
Abstract
Tyrosine (Tyr) phosphorylation is implicated in the modification of several erythrocyte functions, such as metabolic pathways and membrane transport, as well as in signal transduction systems. Here we describe the map of Tyr-phosphorylated soluble proteins of newborn red blood cells (RBC) using an in vitro model simulating RBC reoxygenation at birth after an intrauterine hypoxic event. We tested the hypothesis that a hypoxic environment and subsequent reoxygenation promote post-translational changes in the RBC protein map of newborns, in addition to desferrioxamine (DFO)-chelatable iron (DCI) release and methemoglobin (MetHb) formation. Umbilical cord blood RBC were incubated under hypoxic conditions for 16 h at 37°C, and subsequently for 8 h under aerobic conditions. Control erythrocytes were incubated under aerobic conditions at 37°C for the period of the experiment, i.e. for 24 h. Tyr-phosphorylation proteins were assessed using advanced high-resolution two-dimensional electrophoresis, 2-D immunoblot analysis with anti-phosphotyrosine (anti-pTyr) antibodies, and computer-aided electrophoretogram analysis. Higher DCI release and MetHb formation were observed in newborn RBC incubated under hypoxic conditions than in those incubated aerobically. Different immunoreactivity patterns with anti-pTyr antibodies were also observed between newborn RBC incubated under hypoxic conditions and controls. A hypoxic environment is a factor promoting DCI release, a well-known condition of oxidative stress. This is the first map of Tyr-phosphorylated soluble proteins of newborn RBC obtained using an in vitro model simulating RBC reoxygenation at birth after an intrauterine hypoxic event. Our results suggest that hypoxia increases Tyr-phosphorylation of antioxidant proteins, protecting RBC against oxidative stress.
Similar content being viewed by others

Main
RBC are exposed to oxidative stress more than other cells of the body as a result of their high membrane polyunsaturated fatty acid content and high cellular concentrations of oxygen and heme iron. Iron plays a central role in generating harmful ROS. Its redox cycling promotes the Fenton reaction, which produces the potent oxidant hydroxyl radical (1). To be redox-cycling active, iron must be released from its macromolecular complexes (mainly transport and storage proteins) (2). We previously showed that iron is released from Hb in a DFO-chelatable form when erythrocytes are exposed to oxidative stress, such as when they are incubated with oxidizing agents or undergo prolonged aerobic incubation, two models of erythrocyte ageing (3). Iron release is accompanied by Met-Hb formation and oxidative damage to erythrocyte membrane proteins and lipids (3). In several studies, we used DCI in erythrocytes as an index of oxidative stress in various adult and neonatal pathophysiological conditions (4–7). We found that RBC of hypoxic newborns have a higher DCI content (6,7) and greater susceptibility to iron release after aerobic incubation than RBC of nonhypoxic newborns (5). In vitro experiments clearly showed that hypoxia was more significant than reoxygenation as a factor promoting iron release and MetHb formation in erythrocytes (8). During hypoxic incubation of partially oxygenated Hb, the superoxide radical is formed and autoxidation of Hb occurs (9). Hypoxia also induces glycolytic hyperactivity and accumulation of 2,3-DPG (10).
It was recently reported that Tyr phosphorylation is implicated in the modification of several erythrocyte functions, including metabolic pathways (11), membrane transport (12), and cell volume and shape (13). Phosphorylation of proteins is a common characteristic of signal transduction systems. Immunoblotting with anti-pTyr antibodies of proteins separated by monodimensional SDS-PAGE has been widely used to study intracellular signaling pathways activated by extracellular stimuli (14). High-resolution 2-D SDS-PAGE in conjunction with 2-D immunoblot analysis with anti-pTyr antibodies enables several hundred Tyr-phosphorylated proteins to be studied at the same time (15).
In the present study, we assessed Tyr-phosphorylation proteins in normal and hypoxic RBC from newborns using advanced 2-D electrophoresis and computer-aided electrophoretogram analysis. We tested the hypothesis that a hypoxic environment and subsequent reoxygenation promote post-translational changes in the RBC protein map of newborns, in addition to DCI release and MetHb formation. We used an in vitro model simulating RBC reoxygenation at birth after an intrauterine hypoxic event.
