
Abstract
Mechanisms of adrenomedullin-induced increases in fetal pulmonary blood flow were examined in 19 near-term fetal sheep using four key blocker drugs: nitric oxide synthase inhibitor (Nω-nitro-L-arginine), calcitonin gene-related peptide (CGRP) receptor blocker, ATP-dependent potassium (KATP) channel blocker (glibenclamide), and cyclooxygenase inhibitor (indomethacin). Catheters were inserted into the left pulmonary artery and superior vena cava to administer drugs and into the main pulmonary and carotid arteries to measure pressures and heart rate. An ultrasonic flow transducer was placed around the left pulmonary artery to measure flow continuously. Adrenomedullin (mean 1.06 µg/kg) was injected into the left pulmonary artery before and after infusion of Nω-nitro-L-arginine (mean 96.5 mg/kg, n = 6), glibenclamide (mean 11.8 mg/kg, n = 6), CGRP receptor blocker (mean 312.0 µg/kg, n = 6), and indomethacin (mean 1.7 mg/kg, n = 8). Blockade was confirmed by appropriate agonist injection. The adrenomedullin-induced response in left pulmonary artery blood flow was inhibited by Nω-nitro-L-arginine (inhibition rate 99%) and significantly attenuated by glibenclamide (inhibition rate 44%); however, no significant changes were found with CGRP receptor blocker or indomethacin (inhibition rate 0 and 17%, respectively). The responses of the main pulmonary and carotid arterial pressures were similarly affected by those blockers. Our data suggest that in the fetal pulmonary circulation, the adrenomedullin-induced increase in pulmonary blood flow depends largely on nitric oxide release and partly on KATP channel activation, and does not involve the CGRP receptor or a cyclooxygenase-mediated mechanism.
Similar content being viewed by others

Main
During the transition from fetal to neonatal circulation, pulmonary vascular resistance falls rapidly and pulmonary arterial blood flow increases approximately 8- to 10-fold (1,2). Although the mechanism of this dramatic change in the pulmonary circulation is still not fully elucidated, it has been suggested that endogenous vasoactive substances play an important role in the transition, e.g. prostacyclin, ATP, bradykinin, acetylcholine, or nitric oxide (3–7). Adrenomedullin is a potent vasodilating peptide consisting of 52 amino acids (8). This peptide is synthesized mainly in vascular endothelial cells and is distributed throughout many human tissues, especially lung (8,9); however, its role in the transitional pulmonary circulation is not known. In a recent study (10), we demonstrated that exogenously administered adrenomedullin, a novel peptide that is a potent vasodilator, significantly decreased pulmonary arterial blood pressure and increased pulmonary arterial blood flow in fetal sheep, which suggested this peptide might be another vasoactive substance involved in the transition in the pulmonary circulation. Our study was designed to evaluate possible mechanisms of such an adrenomedullin-induced increase in pulmonary arterial blood flow in fetal sheep.
Several studies in adult animals (11–19) have suggested possible mechanisms of adrenomedullin-induced vasodilator, e.g. through contributing actions of cAMP, NO, KATP channels, CGRP receptors, or prostaglandins. These mechanisms still are controversial, and differences in effect are reported between organs and between species (12,20). Further, these studies did not evaluate fetal pulmonary vessels, in which the control of vascular resistance and blood flow may be quite different from that in adult vessels due to the naturally high vascular resistance and low blood flow in the fetus. In this study, we chose four key blocker agents-a NO synthase blocker, a KATP channel blocker, a CGRP receptor blocker, and a cyclooxygenase inhibitor-and examined how these agents affect the adrenomedullin-induced increase in pulmonary arterial blood flow in chronically instrumented fetal sheep.
METHODS
Animals. Nineteen mixed Western pregnant sheep were studied at 126-135 d gestational age (full term is about 145 d). Animal husbandry and the study design followed the guidelines of the National Institutes of Health and were approved by the Committee on Animal Research of the University of California, San Francisco, CA.
