
Abstract
We measured cell surface expression of CD34, HLA-DR, CD38, CD19, CD33, CD71, and CD45 antigens in the hematopoietic progenitor cells of fetal cord blood to investigate immunophenotypic changes at different gestational ages. These antigens were identified by flow cytometry in 11 fetuses (gestational age 19–24 wk, in 12 preterm (25–28 wk) and in ten newborn infants born at term. The frequency and number of CD34+ cells were higher in the blood of the 11 fetuses; in addition, a statistically significant inverse correlation between number of CD34+ cells and advancing gestational age was noted. The numbers of CD34+CD19+, CD34+CD33+, and CD34+CD45+ coexpressing cells were significantly higher in the fetuses, whereas CD34+CD38+ cells were more represented in the neonates at term. Gestational age was inversely correlated with the number of CD34+CD19+ and CD34+CD33+ coexpressing cells. A positive correlation between gestational age and CD34+CD38+ cells was noted. The number of CD34–CD19+, CD34–CD38+, and CD34–CD45+ cells was higher in term infants; furthermore, a significant correlation between advancing gestational age and CD34–CD38+ or CD34–CD45+ cells was demonstrated. The proliferative capacity was also higher at lower gestational ages. These data suggest that the development and lineage commitment of fetal cord blood hematopoietic progenitor cells are very active during the last two trimesters of pregnancy. The most significant changes of hematopoietic cells maturation seem to occur within 25 wk of gestation.
Similar content being viewed by others

Main
Human umbilical cord blood (HUCB) has been used as an alternative source for bone marrow transplantation in children with various malignant and nonmalignant disorders due to the high number of early and committed hematopoietic progenitor cells (1, 2).
Hematopoietic cells appear in the embryo at the third to fourth week of gestation, they migrate from the yolk sac to the liver and spleen during the fifth to 12th week, and ultimately they reach the bone marrow throughout fetal circulation at the second to third trimester of gestation.
T lymphocyte differentiation initiates in the thymus after the seventh week of gestation, but T cells do colonize the fetal liver, spleen, and bone marrow only later than the 13th week. Helper and suppressor activities of fetal thymocytes and splenocytes are acquired between the 12th and the 16th week. HLA antigens are firstly detected in the lymphocytes at the beginning of the second trimester (3). The mechanisms regulating the ontogenetic shifts in hematopoiesis are still unknown, and it remains uncertain whether progenitor cells circulating in fetal blood include the most primitive progenitor cell compartments to colonize the bone marrow (4).
The development of hematopoietic progenitor cells has been evaluated in fetal HUCB by analysis of the proliferative capacity and the expression of cell surface antigens that can reflect the lineage commitment and the maturation. Haneline et al. (5), Migliaccio et al. (4), and Shields et al. (6) suggested that the frequency and the proliferative capacity of primitive hematopoietic progenitors in the cord blood was higher in extremely low gestational age fetuses.
However, little is known about the onset and the variability of cell surface antigen expression at different gestational ages. The aim of our study was to investigate the immunophenotypic changes and the proliferative capacity of cord blood hematopoietic progenitor cells in three populations: fetuses (gestational age 19–24 wk), preterm (25–29 wk), and newborn infants at term.
METHODS
Umbilical cord blood was collected in sterile heparinized tubes from three groups of infants:
-
1. Eleven spontaneously aborted fetuses (gestational age 19–24 wk, mean 21.2 wk): maternal history was negative for significant disease and no biochemical or ultrasound abnormality was noted; gross malformations were excluded by inspection.
-
2. Twelve preterm newborns (gestational age 25–28 wk, mean 27.1 wk): nine babies were born by elective caesarean section and three by spontaneous preterm delivery with labor duration less than 6 h.
-
3. Ten term neonates born by elective caesarean section after uncomplicated pregnancy (gestational age 38–42 wk, mean 39.8 wk).
We excluded infants who were born after prolonged labor (>6 h) because HUCB cell populations can be influenced by stress in these circumstances (7).
Term or preterm newborns with malformations or congenital hematological or immunologic diseases were excluded from the study. The cord blood samples were collected only in the absence of maternal or intrapartum infection or illness. Gestational age was assessed by maternal dates and ultrasound examination.