MATERIALS AND METHODS
Blood samples.
Heparinized blood samples were obtained from the umbilical vein of five healthy full-term newborns, immediately after cord clamping. Gestational age ranged from 37 to 41 wk and birth weight ranged from 2900 to 4200 g. Informed consent was obtained from the parents. The study was approved by the Human Ethics Committee of the Medical Faculty of the University of Siena.
Erythrocyte incubation.
After centrifuging at 600 × g for 10 min at room temperature, plasma and buffy coats were removed. The erythrocytes were washed three times with 0.123 M NaCl, 28 mM sodium phosphate/potassium phosphate buffer, pH 7.4, and resuspended in the same buffer at 50% (vol/vol) as previously reported (16). To simulate the situation occurring in neonates at birth after an intrauterine hypoxic event (17), we used a model of hypoxia-reoxygenation that consisted of incubating RBC without glucose under hypoxic conditions (exposed to a mixture of 4% O2, 6% CO2, and 90% N2) for 16 h at 37°C and subsequently for 8 h under aerobic conditions. Control erythrocytes were incubated under aerobic conditions at 37°C for the period of the experiment, i.e. for 24 h. All incubations were carried out in a shaker apparatus with antibiotics (20 units penicillin and 20 μg streptomycin/mL buffer). Samples were withdrawn at the indicated times for determination of DCI, MetHb (18), and hemolysis (16) and for 2-D PAGE (19). pH was measured in the erythrocyte suspension before and after incubation using a Radiometer ABL505 pH meter (Copenhagen, Denmark).
Erythrocyte DCI.
DCI was determined as a DFO-iron complex, as previously reported (16). Briefly, at the end of incubation, 25 μM DFO was added to samples. The erythrocytes were then hemolyzed by adding water (1 vol) and thawing from −70°C. The hemolysate was ultrafiltered in centrifugal filter devices (Centriplus YM-30; Amicon, Millipore, Bedford, MA) and the excess of DFO removed by silica column chromatography. The DFO-iron complex was determined by HPLC. HPLC conditions were as follows: 120–140 bars, flow rate 1.5 mL/min, silica column 5 μm (250 mm × 4 mm), retention time 7–8 min.
Statistical analysis.
Results were reported as means ± SEM. Comparisons between groups were carried out by t test. The value of p < 0.05 was considered statistically significant.
High-resolution 2-D gel electrophoresis.
Erythrocyte suspension (200 μL) was withdrawn before incubation (time 0) and after 16 and 24 h of incubation and was centrifuged at 2500 × g for 5 min. 2-D gel electrophoresis was carried out on a 7-μL aliquot of erythrocyte pellet mixed with 483 μL of a denaturing solution containing 8 M urea, 4% (wt/vol) 3-(3-cholamido propyl) dimethylammonio-1-propane sulfonate (CHAPS), 40 mM Tris-base, 65 mM DTE. Total protein assay was achieved by a standard procedure according to the method of Bradford (20). The quantity of erythrocyte proteins loaded for first dimension separation was 60 μg.
2-D electrophoresis was performed as described (21) using the Immobiline-polyacrylamide system (Amersham Biosciences AB, Uppsala, Sweden) Isolectric focusing (first dimension) was carried out on nonlinear wide-range immobilized pH gradients (IPG strips pH 3–10; 18 cm-long) using the IPGphor system (Amersham Biosciences AB, Uppsala, Sweden). After electrophoresis, IPG strips were equilibrated for 12 min in 6 M urea, 30% (vol/vol) glycerol, 2% (wt/vol) SDS, 0.05 M Tris-HCl, pH 6.8, 2% (wt/vol) DTE, and subsequently for 5 min in the same urea/SDS/Tris buffer solution with 2.5% (wt/vol) iodoacetamide instead of 2% (wt/vol) DTE. The second dimension was carried out on 9–16% polyacrylamide linear gradient gels (18 cm × 20 cm × 1.5 mm) at 40 mA/gel constant current and 10°C until the dye front reached the bottom of the gel (21). Gels were stained with ammoniacal silver nitrate as described (22).
Immunoblot analysis.
