
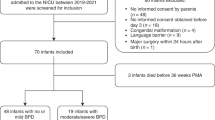
Abstract
Nestin is an intermediate filament protein found in CNS progenitor cells. Nestin reappears in CNS tumor cells and reactive astrocytes after CNS injury. In this study we investigated whether nestin could be detected in the cerebrospinal fluid (CSF) of newborn infants and whether expression levels change with gestational age (GA) and/or brain injury. Using Western blot analysis, we examined the expression of nestin in the CSF of newborn infants(GA 25-42 wk) with asphyxia (n = 14), periventricular leukomalacia and peri(intra)ventricular hemorrhage (n = 7), and in a control group (n = 11). Protein extract from the periventricular brain tissue of a 1-wk-old infant was also analyzed. Nestin was detected in all the CSF samples and in the protein extract from the periventricular brain tissue. Although the CSF levels of nestin expression did not change with increasing GA, the asphyxia group had significantly lower levels of nestin in the CSF. An unexpected finding was that brain-derived nestin had an apparent molecular mass of approximately 240 kD, whereas all analyzed CSF samples contained two nestin-immunoreactive proteins at 200 and 220 kD. Experimental deglycosylation of the 240-kD form reduced the molecular mass to 220 kD, indicating that nestin undergoes a specific deglycosylation upon release into the CSF.
Similar content being viewed by others

Main
Neurons and glial cells of the adult CNS are derived from proliferating and differentiating neuroepithelial progenitor cells of the neural tube. The classical view holds that this proliferation is transient and that the mature CNS is devoid of stem cells. This attitude has, however, been challenged recently by the finding that the brain of adult mice contains a small stem cell population in the subependymal layer of the lateral ventricles(1, 2). The stem cells migrate from the subventricular zone and differentiate into neurons in the olfactory bulb(3). Furthermore, stem cells removed from the striatum and subventricular zone of an adult mouse and grown in primary culture have the capacity to generate more stem cells or give rise to neurons and astrocytes, depending on the culture conditions(4, 5). The identification of a corresponding stem cell population in the human brain is clearly of interest and may be facilitated by the fact that CNS progenitor and stem cells express the protein nestin.
Nestin is a recently discovered IF related to neurofilaments and glial fibrillary acidic protein, which are found in neurons and astrocytes, respectively. Nestin is predominantly expressed in the dividing neuroepithelial cells of the newly closed neural tube and has been extensively used to identify the population of CNS progenitor cells in rodents(6–10). In addition, it has been shown that the stem cells of the adult CNS also express nestin(1, 4). The expression of nestin is up-regulated in response to situations of cellular stress and/or induced proliferation in the nervous system. Thus, nestin is reexpressed in CNS tumors(11, 12) and in immortalized cell lines derived from cells of early CNS(13). Rat and mouse nestin is also up-regulated after brain injury(14, 15), nervus opticus damage, and lesions of the dorsal funiculus in the spinal cord(16). Regulatory regions responsible for the expression of nestin in the progenitor cells of the early CNS have been analyzed in transgenic mice(17), and the same regions appear to control the reexpression of nestin in response to CNS injury(15, 16). Little is yet known about the expression of nestin during human fetal and postnatal development, but it has been demonstrated that cells in the germinal matrix of the human fetus at 17 and 20 wk GA and some morphologically immature subependymal cells at 40 wk GA express nestin(12).
In this study we investigated whether nestin could be detected in the CSF of newborn infants and if levels of nestin changed with GA or in case of perinatal brain injury. Using Western blot analysis, we examined the expression of nestin in the CSF of newborn infants with asphyxia, PVL, and PIVH and in a control group. We also examined a protein extract from the periventricular brain tissue of a full-term 1-wk-old infant.
METHODS
Patients and CSF Sampling
The protocol for CSF sampling for the analysis of brain proteins was approved by the local ethics committee of the Karolinska Hospital. Parental consent was obtained in all cases. Thirty-two newborn infants were divided into three groups.
