a 96-well plate (NUNC PP Sigma-Aldrich 7116, Zwijndrecht, The Netherlands) for 30 min at 4 °C, washed with cold FACS buffer (PBS, 0,5% BSA, 3% FCS, 2 mM EDTA, 0,05% NaN3) and centrifuged at 400×g for 3 min at 4 °C. Cells were analyzed at medium flow for 180 s using a FACS CantoII. Due to technical issues, the FACS measurement had to be repeated for all day 0 samples, using defrosted PBMCs that had been stored in liquid nitrogen. The effect of freeze/thawing was examined on five donors at time point day 14 and most markers were slightly increased or equal after thawing (Integrin Beta 7, Integrin Beta 1, CCR10, and CLA), while only CCR9 is slightly lower expressed (data not shown). However, this did not compromise the results, as most of these markers at day 0 resulted in lower number of cells with these homing profile compared to other time points and for CCR9 even higher numbers at day 0 compared to day 14. To exclude dead cells in these samples (day 0), specific B cells were stained with fixable viability dye V450 in PBS for 30 min. For the gut, airways and skin homing panel 3, DRAQ 7 (70250, Biostatus, Leicestershire, United Kingdom) was added as live/dead marker to exclude dead cells (day 0 only). The viability of cells was measured for specific B cells (panel 1) at all time points using fixable viability dye V450 and resulted in 100% alive cells as expected for fresh cells. This in combination with strict gating on preferably mutually exclusive markers as CD3 and CD19 should result in reliable data for all panels at all time points. For all panels fluorescent minus one (FMO) controls and isotype controls were included. This was done for CTB (only FMO), CCR7, CLA, CCR10, and CCR9.
Flow cytometry data analysis was performed by using FlowJo software (version 10 TreeStar, Inc.) and gating was performed as is shown in Supplementary Figure S1. Data were exported as % of memory B cells and subsequently converted to ‘number of homing memory B cells/million of memory B cells’.
ELISA
CTB- and Dukoral®- specific IgA and IgG antibodies in serum, saliva, nasal wash, and fecal extract were determined by enzyme-linked immunosorbent assay (ELISA) in high binding 96-well plates (Greiner Bio one 655061, Monroe, North Carolina, USA) and performed as described previously.55 In short, plates were coated with either 0.5 μg/ml CTB or 1 × 108 bacteria + 0.8 μg/ml CTB from Dukoral®/ml PBS (Lonza Be17–516F, Basel, Switzerland) and incubated overnight at 4 °C. After washing with wash buffer (PBS + 0.05% Tween-20), plates were blocked with ELISA blocking buffer (Roche 11112589001, Mannheim, Germany) for 1 h. For specific-IgA and IgG measurements serum was diluted in ELISA blocking buffer. Before diluting the samples nasal wash and saliva were centrifuged for 15 min at 1500×g and fecal extracts for 2 min at 1000×g. Nasal wash, saliva, and fecal extracts were incubated for 15 min in High Performance Elisa (HPE) buffer (Sanquin M1940, Amsterdam, The Netherlands). Samples were added to the plate in duplicate and incubated for 2 h. A pooled serum sample, from all vaccinated donors at day 18, was used to make a reference calibration line in all ELISAs and is expressed in arbitrary units/ml (AU/ml). After sample incubation, detection antibodies goat-anti-human IgA or IgG-horseradish peroxidase (HRP) (Southern Biotech 2050–05, 2040–05, Birmingham, Alabama, USA) diluted in Elisa blocking buffer (serum) or HPE buffer (saliva, nasal wash and fecal extracts samples) were incubated for 1 h. For development of all ELISAs TMB (SDT, Baesweiler, Germany) was added and the reaction was stopped by adding 2% HCL solution. Plates were read in a Filtermax at 450 nm minus 620 nm as reference value.
Statistics
Data were 10 log-transformed (ELISA) or logistically transformed (FACS) to obtain normally distributed data. A one-way repeated measures ANOVA was conducted using transformed data with IBM SPSS Statistics 23. Differences were considered significantly different when p < 0.05. First, the assumption of sphericity was tested, using Mauchly’s Test of Sphericity. If p < 0.05, sphericity could not be assumed and hence, a Greenhouse–Geisser correction was applied to test within-subject effects (time). Only when a significant within subject effect was observed (time), a pairwise comparison between timepoints with a Bonferroni correction was made.
References
Neutra, M. R. & Kozlowski, P. A. Mucosal vaccines: the promise and the challenge. Nat. Rev. Immunol. 6, 148–158 (2006).