After 2-D electrophoresis, proteins were electroblotted onto nitrocellulose membrane as described (23). Before immunodetection, membranes were reversibly stained with 0.2% (wt/vol) Ponceau S in 3% (wt/vol) trichloroacetic acid (TCA) for 3 min and spot position was marked to facilitate computer-aided matching of immunoreactive spots on the silver-stained gels. Working dilution for mouse MAb to anti-pTyr (Santa Cruz Biotechnology, Santa Cruz, CA) was 1:1000. Immunoreactive spots were detected using goat anti-mouse IgG (working dilution 1:3000) conjugated with horseradish peroxidase (Bio-Rad, Hercules CA) and a chemiluminescence detection system (Amersham Pharmacia Biotech UK Ltd., Little Chalfont, UK), according to the manufacturer's instructions.
Computer-aided image acquisition, data analysis, and protein identification.
Electrophoretic images were digitized using a Molecular Dynamics (Amersham Biosciences AB, Uppsala, Sweden) computing densitometer (4000 × 5000 pixels; 12 bits/pixel), and analyzed on a SUN SPARC station 10 with Melanie II (Bio-Rad Laboratories, Milan, Italy) software, which automatically detects, quantifies, and compares protein spots in digitized 2-D images.
Gel matching is a key operation in 2-D image analysis: it consists of comparing gel images and finding “pairs” of spots representing the same protein in both gels. The matching program Melanie II was also used for protein identification by comparison with 2-D reference maps (24).
Protein identification was based on matching electrophoretic coordinates (pI and Mr directly related to amino acid composition) of spots on our map with known spots on a 2-D reference map of human red cells, available in the ExPASy World Wide Web molecular biology server of Geneva University Hospital and the University of Geneva (Switzerland).
RESULTS
DCI release and MetHb formation in newborn erythrocytes.
DCI release and MetHb formation in newborn RBC exposed to hypoxia-reoxygenation or aerobic conditions (control) are reported in Table 1. Aerobic incubation was associated with progressive DCI release and MetHb formation. Hypoxic incubation for 16 h induced significantly higher DCI release than aerobic incubation for 16 h. Reoxygenation for 8 h, after 16 h of hypoxia, induced a further but nonsignificant increase in DCI. MetHb formation roughly paralleled iron release. No hemolysis or significant changes in pH were observed throughout the experiment.
2-D SDS-PAGE and immunoblot analysis.
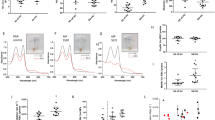
No substantial differences were observed among silver-stained 2-D SDS-PAGE electrophoretograms of RBC of healthy newborns. Figure 1 shows a typical silver-stained 2-D electrophoretogram of RBC proteins. This electrophoretogram only shows proteins soluble in conventional denaturating solution. Membrane proteins, such as band 3 protein, were not present on 2-D PAGE maps, because they are not solubilized during sample preparation as previously reported (25). RBC soluble proteins were identified by matching with a 2-D reference map of human RBC proteins using Melanie II software (24).

Typical 2-D silver-stained electrophoretogram of RBC proteins of healthy term newborn. Identified proteins are indicated.
Different immunoreactivity patterns with the anti-pTyr antibody were observed between RBC incubated under aerobic and hypoxic/aerobic (reoxygenation) conditions. A few weakly immunoreactive proteins were observed in RBC at time 0 (catalase, glyceraldehyde 3-phosphate dehydrogenase, and α, β and β/γ Hb chains, Fig. 2A), after 16 h (catalase, α, β and β/γ Hb chains, Fig. 2B), and after 24 h of aerobic incubation (α, β and β/γ Hb chains, Fig. 2C). In contrast, many strongly immunoreactive proteins were observed after 16 h of hypoxic incubation (acetylcholinesterase, catalase, glyceraldehyde 3-phosphate dehydrogenase, antioxidant protein 2, biliverdin reductase, α, β, and β/γ Hb chains, β-actin, URA 6, and thioredoxin peroxidase 1) (Fig. 2D). After subsequent reoxygenation for 8 h, five proteins (acetylcholinesterase, antioxidant protein 2, α, β and β/γ Hb chains, and URA 6) had decreased immunoreactive intensity and two proteins (β-actin and thioredoxin peroxidase 1) lost all immunoreactivity (Fig. 2E). Proteins disappearing from blots after further aerobic incubation of RBC could be due to metabolization of these proteins as occurs in RBC aging (26). The different patterns of Tyr-phosphorylated proteins are shown in Table 2.