Asphyxia group. Fourteen infants fulfilling the following criteria of neonatal asphyxia were included into this group: 1) infants with intrapartum distress as indicated cardiotochographically (late decelerations for >1 h or severe abnormalities as absent variability or persistent bradycardia for >30 min before birth), early passage of thick meconium, or scalp pH < 7.2 immediately before birth, and 2) infants who needed neonatal resuscitation with positive pressure ventilation for >3 min, Apgar score at 5 min <6, or umbilical arterial/first postnatal pH < 7.1, and/or base excess < -10. At the time of CSF sampling, these infants had symptoms of hypoxicischemic encephalopathy according to Sarnat and Sarnat(18).
PVL and PIVH group. Seven infants who suffered from PVL and PIVH (diagnosed by cranial ultrasonography) were included in this subgroup.
Control group. Eleven infants with clinical signs of systemic infections necessitating lumbar puncture were used as controls. In these cases the routine CSF cell counts were normal, and bacterial/viral cultures were negative.
CSF was collected by spinal tap in the L3-4 region. After 1.5 mL of CSF necessary for clinical purposes had been tapped, 0.5 mL was collected, and the sample was frozen (-70 °C) for the assay.
Brain tissue was obtained from the periventricular region of a full-term 1-wk-old infant, who had no brain injury but who died due to congenital heart disease. The tissue was frozen in liquid nitrogen and stored at -70 °C.
Generation of the Polyclonal Antiserum 4350 against Human Nestin Protein
A region from the fourth exon, corresponding to the amino acid 1454-1618 in the human nestin protein(19), was amplified by PCR under the following conditions: 20 cycles at 94 °C for 1 min, 50 °C for 1 min, and 72 °C for 1 min. Two primers, 5′-GAA TTC CCT GGA GTC TGA TTC TGT AAG-3′ and 5′-ATT GCG AAT TCC CTG GAG TCT GAT TCT GTA AG-3′, and 100 ng of a genomic human nestin clone(19) were used. The resulting 462-bp PCR fragment was cloned into the TA vector (Invitrogen), sequenced to verify that no mutations were introduced in the PCR, and excised with the restriction enzymesEco RI and HindIII (Biolabs). The resulting fragment was subcloned into the pEZZ18 vector, and a fusion protein between the Z repeat of Staphylococcus aureus protein A and nestin was then produced from the plasmid in Escherichia coli as previously described(20) and used to immunize rabbits. The rabbits were reimmunized at 4 wk after the first immunization, followed by terminal bleeding 12 wk later. The new antiserum was called 4350.
Cell Culture
The human embryonic muscle cell line G6(21), previously shown to express nestin(22), was grown in Ham's nutrient mixture F10 (Sigma Chemical Co., St.Louis, MO) with 15% FCS. The cell line ST15A, immortalized from rat cerebellum(23) and also expressing nestin, was grown in Dulbecco's modified Eagle's medium (Life Technologies, Inc., Gaithersburg, MD) with 10% FCS.
Protein Preparation and Deglycolysation Assays
The brain tissue was homogenized with an immersible ultrasound homogenizator in ice-cold buffer containing: 0.1 M NaCl, 0.01 M Tris-Cl (pH 7.6), 0.001 M EDTA (pH 7.8), 1 μg/mL aprotinin, pepstatin A, leupeptin, and 100 μg/mL phenylmethylsulfonyl fluoride. Cultured cells were scraped off the culture dish and lysed in 1% SDS. The extracts were centrifuged at 8000× g to remove cellular debris. Total protein content in the extracts was determined by the Bio-Rad protein assay. The glycosylation status of nestin was analyzed by removing sugar groups. Aliquots of the protein extract (30 μL, total protein content 1 μg/μl) from the cell line G6 were incubated with the following enzymes: N-glycosidase F (0.4 U), neuraminidase (2 mU), and O-glycosidase (2.5 mU) (Boehringer Mannheim) for 20 h at 37 °C. The enzymes were omitted in the control samples. The protein extracts were examined using Western blot analysis (see below). Removal of the sugar groups in the protein samples was controlled with digoxigenin glycan/protein double labeling kit (Boehringer Mannheim).