Belyakov, I. M. & Ahlers, J. D. Comment on “Trafficking lymphocytes to mucosal surfaces following intramuscular vaccination. J. Immunol. 182, 1779 (2009).
Holmgren, J. & Svennerholm, A.-M. Vaccines against mucosal infections. Curr. Opin. Immunol. 24, 343–353 (2012).
Srivastava, A., Gowda, D. V., Madhunapantula, S. V., Shinde, C. G. & Iyer, M. Mucosal vaccines: a paradigm shift in the development of mucosal adjuvants and delivery vehicles. APMIS 123, 275–288 (2015).
Jia, Y., Krishnan, L. & Omri, A. Nasal and pulmonary vaccine delivery using particulate carriers. Expert Opin. Drug Deliv. 12, 993–1008 (2015).
Quiding, M. et al. Intestinal immune responses in humans. Oral cholera vaccination induces strong intestinal antibody responses and interferon-gamma production and evokes local immunological memory. J. Clin. Invest. 88, 143–148 (1991).
Rudin, A., Johansson, E. L., Bergquist, C. & Holmgren, J. Differential kinetics and distribution of antibodies in serum and nasal and vaginal secretions after nasal and oral vaccination of humans. Infect. Immun. 66, 3390–3396 (1998).
Rudin, A., Riise, G. C. & Holmgren, J. Antibody responses in the lower respiratory tract and male urogenital tract in humans after nasal and oral vaccination with cholera toxin B subunit. Infect. Immun. 67, 2884–2890 (1999).
Czerkinsky, C. & Holmgren, J. Mucosal delivery routes for optimal immunization: targeting immunity to the right tissues. Curr. Top. Microbiol Immunol. 354, 1–18 (2012).
Kozlowski, P. A., Cu-Uvin, S., Neutra, M. R. & Flanigan, T. P. Comparison of the oral, rectal, and vaginal immunization routes for induction of antibodies in rectal and genital tract secretions of women. Infect. Immun. 65, 1387–1394 (1997).
Eriksson, K. et al. Specific-antibody-secreting cells in the rectums and genital tracts of nonhuman primates following vaccination. Infect. Immun. 66, 5889–5896 (1998).
Kutteh, W. H., Kantele, A., Moldoveanu, Z., Crowley-Nowick, Pa & Mestecky, J. Induction of specific immune responses in the genital tract of women after oral or rectal immunization and rectal boosting with Salmonella typhi Ty 21a vaccine. J. Reprod. Immunol. 52, 61–75 (2001).
Chi, J.-T. et al. Endothelial cell diversity revealed by global expression profiling. Proc. Natl Acad. Sci. USA 100, 10623–10628 (2003).
Brandtzaeg, P. & Johansen, F. Mucosal B cells: phenotypic characteristics, transcriptional regulation, and homing properties. Immunol. Rev. 206, 32–63 (2005).
Zabel, B. A. et al. Human G protein-coupled receptor GPR-9-6/CC chemokine receptor 9 is selectively expressed on intestinal homing T lymphocytes, mucosal lymphocytes, and thymocytes and is required for thymus-expressed chemokine-mediated chemotaxis. J. Exp. Med. 190, 1241–1256 (1999).
Johansen, F. et al. Regional induction of adhesion molecules and chemokine receptors explains disparate homing of human B cells to systemic and mucosal effector sites: dispersion from tonsils. Blood 106, 593–600 (2005).
Pan, J. et al. Cutting edge: a novel chemokine ligand for CCR10 And CCR3 expressed by epithelial cells in mucosal tissues. J. Immunol. 165, 2943–2949 (2000).
Andrian, U. Hvon & Mackay, C. R. T-cell function and migration. Two sides of the same coin. N. Engl. J. Med. 343, 1020–1034 (2000).
Seong, Y., Lazarus, N. H., Sutherland, L., Greenberg, H. B. & Butcher, E. C. Trafficking receptor signatures define blood plasmablasts responding to tissue-specific immune challenge. JCI Insight 2, e90233 (2017).
Kantele, A. et al. Homing potentials of circulating lymphocytes in humans depend on the site of activation: oral, but not parenteral, typhoid vaccination induces circulating antibody-secreting cells that all bear homing receptors directing them to the gut. J. Immunol. 158, 574–579 (1997).