2-D immunoblotting with anti-pTyr MAb of RBC. RBC from healthy term newborn before incubation (A), and incubated for 16 h in aerobic conditions (B), for 24 h in aerobic conditions (C), for 16 h in hypoxic conditions (D), and for 16 h in hypoxic and subsequently for 8 h in aerobic conditions (E). Identified immunoreactive proteins are indicated by numbers (1: acetylcholinesterase, 2: catalase, 3: G3PDH, 4: AOP2, 5: BVR, 6: Hb α-chain, 7: Hb β-chain, 8: Hb β/γ-chain, 9: β-actin, 10: URA 6, 11: Tpx 1).
DISCUSSION
RBC were the first cells in which susceptibility of neonates to oxidative stress was demonstrated (5–7). In the present study, newborn RBC incubated under hypoxic condition showed higher DCI release and MetHb formation than RBC incubated under aerobic conditions, as expected. Reoxygenation of hypoxic erythrocytes induced a further but not significant increase in iron release. These results are in line with previous in vivo studies in which we found higher DCI release and MetHb formation in RBC of hypoxic than normoxic babies (5–7) and they confirm that hypoxic environments are a factor promoting iron release. Because iron release is related to oxidative stress, hypoxia paradoxically seems to be a condition of oxidative stress.
Because PTK is abundant in RBC (11), we investigated Tyr-phosphorylated components of the RBC protein map. We tested the hypothesis that a hypoxic environment and subsequent reoxygenation promote post-translational changes in the newborn RBC protein map. This was suggested by oxidative stress occurring in our in vitro model simulating the perinatal hypoxia-reperfusion event. It has also been suggested that ROS may interfere with intracellular signalling pathway, switching on Tyr-phosphorylation (27), and, as mentioned above, that the superoxide radical is formed under hypoxic conditions (9).
Proteomics is a powerful procedure for investigating complex processes in cells and tissues. Reversible protein tyrosine phosphorylation is a widespread mechanism of modulation of protein/enzyme activity. Protein phosphorylation is the principal mechanism controlling the rate of metabolic enzyme pathways. Phosphate is a negatively charged residue that interacts with proteins to create a conformational change that switches protein activity on and off. Most enzymes regulated by phosphorylation have multiple phosphorylation sites. For some enzymes, the phosphorylation sites are antagonistic, for others they are synergistic, and phosphorylation at one site may act synergistically with phosphorylation at another site.
Our results showed different immunoreactivity patterns in newborn RBC incubated under aerobic and hypoxic conditions. All immunoreactive proteins identified by immunoblot in the RBC protein map were Tyr-phosphorylated after 16 h of hypoxic incubation. Some are involved in cell defence against hypoxia, such as G3PDH (glycolytic enzyme) or against oxidative stress, such as catalase, AOP2, BVR, and Tpx 1.
What this means is debatable. Indeed, although chemiluminescent detection did not enable precise quantification, the changes we observed in the RBC protein map after hypoxic incubation may be explained as a defensive reaction of neonatal RBC against ROS. In another study, we reported that hypoxia remodels the signaling pathway by switching on Tyr-phosphorylation of some proteins (28).
Studies by Zylinska et al. (29) have provided some evidence of a potential role of asphyxia in disrupting phosphorylation processes in newborn erythrocytes, as reflected by altered protein kinase activities in erythrocyte membranes of asphyxiated newborns.