Western Blot and Immunocytochemistry
Thirty to 100 μg of total protein in sample buffer (15% SDS, 0.31 M Tris-HCl, pH 6.8-6.9, 25% glycerol, 1.5 mM β-mercaptoethanol, 0.05% bromphenol blue) were electrophoresed in 6% SDS-polyacrylamide gel together with protein molecular weight markers (Amersham Corp.) at 160 V for 6 h. After electrophoresis the proteins were blotted onto nitrocellulose membrane(Schleicher & Schuell) for 16 h at 0.35 A. Membranes were blocked with 5% nonfat dried milk and incubated with anti-nestin antisera 129 (11), 130 (11), and 4350. The following dilutions were used: 129 at 1:1000, and 130 and 4350 at 1:2000. Bound primary antibodies were visualized using secondary goat-anti-rabbit antibodies (dilution 1:2000) conjugated with horseradish peroxidase (the Binding Site) and a chemiluminescence kit (ECL, Amersham Corp.). Three different protein concentrations were tested in each case to verify that the signal analyzed by densitometric scanning was in the linear range. The films were scanned using the Ofoto program, and nestin amounts were evaluated with the Image 1.52 program. Immunocytochemistry was carried out on G6 and ST15A cells, which were grown on coverslips. The cells were fixed for 8 min in 4% paraformaldehyde in PBS, then rinsed in PBS and blocked with 3% BSA in PBS for 30 min. Incubation with the primary antibody (4350 at 1:1000 or 130 at 1:1000) was performed overnight. After thoroughly rinsing in PBS, the cells were incubated with secondary anti-rabbit antibodies conjugated with rhodamine(Dakopatts), rinsed in PBS again, and photographed.
Statistics
The Spearman rank correlation test was applied to determine correlation between the levels of nestin in the CSF and GA of the control newborn infants. Nonparametrical Mann-Whitney rank sum test was used to compare the levels of nestin in the CSF of PVL and PIVH group, asphyxia group, and age-matched control newborn infants. Newborn infants younger than 35 wk of GA were age-matched controls to the PVL and PIVH group, and newborn infants older than 35 wk of GA were age-matched controls to the asphyxia group.
RESULTS
Analysis of nestin immunoreactivity in newborn brain and CSF. We first wanted to determine whether nestin-immunoreactive protein could be detected in the CSF of newborn infants and/or in the protein extract from the periventricular brain tissue. Extracts from the nestin-producing cell line G6 were run in parallel as a positive control. Western blot analysis, using two antisera against rat nestin, 129 and 130, revealed that the protein extracts from the periventricular brain tissue, and the cell line G6 showed a nestin band of approximately 240 kD, whereas two bands at 200 and 220 kD were reproducibly found in the CSF (Fig. 1).

Western blot analysis of the protein extracts from(A) brain tissue, (B) the cell line G6, and (C) CSF from a control newborn, analyzed with the 130 anti-nestin antiserum. The migration of proteins with known molecular masses is indicated to the left.
Generation and analysis of the new antiserum 4350 against human nestin. To investigate the unexpected size difference between brain- and CSF-derived nestin further, and to rule out that it was the result of a nonspecific immunoreaction to a protein in the CSF, we generated a new antiserum against human nestin. We subcloned a region of the human nestin gene encoding a portion of the long C-terminal tail region of the nestin protein. This region was selected to minimize the risk for cross-reactivity, because it lies outside the region encoding the α-helical domain (also referred to as the rod domain), which is conserved among the various IF proteins(Fig. 2) (see Heins and Aebi(24) for review on IF structure). The nestin subclone was introduced into the pEZZ18 vector(20), and a fusion protein between S. aureus protein A and the C-terminal region of human nestin was produced and used to immunize rabbits (Fig. 2).