Surgical preparation. The surgical preparation has been described in detail (2,10). Briefly, under a combination of ketamine and diazepam anesthesia, a midline laparotomy was performed on the ewe. The fetus was exposed through a small uterine incision; after local lidocaine anesthesia was administered, a skin incision was made in the fetal fore limb to insert catheters. Polyvinyl catheters were advanced into the carotid artery and the superior vena cava from the brachial artery and the brachial vein, respectively, and the incision was closed. Another incision was made over the left chest of the fetus for a left thoracotomy. The pericardium was opened, and Teflon-tipped polyvinyl catheters were inserted directly into the main and left pulmonary arteries. The catheter in the LPA was divided in two using a Y-connector 2 cm distal from the LPA. Formalin (10%) colored with a few drops of sterile indigo carmine solution was infiltrated into the adventitia and media of the ductus arteriosus to prevent vasoactivity during the subsequent experimental protocol (21). An ultrasonic transit time Doppler flow transducer (Transonic Systems, Ithaca, NY) 4-6 mm in diameter was placed around the LPA, then the thoracotomy and fetal skin were closed. Another polyvinyl catheter was placed in the amniotic cavity, and the uterine incision was closed after instilling antibiotics (100 mg gentamicin sulfate, 1 × 106 U penicillin G-K) and warm saline to replace amniotic fluid losses. All vascular catheters were sealed with heparin sodium solution (1000 U/mL) and exteriorized to the left flank of the ewe with the transducer cable. The laparotomy was closed in layers, and the ewe was returned to the cage for recovery. Antibiotics (100 mg gentamicin sulfate, 1 × 106 U penicillin G-K) were administered daily, both i.v. and directly into the amniotic cavity.
Drug preparation. Adrenomedullin (Sigma Chemical Co., St. Louis, MO), CGRP (Sigma), and acetylcholine (Miochol, Iolab Pharmaceuticals, Clairmont, CA) were dissolved in normal saline at a concentration of 10 µg/mL. Cromakalim (Sigma) was dissolved in normal saline at a concentration of 400 µg/mL. LNA (Sigma) was dissolved in normal saline at a concentration of 4 mg/mL. Glibenclamide (Sigma) was dissolved in distilled water at a concentration of 1 mg/mL and titrated to a pH 12-13 by the addition of sodium hydroxide. CGRP8-37 (Sigma) was dissolved in normal saline at a concentration of 50 µg/mL. Indomethacin was dissolved in 100 µL of ethyl alcohol and then suspended with a Tris-buffer solution (pH 8.0) at a concentration of 0.25 mg/mL.
Experimental protocol. We performed four protocols based on the same study design: comparison of adrenomedullin-induced hemodynamic changes before and after infusion of a blocker agent (Fig. 1A-D). A total of 26 studies was performed in 19 fetuses, in which 1 study was performed on 12 fetuses and 2 studies on 7 fetuses. When we performed two studies in the same fetus, they were conducted 2-3 d apart to avoid interaction between studies.

Protocols and examples of LPA blood flow before and after blocker agents (see text for abbreviations).
Experiments were performed 1-2 d after surgery. The ewe was placed in a study cage and allowed free access to alfalfa pellets and water. After a steady-state period of at least 20 min, a blood gas sample was obtained from the carotid artery, and baseline hemodynamic variables were measured: mean LPA blood flow, mean carotid and pulmonary arterial blood pressures, and heart rate. These variables were recorded continuously throughout the experiment. In each of the following protocols, adrenomedullin was injected as a 2-µg bolus (over 5 s) into the LPA. After the experiments, the fetuses were removed from the uterus to measure fetal body weight and to verify catheter positions.
Protocol 1: Effects of NO synthesis inhibition by LNA (Fig. 1A). After baseline measurements, 1 µg acetylcholine, an agonist to NO release and a potent vasodilator in fetal pulmonary arteries (6), was injected as a bolus (over 5 s) into the LPA to produce an increase in LPA blood flow. After LPA blood flow and arterial blood pressures had returned to baseline values, adrenomedullin was injected. After 120 min recovery, 30 mg LNA were injected as a slow bolus (over 5 min) into the LPA followed by a 3 mg/min infusion of LNA into the superior vena cava. At 60 min of continuous infusion of LNA, another bolus of acetylcholine was injected, as before, to check effective blockade of NO release by demonstrating no increase in flow with acetylcholine. Then a second injection of adrenomedullin was given. If the acetylcholine-induced response was not inhibited, LNA infusion was continued for an additional 60 min, then acetylcholine and adrenomedullin injections were repeated.