We do not know whether Tyr-phosphorylation means activation or deactivation, but phosphorylation of G3PDH strongly suggests a hypoxia-induced increase in enzyme activation. A major substrate for human erythrocyte PTK is reported to be the anion transporter band 3 protein (11). G3PDH is associated reversibly with the N-terminus of the cytoplasmic domain of band 3 (30). Once the enzyme is bound, it is totally inhibited; on release, however, it is restored to full activity. Control of the G3PDH binding site and consequently glycolytic flux through this control point is achieved by phosphorylation of Tyr 8 and Tyr 21 at the G3PDH binding site on band 3. Phosphorylation blocks enzyme binding, activating G3PDH. Other authors have observed that deoxygenation increases the degree of Tyr phosphorylation of band 3 in RBC (31) with an increase in glycolytic enzyme activities (11). Gonzalez et al. (32) observed that exposure to hypobaric hypoxia followed by reoxygenation induced inhibition of band 3 anion transport function and a decrease in binding of G3PDH to band 3. In the present study, we observed increased G3PDH Tyr-phosphorylation in neonatal RBC samples after 16 h of hypoxic incubation as well as after 8 h of reoxygenation with respect to starting level at time 0. No G3PDH Tyr-phosphorylation was observed in samples after aerobic incubation (Table 2). Phosphorylated G3PDH, released by band 3 protein, consequent to hypoxia, may therefore enhance glycolysis.
New types of antioxidant proteins have recently been reported in RBC and other cells: the thiol-specific antioxidant proteins or as recently renamed, Prxs (33). Prxs detoxify alkyl-hydroperoxides, peroxynitrates, and hydrogen peroxides using reducing equivalents from thiol-containing molecules, such as thioredoxins and glutathione. In addition to their antioxidant activity, Prxs have been implicated in a number of cellular functions such as cell proliferation and differentiation, neuronal apoptosis, protection of radical-sensitive proteins, heme metabolism, and intracellular signaling. Prxs may be regulated by changes in phosphorylation, redox, and possibly oligomerization state (33). Six subclasses of mammal Prxs (Prx I–VI) are considered.
Prx VI (AOP2) and Prx II (Tpx1) are members of this protein family. Stuhlmeier et al. (34) have demonstrated that AOP2 is a cytoplasmic protein found in lymphocytes, endothelial cells, and RBC. It protects RBC from ROS in a dose-dependent manner by binding to Hb, particularly to heme, thus preventing induced and spontaneous oxidation to MetHb. Heme is the prosthetic group of several proteins and enzymes (such as myoglobin, cytochrome c and cytochrome P450, and others), and AOP2 may protect them also. This would explain its nearly ubiquitous presence and provide further evidence of its importance as a major protective protein.
We observed that AOP2 was Tyr-phosphorylated after hypoxia (Table 2), when DCI release and MetHb formation are high (Table 1), suggesting that hypoxia induces Tyr-phosphorylation which presumably leads to a change in antioxidant function with consequent oxidation of Hb. Tyr-phosphorylation could also involve the α, β, and γ chains of Hb that are more Tyr-phosphorylated under hypoxia than normoxia.
Tpx 1 is another antioxidant protein that limits the activity of ROS. Studies by Ichimiya et al. (35) showed that mouse Tpx had broad tissue distribution, but that its expression was especially marked in cells very active in metabolizing oxygen molecules, such as erythroid cells, renal tubule cells, heart and skeletal muscle cells, and certain types of neurons. Increased expression of Tpx has been observed in areas of the brain most susceptible to hypoxic-ischemic injury in humans. The authors proposed that Tpx contributes to antioxidant defense in erythrocytes and neurons by limiting the destructive capacity of oxygen radicals. Lee et al. (36) showed that Prx II−/− mice have hemolytic anemia and that Prx II plays a major role in protecting RBCs from oxidative stress in mice. In our samples, Tpx 1 was partially Tyr-phosphorylated after 16 h of hypoxia, suggesting that its phosphorylation may increase, counteracting hypoxia-induced stress. No Tyr-phosphorylation was observed after reoxygenation.
Catalase occurs in almost all aerobic organisms and is an important factor of cellular defence against hydrogen peroxide (H2O2). One antioxidant role of catalase is to lower the risk of hydroxyl radical formation from H2O2 via the Fenton-reaction catalyzed by Cu or Fe ions. Recent in vitro studies by Cao et al. (37) have proven that Tyr-phosphorylation is required for catalase activation and thereby degradation of H2O2. In our experiments, we found that catalase is Tyr-phosphorylated. According to Cao et al., this could mean an increase in antioxidant defences as a consequence of DCI release after hypoxic incubation and reoxygenation.