Bakdash, G., Vogelpoel, L. T. C., Capel van, T. M., Kapsenberg, M. L. & de Jong, E. C. Retinoic acid primes human dendritic cells to induce gut-homing, IL-10-producing regulatory T cells. Mucosal Immunol. 8, 265–278 (2015).
Hammerschmidt, S. I. et al. Retinoic acid induces homing of protective T and B cells to the gut after subcutaneous immunization in mice. J. Clin. Invest. 121, 3051–3061 (2011).
Alam, M. M. et al. Antigen-specific memory B-cell responses in Bangladeshi adults after one- or two-dose oral killed cholera vaccination and comparison with responses in patients with naturally acquired cholera. Clin. Vaccin. Immunol. 18, 844–850 (2011).
Leach, S., Lundgren, A. & Svennerholm, A. M. Different kinetics of circulating antibody-secreting cell responses after primary and booster oral immunizations: a tool for assessing immunological memory. Vaccine 31, 3035–3038 (2013).
Klein, S. L., Marriott, I. & Fish, E. N. Sex-based differences in immune function and responses to vaccination. Trans. R. Soc. Trop. Med. Hyg. 109, 9–15 (2015).
Cook, I. F. Sexual dimorphism of humoral immunity with human vaccines. Vaccine 26, 3551–3555 (2008).
Rahman, A. et al. Antibody-secreting cell responses after Vibrio cholerae O1 infection and oral cholera vaccination in adults in Bangladesh. Clin. Vaccin. Immunol. 20, 1592–1598 (2013).
Akhtar, M. et al. Kinetics of antibody-secreting cell and fecal IgA responses after oral cholera vaccination in different age groups in a cholera endemic country. Vaccine 35, 321–328 (2017).
Brandtzaeg, P. Potential of nasopharynx-associated lymphoid tissue for vaccine responses in the airways. Am. J. Respir. Crit. Care Med. 183, 1595–1604 (2011).
Kunkel, E. J. & Butcher, E. C. Chemokines and the tissue-specific migration of lymphocytes. Immunity 16, 1–4 (2002).
Berg, B. E. L. et al. The cutaneous lymphocyte antigen is a skin lymphocyte homing receptor for the vascular lectin endothelial cell-leukocyte adhesion molecule 1. J. Exp. Med. 174, 1461–1466 (1991).
Hirahara, K. et al. The majority of human peripheral blood CD4+CD23high Foxp3+regulatory T cells bear functional skin-homing receptors. J. Immunol. 177, 4488–4494 (2006).
Morales, J. et al. CTACK, a skin-associated chemokine that preferentially attracts skin-homing memory T cells. Proc. Natl Acad. Sci. USA 96, 14470–14475 (1999).
Bentley, A. et al. Expression of endothelial and leukocyte adhesion molecules intercellular adhesion molecule-1, E-selectin, and vascular cell adhesion molecule-1 in the bronchial mucosa in steady-state and allergen-induced asthma. J. Allergy Clin. Immunol. 92, 857–868 (1993).
Uhlén, M. et al. Tissue-based map of the human proteome. Science 347, 1260419 (2015).
Danilova, E. et al. A role for CCL28-CCR3 in T-cell homing to the human upper airway mucosa. Mucosal Immunol. 8, 107–114 (2015).
Sigmundsdottir, H. et al. DCs metabolize sunlight-induced vitamin D3 to ‘ program’ T cell attraction to the epidermal chemokine CCL27. Nat. Immunol. 8, 285–293 (2007).
Shirakawa, A. et al. 1,25-dihydroxyvitamin D3 induces CCR10 expression in terminally differentiating human B cells. J. Immunol. 180, 2786–2795 (2008).
Kunkel, E. J. et al. Lymphocyte CC chemokine receptor 9 and epithelial thymus-expressed chemokine (TECK) expression distinguish the small intestinal immune compartment: epithelial expression of tissue-specific chemokines as an organizing principle in regional immunity. J. Exp. Med. 192, 761–768 (2000).
Dorner, M. et al. Beta1 integrin expression increases susceptibility of memory B cells to Epstein-Barr virus infection. J. Virol. 84, 6667–6677 (2010).
Kamel, A. M. et al. Adhesion molecules expression in CLL: potential impact on clinical and hematological parameters. J. Egypt Natl Canc. Inst. 28, 31–37 (2016).
Rott, L. S., Briskin, M. J. & Butcher, E. C. Expression of alpha4beta7 and E-selectin ligand by circulating memory B cells: implications for targeted trafficking to mucosal and systemic sites. J. Leukoc. Biol. 68, 807–814 (2000).