Leukocyte counts were obtained by using an automatic analyzer.
Cell preparation.
Leukocytes from heparinized cord blood samples were separated by Ficoll gradient centrifugation (8) and cryopreserved with 10% DMSO, at −180°C for less than 1 mo before analysis. After rapid thawing at 37°C, cells were incubated in RPMI 1640 media (GIBCO, Grand Island, NY), combined with 2 mmol/L glutamin and 50 μg/mL gentamicin, with 10% FCS for 40 min and then centrifuged at 300 g for 10 min and resuspended in 1 mL of RPMI with FCS. Cell numbers and viability were determined using ethidium bromide/acridine orange staining. Cells were divided in aliquots and incubated for 30 min at 4°C with FITC and phycoerythrin (PE) conjugated MAb.
The hematopoietic cell surface expression of the following antigens was evaluated: anti-HLA-DR FITC (MHC II antigen), CD19 FITC (immature B cells), CD71 FITC (transferrin receptor, erythroid progenitor cells), CD45 FITC (leukocyte common antigen), CD8 FITC, CD34 PE (hematopoietic progenitor/stem cell), CD4 PE (Becton Dickinson, Oxford, UK), CD38 FITC (early differentiating and activating cells), CD33 FITC (myeloid progenitor cells; Immunotech, Marseille, France).
Aliquots of cells were labeled with CD34 and one of the MAb FITC. After incubation, cells were washed with PBS containing 0.2% BSA and NaN3 (PBS); the pellet with the immunofluorescent-labeled cells was resuspended in 0.5 mL PBS for double-staining flow cytometric analysis.
Flow cytometry.
Flow cytometric analysis was performed on a FACScan flow cytometer (Becton Dickinson). Forward light-scattering, orthogonal light-scattering, and two fluorescence signals were determined for each cell and analyzed using Cell Quest software (Becton Dickinson). Fluorescence compensation on the flow cytometer was adjusted using CD4 and CD8 MAb to minimize overlap of the FITC and PE signals. FITC and PE-conjugated isotype-matched MAb were used as negative control. A total of 10 000 events was acquired for each sample.
The forward (size) and side scatter (granularity) properties were used to define windows L (L = lymph, which would select lymphocytes if the analyzed sample was peripheral blood), and B (B = blast, a population of cells that is large by forward scatter, with moderate amounts of right-angle light scattering) (9, 10).
The frequency of antigen expressing or not expressing cells was recorded for each sample and the absolute number of cells was calculated as percentage of the total leukocyte count.
In vitro progenitor cell cultures.
HUCB cells were isolated by separation on Lymphocyte Separation Medium Flow (density 1077 g/mL) and centrifuged at 400 × g for 20 min at room temperature (20°C). HUCB were washed three times and resuspended in Iscove's Modification of Dulbecco Medium (IMDM) (Flow). Then, 1 mL of complete medium, with cells concentrated at 105 cells/mL, in IMDM containing 0.8% methylcellulose (MethoCult H4230, StemCellTechnologies Inc., Vancouver, BC, Canada), eythropoietin 2 U/mL (Eprex Cilag, Schaffausen, Svizzera, Sollentuna, Sweden), 1–5% LCM-PHA medium conditioned by peripheral blood leukocytes (HemostimH2400, StemCells Technologies Inc.) as growth factors, was divided in three wells (333 μL/well) of a 24-well tissue culture plate (Falcon Products, Becton Dickinson Lab, Lincoln Park, NJ). The cultures were examined for presence of erythroid burst-forming units (BFU-E), granulocyte-erythroid-macrophage-megakaryocyte colony-forming units (CFU-GEMM), and granulocyte-macrophage colony-forming units (CFU-GM) with an inverted microscope after incubation for 14 d in a humidified atmosphere supplemented with 5% CO2. CFU-GM were defined on their capability to produce nonhemoglobinized colonies containing a minimum of 40 clonogenic progenitors of granulocytes/macrophages. BFU-E were defined by their ability to give intermediate-sized hemoglobinized colonies that contained at least three erythroblast clusters. CFU-GEMM were defined as colonies containing multiple lineages of cells, including erythroid cells.