Protocol 2: Inhibition of potassium-ATP channel by glibenclamide (Fig. 1B). After baseline measurements, 200 µg cromakalim, the agonist of KATP channels (22), was injected as a bolus (over 5 s) into the LPA to produce an increase in LPA blood flow. After LPA blood flow and arterial blood pressures had returned to baseline values, adrenomedullin was injected. After 120 min recovery, 40 mg glibenclamide was infused over 40 min into the LPA. A second bolus of cromakalim was injected into the LPA, as before, to check effective KATP channel blockade by demonstrating lack of response to cromakalim. Then a second injection of adrenomedullin was given. If LPA blood flow increased during the infusion of glibenclamide, cromakalim was injected after LPA blood flow and arterial blood pressures had returned to baseline values. In five of six studies in this protocol, 1 µg acetylcholine was injected before and after the glibenclamide infusion to assess how glibenclamide affects the acetylcholine-induced increase in LPA blood flow.
Protocol 3: Effects of CGRP receptor blockade by CGRP8-37 (Fig. 1C). After baseline measurements, 2 µg CGRP were injected as a bolus (over 5 s) into the LPA to produce an increase in LPA blood flow (23,24). After LPA blood flow and arterial blood pressures had returned to baseline values, adrenomedullin was injected. After a 120-min recovery, 1000 µg CGRP8-37 (Sigma) was infused over 30 min into the LPA. After 20 min, although CGRP8-37 was being infused continuously, CGRP was injected into the other LPA catheter, as before, to check effective blockade of CGRP receptors by demonstrating lack of response to CGRP. Then adrenomedullin was injected again.
Protocol 4: Inhibition of cyclooxygenase pathway by indomethacin (Fig. 1D). After baseline measurements, adrenomedullin was injected. After a 120-min recovery, a total of 5 mg indomethacin (Sigma) were injected into the superior vena cava as a slow bolus (1 mg over 10 min) followed by an infusion (4 mg over 90 min), a protocol that previously has been shown to effectively block prostaglandin production (25,26). At the end of the indomethacin infusion, adrenomedullin was injected again.
Hemodynamic measurements. LPA blood flow was measured from the ultrasonic flow transducers with a flow meter (Transonic Systems, Ithaca, NY). Vascular catheters were connected to Statham P23Db strain-gauge transducers (Statham Instruments, Oxnard, CA), and mean systemic and pulmonary arterial blood pressures were measured. Heart rate was obtained by a cardiotachometer triggered from the phasic pulmonary arterial pressure tracing. All variables were recorded continuously on a direct writing polygraph (Gould, Inc., Cleveland, OH). Blood samples were obtained from the carotid artery for determination of pH, PCO2, and PO2 (Corning 158 Blood Gas Analyzer, Corning Medical, Medfield, MA) and O2 saturation and Hb concentration (OSM2 hemoximeter, Radiometer, Copenhagen, Denmark).
Data collection and analysis. For every injection of adrenomedullin, hemodynamic data were acquired at two points: before injection (baseline value) and at the point of maximal change (maximal change). Hemodynamic response was calculated: (maximal change) - (baseline value). These three values (baseline, maximal change, hemodynamic response) were compared statistically before and after the blocker agent by a paired sample t test. To assess the effect of blocker agents, the response in LPA blood flow induced by the agonists (acetylcholine, CGRP, cromakalim) was statistically compared before and after administration of the blocker (except in protocol 4) by a paired sample t test, and expressed as inhibition rate: [(response before blocker) - (response after blocker)]/(response before blocker) × 100. We considered statistical significance present when p < 0.05. Data are presented as mean ± 1 SD except for those presented in a box plot.
RESULTS
Fetal blood gases before the experiments are summarized in Table 1. Hb, O2 saturation, and PO2 were moderately low in some fetuses, but did not interfere with hemodynamic status. In all fetuses, pH was within the normal range. Fetal body weight ranged from 2200 to 4200 g (median 3250 g) at autopsy.