Studies by Shalloe et al. (38) show that human BVR and FR, isolated from bovine erythrocytes, are identical. Human FR catalyzes the NADPH-dependent reduction of flavin-mono-nucleotide and methylene blue and, in the presence of redox couplers, reduction of MetHb. FR has been shown to be identical with a major RBC protein of unknown function termed “green heme-binding” protein, found in erythrocytes and liver (39). BVR may therefore have a role in protecting cells from oxidative damage. In our experiments, BVR was not Tyr-phosphorylated during aerobic incubation, when MetHb formation was low (Table 1), but it was Tyr-phosphorylated after hypoxia and hypoxia-reoxygenation, when concentrations of MetHb were higher than during aerobic incubation. This suggests that Tyr-phosphorylation may induce changes in enzyme activity.
AChE, a key component of cholinergic transmission, is responsible for the Yt blood group system. A recent study suggests that AChE is an excellent enzyme marker for ageing of human RBC (26). We observed that AChE was Tyr-phosphorylated after 16 h of hypoxia, when DCI release is high, as it is in erythrocyte aging (3), suggesting that Tyr-phosphorylation could modify enzyme activity.
In our study, another protein Tyr-phosphorylated after 16 h of hypoxia was URA6. URA6 is a fragment of a RBC protein of unknown function.
Beta-actin is an isoform of actin protein. Three main groups of actin isoforms, alpha, beta, and gamma, have been identified. Alpha actins are found in muscle and are a major constituent of the contractile system. Beta and gamma actin isoforms coexist in most cell types as components of the cytoskeleton and as mediators of internal cell motility. One of the many targets of tyrosine kinase–mediated signaling pathways is the actin cytoskeleton. Actin cytoskeletal rearrangements are important fundamental events that occur during a variety of cellular processes (40). Beta-actin is Tyr-phosphorylated after 16 h of hypoxia in RBC of newborns, which suggests that hypoxia could induce cytoskeletal reorganization.
In conclusion, high-resolution 2-D gel electrophoresis of neonatal RBC incubated under hypoxic conditions revealed a complex pattern of Tyr-phosphorylation in the protein map. Hypoxia may possibly increase Tyr-phosphorylation of antioxidant proteins in RBC of newborns. We hypothesize that this increase is a cellular response of neonatal RBC to oxidative stress. To our knowledge, this is the first report describing the 2-D map of Tyr-phosphorylated soluble proteins of neonatal RBC after hypoxia. These findings may help to understand the molecular events underlying hypoxia-induced modification of neonatal RBC. As observed in brain tissue (28), similar mechanisms could be implicated in perinatal hypoxic-ischemic damage of other cells. Further research using different methodologies is needed to clarify overall membrane protein expression in RBC of newborns.
Abbreviations
- AChE:
-
acetylcholinesterase
- anti-pTyr:
-
anti-phosphotyrosine
- AOP2:
-
antioxidant protein 2
- BVR:
-
biliverdin IX-β reductase
- DCI:
-
desferrioxamine-chelatable iron
- DFO:
-
desferrioxamine
- DTE:
-
dithio-erythritol
- FR:
-
flavin reductase
- G3PDH:
-
glyceraldehyde 3-phosphate dehydrogenase
- IPG:
-
immobilized pH gradient
- MetHb:
-
methemoglobin
- Prxs:
-
peroxiredoxins
- PTK:
-
protein tyrosine kinase
- RBC:
-
red blood cells
- ROS:
-
reactive oxygen species
- Tpx 1:
-
thioredoxin peroxidase 1
- Tyr:
-
tyrosine
- 2-D:
-
two-dimensional
- 2,3-DPG:
-
2,3-diphosphoglycerate
References
Ryan TP, Aust SD 1992 The role of iron in oxygen-mediated toxicities. Crit Rev Toxicol 22: 119–141