The study was approved by the Ethical Committee of Pavia Teaching Hospital and informed parental consent was obtained before each cord sampling.
The statistical analysis was performed using the one way ANOVA, followed by the “post hoc” Newman-Keuls test to evaluate differences of mean antigen expression among the three groups of infants in windows L and B. Relationships were assessed with the Pearson correlation coefficients; linear regression lines were fitted by the least square method. A p value > 0.05 was considered not significant. All computations were performed using a commercial statistical package (STATISTICA for Windows, StatSoft, Inc., Tulsa, OK).
RESULTS
The majority of cells were gated in window L (Table 1). The number of gated cells in windows L or B achieved significant statistical difference between preterm and term neonates (Table 1).
The absolute number and frequency of CD34+ cells were significantly higher in very preterm fetuses compared with the other infants, although no significant difference was observed between preterm infants older than 25 wk of gestation and term infants (Table 1). A significant negative correlation was found between the absolute number of CD34+ cells and gestational age (r = −0.411;p < 0.018) (Fig. 1).

Correlation between number of CD34+ cells/μL in window B and gestational age. Regression and 95% confidence interval lines are reported. n = 33 data points;r = −0.411;p < 0.018.
We observed in window L:
Table 2. CD34+ noncoexpressing cells (window L)
-
A significantly higher number of CD34+CD19–, CD34+CD38–, and CD34+CD71– cells in fetuses below 25 wk of gestation (Table 2).
Table 2 CD34+ coexpressing cells (window L) Results are expressed as mean ± SD. *p < 0.02 vs >25 wk and term infants. †p < 0.01 vs >25 wk and p < 0.02 vs term infants. ‡p < 0.01 vs >25 wk infants. -
The number of CD34+CD19+, CD34+CD33+, and CD34+CD45+ coexpressing cells revealed a quick reduction reaching statistical differences between <25/>25 wk groups, whereas CD34+CD38+ cells increased in term infants (Table 3). Gestational age was inversely correlated with the number of CD34+CD19+ (r = −0.415;p < 0.017) and CD34+CD33+ coexpressing cells (r = −0.357;p < 0.042). Gestational age was positively correlated with the number of CD34+CD38+ coexpressing cells (r = 0.408;p < 0.019). The decrease of CD34+CD19+ cells was inversely correlated with the increase of CD34+CD38+ cells (r = −0.361;p < 0.039).
Table 3 CD34− cells (window L) Results are expressed as mean ± SD. *p < 0.01 vs >25 wk and term infants. †p < 0.01 vs >25 wk and term infants. ‡p < 0.01 vs <25 wk and >25 wk infants. §p < 0.04 vs >25 wk infants. ¶p < 0.04 vs term infants. -
The number of CD34–CD19+, CD34–CD38+, and CD34–CD45+ cells was significantly higher in term infants (Table 4), with a significant correlation between gestational age and CD34–CD38+ (r = 0.755; p= < 0.001) or CD34–CD45+ cells (r = 0.714;p < 0.001) (Fig. 2).
Table 4 CD34− cells (window L) Results are expressed as mean ± SD.0.04 vs >25 wk infants. †p < 0.001 vs <25 wk and term infants. ‡p < 0.01 vs <25 wk and >25 wk infants. †p < 0.001 vs <25 wk and >25 wk infants. ‡p < 0.001 vs >25 wk and p < 0.01 vs >25 wk infants. Figure 2 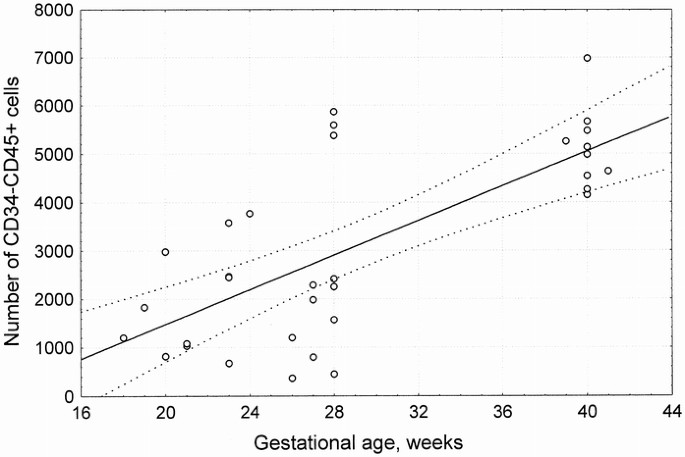
Correlation between number of CD34–CD45+ cells/μL in window B and gestational age. Regression and 95% confidence interval lines are reported. n = 33 data points;r = 0.714;p < 0.001.