Hammerschmidt, S. I. et al. Stromal mesenteric lymph node cells are essential for the generation of gut-homing T cells in vivo. J. Exp. Med. 205, 2483–2490 (2008).
Baaten, B. J. G., Cooper, A. M., Swain, S. L. & Bradley, L. M. Location, location, location: the impact of migratory heterogeneity on T cell function. Front Immunol. 4, 1–11 (2013).
Engel, P. et al. Abnormal B-lymphocyte development, activation, and differentiation in mice that lack or overexpress the Cd19 signal-transduction molecule. Immunity 3, 39–50 (1995).
Czerkinsky, C., Svennerholm, A. M., Quiding, M., Jonsson, R. & Holmgren, J. Antibody-producing cells in peripheral-blood and salivary-glands after oral cholera vaccination of humans. Infect. Immun. 59, 996–1001 (1991).
Tarlton, N. J. et al. Plasmablast frequency and trafficking receptor expression are altered in pediatric ulcerative colitis. Inflamm. Bowel Dis. 18, 2381–2391 (2012).
Arifuzzaman, M. et al. Antigen-specific memory T cell responses after vaccination with an oral killed cholera vaccine in Bangladeshi children and comparison to responses in patients with naturally acquired cholera. Clin. Vaccin. Immunol. 19, 1304–1311 (2012).
Ahmed, T., Arifuzzaman, M., Lebens, M., Qadri, F. & Lundgren, A. CD4+T-cell responses to an oral inactivated cholera vaccine in young children in a cholera endemic country and the enhancing effect of zinc supplementation. Vaccine 28, 422–429 (2010).
Marsland, B. J., Trompette, A. & Gollwitzer, E. S. The gut-lung axis in respiratory disease. Ann. Am. Thorac. Soc. 12, S150–S156 (2015).
Marsland, B. J. & Gollwitzer, E. S. Host-microorganism interactions in lung diseases. Nat. Rev. Immunol. 14, 827–835 (2014).
Budden, K. F. et al. Emerging pathogenic links between microbiota and the gut–lung axis. Nat. Rev. Microbiol 15, 55–63 (2016).
Noval Rivas, M., Crother, T. R. & Arditi, M. The microbiome in asthma. Curr. Opin. Pediatr. 28, 764–771 (2016).
Samuelson, D. R., Welsh, D. A. & Shellito, J. E. Regulation of lung immunity and host defense by the intestinal microbiota. Front. Microbiol. 6, 1–14 (2015).
Bergquist, C. et al. Intranasal vaccination of humans with recombinant cholera toxin B subunit induces systemic and local antibody responses in the upper respiratory tract and the vagina. Infect. Immun. 65, 2676–2684 (1997).
Acknowledgements
We acknowledge all people that were part of the MOSAIC study team, in particular Daphne Montizaan and Olaf Perdijk for helping with the execution of the study and Alwine Kardinaal for helping with the statistical analysis. We also thank all participants of the MOSAICI study. This study was financed by FrieslandCampina.
Author information
Authors and Affiliations
Contributions
Study design: R.J.J.v.N., E.v.H., H.F.J.S., M.v.S., H.T., L.H.U., S.B., and J.M.W. Medical care during study execution: B.W. Execution and logistics of the study: E.G.F.-V., E.v.H., M.v.S., E.L.-v.d.B., and B.M. Data analysis: M.v.S., E.L.-v.d.B., and B.M. Manuscript writing and editing: M.v.S., S.B., H.F.J.S., E.v.H., and R.J.J.v.N.
Corresponding author
Ethics declarations
Conflict of interest
R.J.J.v.N. and L.H.U. are employed by FrieslandCampina. E.v.H., E.G.F.-V., H.T., E.L.-v.d.B., and M.v.S. received research funding from FrieslandCampina to execute the study. All the remaining authors declare no competing financial interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
About this article

Cite this article
van Splunter, M., van Hoffen, E., Floris-Vollenbroek, E.G. et al. Oral cholera vaccination promotes homing of IgA+ memory B cells to the large intestine and the respiratory tract. Mucosal Immunol 11, 1254–1264 (2018). https://doi.org/10.1038/s41385-018-0006-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41385-018-0006-7
This article is cited by
-
Recombinant L. lactis vaccine LL-plSAM-WAE targeting four virulence factors provides mucosal immunity against H. pylori infection
Microbial Cell Factories (2024)
-
Mucosal vaccines — fortifying the frontiers
Nature Reviews Immunology (2022)