Minotti G 1993 Sources and role of iron in lipid peroxidation. Chem Res Toxicol 6: 134–146
Comporti M, Signorini C, Buonocore G, Ciccoli L 2002 Iron release, oxidative stress and erythrocyte ageing. Free Radic Biol Med 32: 568–576
Ciccoli L, Signorini C, Scarano C, Rossi V, Bambagioni S, Ferrali M, Comporti M 1999 Iron release in erythrocytes from patients with β-thalassemia. Free Radic Res 30: 407–413
Ciccoli L, Rossi V, Leoncini S, Signorini C, Paffetti P, Bracci R, Buonocore G, Comporti M 2003 Iron release in erythrocytes and plasma non protein-bound iron in hypoxic and non hypoxic newborns. Free Radic Res 37: 51–58
Buonocore G, Zani S, Sargentini I, Gioia D, Signorini C, Bracci R 1998 Hypoxia-induced free iron release in the red cells of newborn infants. Acta Paediatr 87: 77–81
Buonocore G, Zani S, Perrone S, Caciotti B, Bracci R 1998 Intraerythrocyte nonprotein-bound iron and plasma malondialdehyde in the hypoxic newborn. Free Radic Biol Med 25: 766–770
Ciccoli L, Rossi V, Leoncini S, Signorini C, Blanco-Garcia J, Aldinucci C, Buonocore G, Comporti M 2004 Iron release, superoxide production and binding of autologous IgG to band 3 dimers in newborn and adult erythrocytes exposed to hypoxia and hypoxia-reoxygenation. Biochim Biophys Acta 1672: 203–213
Balagopalakrishna C, Manoharan PT, Abugo OO, Rifkind JM 1996 Production of superoxide from hemoglobin-bound oxygen under hypoxic conditions. Biochemistry 35: 6393–6398
Arnaud J, Gutierrez N 1984 Human red cell glycolysis in high altitude chronic hypoxia. Am J Phys Anthropol 63: 307–314
Harrison ML, Rathinavelu P, Arese P, Geahlen RL, Low PS 1991 Role of band 3 tyrosine phosphorylation in the regulation of erythrocyte glycolysis. J Biol Chem 266: 4106–4111
De Franceschi L, Fumagalli L, Olivieri O, Corrocher R, Lowell CA, Berton G 1997 Deficiency of Src family kinases Fgr and Hck results in activation of erythrocyte K/Cl cotransport. J Clin Invest 99: 220–227
Bordin L, Clari G, Moro I, Dalla Vecchia F, Moret V 1995 Functional link between phosphorylation state of membrane proteins and morphological changes of human erythrocytes. Biochem Biophys Res Commun 213: 249–257
Thannickal VJ, Aldweib KD, Fanburg BL 1998 Tyrosine phosphorylation regulates H2O2 production in lung fibroblasts stimulated by transforming growth factor beta 1. J Biol Chem 273: 23611–23615
Godovac-Zimmermann J, Soskic V, Poznanovic S, Brianza F 1999 Functional proteomics of signal transduction by membrane receptors. Electrophoresis 20: 952–961
Ferrali M, Ciccoli L, Comporti M 1989 Allyl alcohol-induced hemolysis and its relation to iron release and lipid peroxidation. Biochem Pharmacol 38: 1819–1825
Bloom RS 1997 Delivery room resuscitation of the newborn. In: Fanaroff AA, Martin RJ (eds) Neonatal-Perinatal Medicine: Diseases of the Fetus and Infant. CV Mosby, St. Louis, MO, pp 376–402
Evelyn KA, Malloy HT 1938 Microdetermination of oxyhemoglobin, methemoglobin and sulfhemoglobin in a single sample of blood. J Biol Chem 126: 655–662
Bjellqvist B, Pasquali C, Ravier F, Sanchez JC, Hochstrasser D 1993 A nonlinear wide-range immobilized pH gradient for two-dimensional electrophoresis and its definition in a relevant pH scale. Electrophoresis 14: 1357–1365
Bradford MM 1976 A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72: 248–254
Hochstrasser DF, Harrington MG, Hochstrasser AC, Miller MJ, Merril CR 1988 Methods for increasing the resolution of two-dimensional protein electrophoresis. Anal Biochem 173: 424–435
Hochstrasser DF, Patchornik A, Merril CR 1988 Development of polyacrylamide gels that improve the separation of proteins and their detection by silver staining. Anal Biochem 173: 412–423