In window B we observed no significant differences between the number of noncoexpressing or coexpressing cells, although a significant decrease of CD34+CD45+ coexpressing cells in >25 wk and term infants groups compared with <25 wk group was noted. Furthermore, the numbers of CD34–CD33+, CD34–HLA-DR+, CD34−CD38+, and CD34−CD45+ cells were significantly higher in term infants compared with infants born <25 and >25 wk of gestation (data not shown).
Results concerning in vitro cell cultures are reported in Table 5. The proliferative capacity was significantly higher in groups 1 and 2 compared with term infants.
DISCUSSION
The expression or absence of specific cell surface antigens is commonly used to characterize the highly heterogenic hematopoietic progenitor/stem cell compartments. Previous studies showed that hematopoietic progenitor cells usually express the CD34 surface antigen and that the so-called stem cells may be identified by the absence of CD38, HLA-DR, and lineage committed antigens on CD34+ cells (11). The CD34+CD38– immunophenotype represents a rare, quiescent, and functionally primitive population of progenitor cells that seems to have a higher cloning efficiency and a very rapid proliferative response to cytokine stimulation (12). Primitive progenitors have been found in CD34+CD38+ population of cord blood, although they appear to have a reduced proliferative capacity compared with CD34+CD38– cells (13).
We evaluated cell surface expression of the CD34, HLA-DR, CD38, CD19, CD33, CD71, and CD45 antigens aiming to identify the degree of development of hematopoietic cells from progenitor/stem cells to lineage committed cells. Our results showed that both the percentage and the absolute number of CD34+ cells detectable in HUCB were significantly higher in very preterm fetuses <25 wk of gestation compared with more mature preterm and term newborns. In addition, a statistically significant correlation between number of CD34+ cells and advancing gestational age was noticed. We observed that the highest proliferative activity was found at the lowest gestational ages, especially for the most undifferentiated hematopoietic stem cells; in addition, the proliferative activity progressively decreased with advancing gestational age. These results are in accordance with those of Haneline et al. (5) and Opie et al. (14). Both studies reported a significantly higher frequency of primitive progenitors in the circulating blood cells of surviving preterm infants (GA 23–32) and stillborn fetuses (<24 wk) compared with the cord blood of mature newborns. Our data are also in agreement with Shields et al. (6) and Thilaganathan et al. (15) as they observed a declining frequency of CD34+ cells with advancing gestational age. Meister et al. (16) showed a higher percentage of CD34+ cells in preterm infants with 29–35 wk of gestation compared with term newborns. In Meister's study, however, the number of CD34+ cells was not different between term and preterm infants. Our data differed from Meister's (16) in that we found both the frequency and number of CD34+ cells were significantly higher in preterm infants; this may reflect the lower gestational age of our subjects.
In the present study the hematopoietic progenitor cells of cord blood were detected at various stages of development, although their immunologic phenotype consistently changed with the advancing gestational age.
The increased number in window L of CD34+ cells noncoexpressing the CD19, CD38, or CD71 antigen at the lowest gestational ages together with the higher number of CD34– cells expressing various markers of lineage commitment (CD19, CD38, and CD45 in window L, and CD33, HLA-DR, CD38, and CD45 in window B) in term infants seem to indicate a progressive maturation and commitment of the hematopoietic progenitors with advancing gestational age. The highest concentration of stem cells is likely to be found in the most preterm fetuses.
The presence of hematopoietic progenitor cells at various stages of development seems to support the migratory theory of ontogenesis of the human hematopoietic system. The relatively high number of the most primitive progenitor cells (CD34+ noncoexpressing cells) at the very low gestational ages may relate to the migration process that starts in the fetal liver and only in a later stage colonizes the marrow (4). In addition, the presence of colonies at 19–25 wk of gestation in fetal blood supports the hypothesis that the transfer from one anatomic location of hematopoiesis to another is via fetal circulation.