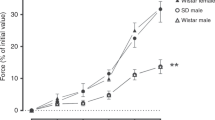
Protocol 1: Effects of NO synthesis blockade by LNA. Changes in LPA blood flow are shown in Figure 1A. In the six fetuses, LNA infusion significantly blocked the acetylcholine-induced increase in LPA blood flow by 96% (Table 2); however, one fetus needed an additional 60 min of LNA infusion for effective blockade. The adrenomedullin-induced increase in LPA blood flow also was almost completely inhibited (99%) (Fig. 2). The adrenomedullin-induced decrease in mean pulmonary and carotid arterial blood pressures was also inhibited significantly (Table 3).

Responses to adrenomedullin in LPA blood flow before and after each blocker agent. Box plot showing the 10th, 25th, 50th (median), 75th, and 90th percentiles. Values above the 90th and below the 10th percentile are shown as circles. Open bar: before blocker agent; filled bar: after blocker agent. *p < 0:05, **p < 0.01.
Protocol 2: Effects of KATP channel blockade by glibenclamide. Changes in LPA blood flow are shown in Figure 1B. In the six fetuses, glibenclamide infusion significantly blocked the cromakalim-induced increase in LPA blood flow by 93% (Table 2). The adrenomedullin-induced increase in LPA blood flow was significantly attenuated (44%) (Fig. 2). There was no significant change in mean systemic or pulmonary arterial blood pressures before and after glibenclamide infusion (Table 4). Acetylcholine increased LPA blood flow from 28 ± 27 to 200 ± 70 mL/min before glibenclamide infusion, and from 24 ± 45 to 140 ± 92 mL/min after the infusion (27% inhibition rate); these increases were not significantly different.
Protocol 3: Effects of CGRP receptor blockade by CGRP8-37. Changes in LPA blood flow are shown in Figure 1C. In the six fetuses, infusion of CGRP8-37 significantly blockade the CGRP-induced increase in LPA blood flow by 97% (Table 2). However, there were no changes in the adrenomedullin-induced increase in LPA blood flow or in the other variables (Fig. 2 and Table 5).
Protocol 4: Effects of blockade in cyclooxygenase pathway by indomethacin. Changes in LPA blood flow are shown in Figure 1D. Indomethacin infusion produced no significant change in the adrenomedullin-induced increase in LPA blood flow (Fig. 2) or in the other variables (Table 6).
DISCUSSION
In this study, we examined how the adrenomedullin-induced increase in pulmonary blood flow in fetal sheep was affected by four blocker agents: NO synthase blocker (LNA), KATP channel blocker (glibenclamide), CGRP receptor blocker (CGRP8-37), and cyclooxygenase inhibitor (indomethacin). In all four protocols, adrenomedullin decreased the main pulmonary arterial pressure and increased the left pulmonary arterial blood flow before blocker infusion, in a pattern similar to that in our previous study (10). The results in this study suggest that the adrenomedullin-induced increase in pulmonary arterial blood flow depends largely on NO release and partly on KATP channel activation, and does not involve CGRP receptors or cyclooxygenase-mediated mechanisms.
The adrenomedullin-induced increase in fetal LPA blood flow was inhibited by 99% after infusion of LNA, the NO synthase blocker, and the changes in pulmonary and systemic arterial blood pressures were also inhibited significantly (Table 3 and Fig. 2). These results indicate the existence of a NO-mediated mechanism of adrenomedullin-induced vasodilation in the fetal pulmonary circulation. Adrenomedullin originally was found by monitoring the increase of cAMP generation in platelet cells stimulated by a novel peptide (adrenomedullin) extracted from human pheochromocytoma tissues (8). Accordingly, subsequent studies demonstrated the accumulation of cAMP in rat vascular smooth muscle cells after adrenomedullin stimulation, and suggested a cAMP-mediated mechanism for the adrenomedullin-induced vasodilatory action (11,12,14,18). However, several studies have demonstrated that some arginine analogues potentially inhibit the adrenomedullin-induced vasodilation in rat renal and hindquarter vessels, suggesting a NO- and endothelium-mediated mechanism (13,14).