Towbin H, Staehelin T, Gordon J 1979 Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proc Natl Acad Sci U S A 76: 4350–4354
Appel RD, Palagi PM, Walther D, Vargas JR, Sanchez JC, Ravier F, Pasquali C, Hochstrasser DF 1997 Melanie II—a third-generation software package for analysis of two-dimensional electrophoresis images: I. Features and user interface. Electrophoresis 18: 2724–2734
Rabilloud T, Adessi C, Giraudel A, Lunardi J 1997 Improvement of the solubilization of proteins in two-dimensional electrophoresis with immobilized pH gradients. Electrophoresis 18: 307–316
Prall YG, Gambhir KK, Ampy FR 1998 Acetylcholinesterase: an enzymatic marker of human red blood cell aging. Life Sci 63: 177–184
Brumell JH, Burkhardt AL, Bolen JB, Grinstein S 1996 Endogenous reactive oxygen intermediates activate tyrosine kinases in human neutrophils. J Biol Chem 271: 1455–1461
Buonocore G, Liberatori S, Bini L, Mishra OP, Delivoria-Papadopoulos M, Pallini V, Bracci R 1999 Hypoxic response of synaptosomal proteins in term guinea pigs fetuses. J Neurochem 73: 2139–2148
Zylinska L, Sobolewska B, Gulczynska E, Ochedalski T, Soszynski M 2002 Protein kinases activities in erythrocyte membranes of asphyxiated newborns. Clin Biochem 35: 93–98
Low PS, Geahlen RL, Mehler E, Harrison ML 1990 Extracellular control of erythrocyte metabolism mediated by a cytoplasmic tyrosine kinase. Biomed Biochim Acta 49: S135–S140
Barbul A, Zipser Y, Nachles A, Korenstein R 1999 Deoxygenation and elevation of intracellular magnesium induce tyrosine phosphorylation of band 3 in human erythrocytes. FEBS Lett 455: 87–91
Gonzalez G, Celedon G, Sandoval M, Gonzalez GE, Ferrer V, Astete R, Behn C 2002 Hypobaric hypoxia-reoxygenation diminishes band 3 protein functions in human erythrocytes. Pflugers Arch 445: 337–341
Wood ZA, Schröder E, Robin Harris J, Poole LB 2003 Structure, mechanism and regulation of peroxiredoxins. Trends Biochem Sci 28: 32–40
Stuhlmeier KM, Kao JJ, Wallbrandt P, Lindberg M, Hammarström B, Broell H, Paigen B 2003 Antioxidant protein 2 prevents methemoglobin formation in erythrocyte hemolysates. Eur J Biochem 270: 334–341
Ichimiya S, Davis JG, O'Rourke DM, Katsumata M, Greene MI 1997 Murine thioredoxin peroxidase delays neuronal apoptosis and is expressed in areas of the brain most susceptible to hypoxic and ischemic injury. DNA Cell Biol 16: 311–321
Lee TH, Kim SU, Yu SL, Kim SH, Park do S, Moon HB, Dho SH, Kwon KS, Kwon HJ, Han YH, Jeong S, Kang SW, Shin HS, Lee KK, Rhee SG, Yu DY 2003 Peroxiredoxin II is essential for sustaining life span of erythrocytes in mice. Blood 101: 5033–5038
Cao C, Leng Y, Kufe D 2003 Catalase activity is regulated by c-Abl and Arg in the oxidative stress response. J Biol Chem 278: 29667–29675
Shalloe F, Elliott G, Ennis O, Mantle TJ 1996 Evidence that biliverdin-IXβ reductase and flavin reductase are identical. Biochem J 316: 385–387
Xu F, Quandt KS, Hultquist DE 1992 Characterization of NADPH-dependent methemoglobin reductase as a heme-binding protein present in erythrocytes and liver. Proc Natl Acad Sci U S A 89: 2130–2134
Jamora C, Fuchs E 2002 Intercellular adhesion, signalling and the cytoskeleton. Nat Cell Biol 4: E101–E108
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Marzocchi, B., Ciccoli, L., Tani, C. et al. Hypoxia-Induced Post-Translational Changes in Red Blood Cell Protein Map of Newborns. Pediatr Res 58, 660–665 (2005). https://doi.org/10.1203/01.PDR.0000180545.24457.AC
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1203/01.PDR.0000180545.24457.AC
This article is cited by
-
Alterations in phosphoproteome under salt stress in Thellungiella roots
Chinese Science Bulletin (2010)