Our findings seem to reproduce the studies of Thilaganathan et al. (15) and Opie et al. (14), showing an increased frequency of CD34+ cells coexpressing the HLA-DR antigen with advancing gestational age, and higher concentration of CD34+CD38– and CD34+HLA-DR– in the youngest fetal hematopoietic progenitor cells.
The distribution of cells showed a higher percentage of gated cells in window L than in window B in preterm babies compared with term infants. This finding is in accordance with the physical features of CD34+ HUCB cells shifting toward window B with advancing maturational stage as observed by Meister et al. (16).
Furthermore, our data seemed to show changes in lineage commitment of CD34+ cells from immature B cells (CD19) and myeloid progenitor cells (CD33) toward early differentiating and activated cells (CD38) with advancing gestational age. The number of CD34+CD19+ cells in HUCB of very low gestational age fetuses was significantly higher compared with >25 wk and term infants, although CD34+CD38+ coexpression was positively correlated with gestational age. The decrease in CD34+CD19+ coexpression with advancing gestational age is coincident with the increase in CD34+CD38+ coexpression with a constantly negative correlation between the two coexpressions. This can indicate a more active proliferative response of B lymphocytes during early gestation that could be linked with early production of fetal IgM antibody in response to infections, and could suggest that the humoral immune response may potentially develop before the cellular one. Further studies are required to verify such a hypothesis.
A very high increase of both the percentage and absolute number of B lymphocytes and a slight delay in granulocyte recovery have been reported in children after transplantation of cord blood progenitor cells compared with children who received bone marrow transplantation (17). These findings seem to confirm the relatively high proliferative capacity of B lymphocyte committed cells sampled from the cord blood.
In conclusion, our data seem to show that the extremely preterm fetus has a significantly more immature pool of circulating cord blood hematopoietic progenitor cells compared with term infants. Furthermore, the process of development in fetal blood seems to be very active during the last two trimesters of pregnancy. In our experience, the most significant changes during maturation of hematopoietic cells seem to occur within the 25th week of gestation.
These findings seem to confirm that cord blood cells of very young fetuses may prove useful targets for genetic manipulation and in utero transplantation (18–20).
Abbreviations
- HUCB:
-
human umbilical cord blood
- PE:
-
phycoerythrin
- BFU-E:
-
erythroid burst-forming unit
- CFU-GEMM:
-
granulocyte-erythroid-macrophage-megakaryocyte colony-forming unit
- CFU-GM:
-
granulocyte-macrophage colony-forming unit
References
Rubinstein P, Carrier C, Scaradavou A, Kurtzberg J, Adamson J, Migliaccio AR, Berkowitz RL, Cabbad M, Dobrila NL, Taylor PE, Rosenfield RE, Stevens CE 1998 Outcomes among 562 recipients of placental-blood transplants from unrelated donors. N Engl J Med 339: 1565–1577
Gluckman E, Rocha V, Boyer-Chammard A, Locatelli F, Arcese W, Pasquini R, Ortega J, Souillet G, Ferreira E, Laporte JP, Fernandez M, Chastang C 1997 Outcome of cord-blood transplantation from related and unrelated donors. N Engl J Med 337: 373–381
Royo C, Touraine JL, De Bouteiller O 1987 Ontogeny of T lymphocyte differentiation in the human fetus: acquisition of phenotype and functions. Thymus 10: 57–73
Migliaccio G, Baiocchi M, Hamel N, Eddleman K, Migliaccio AR 1996 Circulating progenitor cells in human ontogenesis: response to growth factors and replating potential. J Hematother 5: 161–170
Haneline LS, Marshall KP, Clapp DW 1996 The highest concentration of primitive hematopoietic progenitor cells in cord blood is found in extremely premature infants. Pediatr Res 39: 820–825