We previously have shown the role of KATP channels and NO release in the fetal pulmonary circulation (27). The adrenomedullin-induced increase in fetal LPA blood flow was significantly attenuated by the infusion of glibenclamide, the KATP channel blocker (Table 4 and Fig. 2). This indicates the existence of a KATP channel-mediated mechanism of adrenomedullin-induced vasodilation in the fetal pulmonary circulation. However, unlike the almost complete inhibition (99%) produced by the LNA infusion, glibenclamide infusion inhibited only 44% of the total response induced by adrenomedullin (Fig. 2). We could not explain the mechanism of this partial inhibition by glibenclamide, but a recent in vivo study in dog coronary arteries suggests a complex mechanism of adrenomedullin-induced action via KATP channels, with a pharmacologic linkage to adenosine receptors (15). Further analysis is needed to elucidate the mechanism of the adrenomedullin-induced action via KATP channels. In contrast to the significant attenuation of the adrenomedullin-induced response, glibenclamide infusion did not significantly affect the acetylcholine-induced increase in pulmonary blood flow, which is mediated by receptor-mediated NO release. This suggested that, although our previous data have shown that KATP channels act partially through NO release in the fetal pulmonary circulation, receptor-mediated NO release is independent of KATP channels. We speculate that the NO-mediated mechanism is predominant in the fetal pulmonary circulation, and that even when KATP channels are inhibited, adrenomedullin might act through receptor-activated NO-mediated mechanism. Interestingly, in a study using pulmonary vessels of adult rats, adrenomedullin-induced vasodilation was not inhibited by LNA methylester, an NO synthase inhibitor, nor by BWA-1433U83, a KATP channels blocker (28). The differences between these observations and our own might be explained by developmental alterations or by species differences in the mechanisms mediated by NO or KATP channels.
CGRP is a vasodilating peptide consisting of 37 amino acids (29). Adrenomedullin has a 27% structural homology to CGRP, suggesting a similar mechanism of vasodilatory action (30). In fact, in vitro studies in rats demonstrated that the two peptides share the same receptor (16,18). In addition, we recently reported that exogenously administered CGRP increases pulmonary blood flow in fetal sheep, in which the potent and prolonged flow patterns induced by CGRP were quite similar to those induced by adrenomedullin (23). However, in our study, although the CGRP-induced response was almost completely blocked (97%) by the CGRP receptor antagonist (CGRP8-37), the adrenomedullin-induced response was not inhibited (Table 5 and Fig. 2). These results indicate that CGRP receptors are not involved in the adrenomedullin-induced increase in pulmonary blood flow in fetal sheep and support the recent in vitro study that clearly demonstrated that adrenomedullin and CGRP act on different receptors (31). Previous studies give no consistent indication of how CGRP8-37 inhibits the adrenomedullin-induced vasodilatory action, suggesting different mechanisms between species and organs as well as regional variations in adrenomedullin receptor specificity (28,30). In terms of pulmonary vessels, there is only one previous report in which CGRP8-37 did not inhibit the vasodilating action of adrenomedullin in adult rats (28).
Prostaglandins, especially those of the E series or prostaglandin I2, are well known to be potent vasodilators of fetal pulmonary arteries, and endogenous prostaglandins may play important roles in the control of pulmonary vascular tone before and after birth. In this study, the adrenomedullin-induced increase in fetal pulmonary blood flow was not significantly inhibited by indomethacin, an inhibitor of prostaglandin production (Table 6 and Fig. 2). Although the contribution of prostaglandins to adrenomedullin-induced vasodilation is controversial (12,19,28), our data indicate that prostaglandin production is not involved in adrenomedullin-induced vasodilation of pulmonary arteries in fetal sheep.
This study demonstrates that the adrenomedullin-induced increase in pulmonary blood flow in fetal sheep depends largely on NO release, and partly on KATP channel activation. Interestingly, recent studies have demonstrated a role of NO release and KATP channel activation in the rapid decrease in pulmonary resistance and marked increase of pulmonary blood flow during the transition from fetal to neonatal circulation (7,32). Further study will determine whether endogenous adrenomedullin is or is not involved in this transition.