Shields LE, Andrews RG 1998 Gestational age changes in circulating CD34+ hematopoietic stem/progenitor cells in fetal cord blood. Am J Obstet Gynecol 178: 931–937
Lim FT, van Winsen L, Willemze R, Kanhai HH, Falkenburg JH 1994 Influence of delivery on numbers of leukocytes, leukocyte subpopulations and hematopoietic progenitor cells in human umbilical cord blood. Blood Cells 20: 547–558
Kletzel M, Haut P, Atlas M, Olszewski M, Danner-Koptik K, Hubbell M 1997 Red cell depletion of umbilical cord blood (UCB): comparison between unmanipulated and red cell depleted UCB by Ficoll-Paque density gradient separation. J Hematother 6: 269–272
Siena S, Bregni M, Brando B, Ravagnani F, Bonadonna G, Gianni M 1989 Circulation of CD34+ hematopoietic stem cells in the peripheral blood of high-dose cyclophosphamide-treated patients: enhancement by intravenous recombinant human granulocyte-macrophage colony-stimulating factor. Blood 74: 1905–1914
Civin CI, Banquerigo ML, Strauss LC, Loken MR 1987 Antigenic analysis of hematopoiesis. Exp Hematol 15: 10–17
D'Arena G, Musto P, Cascavilla N, Di Giorgio G, Zendoli F, Carotenuto M 1996 Human umbilical cord blood: immunophenotipic heterogeneity of CD34+ hematopoietic progenitor cells. Haematologica 81: 404–409
Almici C, Carlo-Stella C, Wagner JE, Mangoni L, Garau D, Rizzoli V 1997 Biologic and phenotipic analysis of early hematopoietic progenitor cells in umbilical cord blood. Leukemia 11: 2143–2149
Hao QL, Shah AJ, Thiemann FT, Smogorzewska EM, Crooks GM 1995 A functional comparison of CD34+ CD38– cells in cord blood and bone marrow. Blood 86: 3745–3753
Opie TM, Shields LE, Andrews RG 1998 Cell-surface antigen expression in early and term gestation fetal hematopoietic progenitor cells. Stem Cells 16: 343–348
Thilaganathan B, Nicolaides KH, Morgan G 1994 Subpopulations of CD34-positive haematopoietic progenitors in fetal blood. Br J Haematol 87: 634–636
Meister B, Totsch M, Mayr A, Widshwendter M, Huter O, Sperl W 1994 Identification of CD34+ cord blood cells and their subpopulations in preterm and term neonates using three color flow cytometry. Biol Neonate 66: 272–279
Locatelli F, Maccario, Comoli P, Bertolini F, Giorgiani G, Montagna D, Bonetti F, De Stefano P, Rondini G, Sirchia G, Severi F 1996 Hematopoietic and immune recovery after transplantation of cord blood progenitor cells in children. Bone Marrow Transplant 18: 1095–1101
Surbek DV, Hohlfeld P, Gratwohl A, Holzgreve W 1997 Intrauterine transplantation of hematopoietic stem cells for therapy of genetic diseases. Z Geburtshilfe Neonatol 201: 158–170
Touraine JL 1996 Treatment of human fetuses and induction of immunological tolerance in humans by in utero transplantation of stem cells into fetal recipients. Acta Haematol 96: 115–119
Wengler GS, Lanfranchi A, Frusca T, Verardi R, Neva A, Brugnoni D, Giliani S, Fiorini M, Mella P, Guandalini F, Mazzolari E, Pecorelli S, Notarangelo LD, Porta F, Ugazio AG 1996 In-utero transplantation of parental CD34 haematopoietic progenitor cells in a patient with X-linked severe combined immunodeficiency (SCIDXI). Lancet 348: 1484–1487
Acknowledgements
The authors thank Claudia Cova for laboratory support and Antonella Tomasi for help in blood sampling.
Author information
Authors and Affiliations
Rights and permissions
About this article
Cite this article
Gasparoni, A., Ciardelli, L., Avanzini, M. et al. Immunophenotypic Changes of Fetal Cord Blood Hematopoietic Progenitor Cells During Gestation. Pediatr Res 47, 825–829 (2000). https://doi.org/10.1203/00006450-200006000-00024
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1203/00006450-200006000-00024
This article is cited by
-
Term vs. preterm cord blood cells for the prevention of preterm brain injury
Pediatric Research (2017)
-
Age-related Low Expression of HLA-DR Molecules on Monocytes of Term and Preterm Newborns With and Without Signs of Infection
Journal of Perinatology (2003)