Abbreviations
- CGRP:
-
calcitonin gene-related peptide
- KATP:
-
ATP-dependent potassium
- LNA:
-
Nω-nitro-L-arginine
- LPA:
-
left pulmonary artery
- NO:
-
nitric oxide
References
Heymann MA, Soifer SJ 1989 Control of fetal and neonatal pulmonary circulation. In: Weir EK, Reeves JT (eds) Pulmonary Vascular Physiology and Pathophysiology. Dekker, New York, 33–50.
Teitel DF, Iwamoto HS, Rudolph AM 1990 Changes in the pulmonary circulation during birth-related events. Pediatr Res 27: 372–378
Leffler CW, Hessler JR, Green RS 1984 Mechanism of stimulation of pulmonary prostacyclin synthesis at birth. Prostaglandins 28: 877–887
Konduri GG, Gervasio CT, Theodorou AA 1993 Role of adenosine triphosphate and adenosine in oxygen-induced pulmonary vasodilation in fetal lambs. Pediatr Res 33: 533–539
Heymann MA, Rudolph AM, Nies AS, Melmon KL 1969 Bradykinin production associated with oxygenation of the fetal lamb. Circ Res 25: 521–534
Tiktinsky MH, Cummings JJ, Morin FC III 1992 Acetylcholine increases pulmonary blood flow in intact fetuses via endothelium-dependent vasodilation. Am J Physiol 262: H406–H410
Abman SH, Chatfield BA, Hall SL, McMurtry IF 1990 Role of endothelium-derived relaxing factor during transition of pulmonary circulation at birth. Am J Physiol 259: H1921–H1927
Kitamura K, Kangawa K, Kawamoto M, Ichiki Y, Nakamura S, Matsuo H, Eto T 1993 Adrenomedullin: A novel hypotensive peptide isolated from human pheochromocytoma. Biochem Biophys Res Commun 192: 553–560
Sugo S, Minamino N, Kangawa K, Miyamoto K, Kitamura K, Sakata J, Eto T, Matsuo H 1994 Endothelial cells actively synthesize and secrete adrenomedullin. Biochem Biophys Res Commun 201: 1160–1166
de Vroomen M, Takahashi Y, Gournay V, Roman C, Rudolph AM, Heymann MA 1997 Adrenomedullin increases pulmonary blood flow in fetal sheep. Pediatr Res 41: 493–497
Ishizaka Y, Ishizaka Y, Tanaka M, Kitamura K, Kangawa K, Minamino N, Matsuo H, Eto T 1994 Adrenomedullin stimulates cyclic AMP formation in rat vascular smooth muscle cells. Biochem Biophys Res Commun 200: 642–646
Eguchi S, Hirata Y, Kano H, Sato K, Watanabe Y, Watanabe TX, Nakajima K, Sakakibara S, Marumo F 1994 Specific receptors for adrenomedullin in cultured rat vascular smooth muscle cells. FEBS Lett 340: 226–230
Feng CJ, Kang B, Kaye AD, Kadowitz PJ, Nossaman BD 1994 L-NAME modulates responses to adrenomedullin in the hindquarters vascular bed of the rat. Life Sci 55: PL433–PL438
Hirata Y, Hayakawa H, Suzuki E, Ikenouchi H, Kohmoto O, Kimura K, Kitamura K, Eto T, Kangawa K, Matsuo H, Omata M 1995 Mechanisms of adrenomedullin-induced vasodilation in the rat kidney. Hypertension 25: 790–795
Sabates BL, Pigott JD, Choe EU, Cruz MP, Lippton HL, Hyman AL, Flint LM, Ferrara JJ 1997 Adrenomedullin mediates coronary vasodilation through adenosine receptors and KATP channels. J Surg Res 67: 163–168
Takahashi H, Watanabe TX, Nishimura M, Nakanishi T, Sakamoto M, Yoshimura M, Komiyama Y, Masuda M, Murakami T 1994 Centrally induced vasopressor and sympathetic responses to a novel endogenous peptide, adrenomedullin, in anesthetized rats. Am J Hypertension 7: 478–482
Entzeroth M, Doods HN, Wieland HA, Wienen W 1995 Adrenomedullin mediates vasodilation via CGRP1 receptors. Life Sci 56: PL19–PL25
Osajima A, Mutoh Y, Uezono Y, Kawamura M, Izumi F, Takasugi M, Kuroiwa A 1995 Adrenomedullin increases cyclic AMP more potently than CGRP and amylin in rat renal tubular basolateral membranes. Life Sci 57: 457–462
Jougasaki M, Aarhus LL, Heublein DM, Sandberg SM, Burnett JC Jr 1997 Role of prostaglandins and renal nerves in the renal actions of adrenomedullin. Am J Physiol 272: F260–F266
Nossaman BD, Feng CJ, Kaye AD, DeWitt B, Coy DH, Murphy WA, Kadowitz PJ 1996 Pulmonary vasodilator responses to adrenomedullin are reduced by NOS inhibitors in rats but not in cats. Am J Physiol 270: L782–L789
Rudolph AM, Heymann MA, Fishman N, Lakier JB 1975 Formalin infiltration of the ductus arteriosus: a method for palliation of infants with selected congenital cardiac lesions. N Engl J Med 292: 1263–1268
Nelson MT, Quayle JM 1995 Physiological roles and properties of potassium channels in arterial smooth muscle. Am J Physiol 268: C799–C822
de Vroomen M, Takahashi Y, Roman C, Heymann MA 1998 Calcitonin gene-related peptide increases pulmonary blood flow in fetal sheep. Am J Physiol 274: H277–H282
Brain SD, Williams TJ, Tippins JR, Morris HR, MacIntyre I 1985 Calcitonin gene-related peptide is a potent vasodilator. Nature (Lond) 313: 54–56
Frantz E, Soifer SJ, Clyman RI, Heymann MA 1989 Bradykinin produces pulmonary vasodilation in fetal lambs: The role of prostaglandin production. J Appl Physiol 67: 1512–1517
Morin FC III, Egan EA, Norfleet WT 1988 Indomethacin does not diminish the pulmonary vascular response of the fetus to increased oxygen tension. Pediatr Res 24: 696–700
Chang J-K, Moore P, Fineman JR, Soifer SJ, Heymann MA 1992 K+ channel pulmonary vasodilation in fetal lambs: Role of endothelial derived nitric oxide. J Appl Physiol 73: 188–194
Heaton J, Lin B, Chang J-K, Steinberg S, Hyman A, Lippton H 1995 Pulmonary vasodilation to adrenomedullin: A novel peptide in humans. Am J Physiol 268: H2211–H2215
Amara SG, Jonas V, Rosenfeld MG, Ong ES, Evans RM 1982 Alternative RNA processing in calcitonin gene expression generates mRNAs encoding different polypeptide products. Nature 298: 240–244
Muff R, Born W, Fischer JA 1995 Calcitonin, calcitonin gene-related peptide, adrenomedullin and amylin: Homologous peptides, separate receptors and overlapping biological actions. Eur J Endocrinol 133: 17–20
Champion HC, Santiago JA, Murphy WA, Coy DH, Kadowitz PJ 1997 Adrenomedullin-(22-52) antagonizes vasodilator responses to CGRP but not adrenomedullin in the cat. Am J Physiol 272: R234–R242
Tristani-Firouzi M, Martin EB, Tolarova S, Weir EK, Archer SL, Cornfield DN 1996 Ventilation-induced pulmonary vasodilation at birth is modulated by potassium channel activity. Am J Physiol 271: H2353–H2359
Acknowledgements
The authors thank Julien I. E. Hoffman, M.D. for statistical consultation and Ronald I. Clyman, M.D. for important additional data concerning dosages and serum concentrations of indomethacin. We also acknowledge the expertise and assistance of Mario Trujillo and Bruce D. Payne.
Author information
Authors and Affiliations
Additional information
Supported by United States Public Health Service Grant HL40473. The data in this paper were partly presented at the Pediatric Academic Societies' Annual Meeting held in Washington, DC in 1997.
Rights and permissions
About this article
Cite this article
Takahashi, Y., De Vroomen, M., Gournay, V. et al. Mechanisms of Adrenomedullin-Induced Increase of Pulmonary Blood Flow in Fetal Sheep. Pediatr Res 45, 276–281 (1999). https://doi.org/10.1203/00006450-199902000-00020
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1203/00006450-199902000-00020