Generation of the 4350 antiserum. In the top section a schematic representation of the human nestin gene is shown. The region encoding the α-helical domain is shown in black. A region encoding a part of the C-terminal region (gray) was subcloned into the pEZZ18 vector(20), from which a fusion protein, consisting of nestin and Z repeats of protein A, was generated and used to immunize rabbits.
The resulting antiserum 4350 was first tested on cells from the G6 cell line. The immunocytochemical staining revealed the expected cytoskeletal pattern, with a strong perinuclear component and a threadlike cytoplasmic network typical for IF proteins (Fig. 3a). IF-like immunostaining, although not as intense as in G6, was also observed in the mouse cell line ST15A (data not shown). We next tested the 4350 antiserum in Western blots with proteins from human brain, the cell line G6, and the CSF used above. The same pattern was obtained as when 129 and 130 antisera were used (Fig. 3b, compare with Fig. 1). These data strongly indicate that antiserum 4350 correctly identifies human nestin protein and that the size difference between brain- and CSF-derived nestin is not an artefact.

(a) Nestin distribution in G6 cells after immunocytochemistry with the 4350 antiserum. Note the presence of cells with intense perinuclear staining and a threadlike cytoplasmic staining typical for IF. (b) Western blot analysis of the protein extracts from(A) brain tissue, (B) cell line G6, and (C) CSF from a control newborn, analyzed with 4350 anti-nestin antiserum. The migration of proteins with known molecular masses is indicated to the left.
Analysis of the glycosylation status of the nestin protein. The predicted molecular mass of the human nestin protein deduced from the coding sequence is 200 kD(19). The migration of brain- and cell line-derived nestin as a 240-kD protein suggested that posttranslational modification might occur causing an increase in the molecular mass of the protein. We supposed that a cause of the different apparent molecular masses may be a change in the glycosylation pattern and therefore analyzed the glycosylation status of the 240-kD nestin form. After enzymatic deglycosylation in vitro, the apparent molecular mass of nestin derived from the cell line G6 decreased from 240 to 220 kD(Fig. 4, lane A). In the control sample, where the enzymes were omitted, nestin migrated as a 240-kD protein (Fig. 4, lane B).

The glycosylation status of nestin. Western blot analysis, using the 4350 antiserum, of protein extract from the cell line G6(A) after deglycosylation with a mixture of enzymes (see“Methods”) and (B) after omitting the deglycosylating enzymes. The migration of proteins with known molecular masses is indicated to the left.
Analysis of the CSF samples from newborn infants. We next examined whether the levels of the nestin protein change with GA or in response to perinatal brain injury. We analyzed CSF from newborn infants without proven CNS pathology (n = 11), from full-term newborn infants with asphyxia (n = 14), and from preterm newborn infants with PVL and PIVH (n = 7). The GA of the newborn infants ranged from 25 to 42 wk. The results from two patients of each group are displayed inFigure 5a. The same pattern i.e. two nestin bands of 220 and 200 kD, is observed in all cases. In Figure 5b the relative nestin levels of 32 cases are shown. There was no statistically significant correlation between the levels of nestin in the CSF and GA of control newborn infants (Spearman rank correlation test). In contrast, the asphyxia group had significantly lower levels of nestin in the CSF, when compared with age-matched control newborn infants (>35 wk of GA)(p < 0.05). However, the PVL and PIVH group did not show a significant difference of nestin levels in the CSF when compared with age-matched control newborns (<35 wk of GA) (Mann-Whitney rank sum test).

(a) Western blot analysis of (A andB) CSF of two control newborn infants; (C and D) CSF of newborn infants with PVL and PIVH; (E anf F) CSF of newborn infants with asphyxia. The migration of proteins with known molecular masses is indicated to the left. (b) Relative nestin protein values of control newborn infants (open circles), newborn infants with PVL and PIVH (open diamonds), and asphyxiated newborn infants(filled diamonds).
DISCUSSION
In this report we show that nestin protein is present in the CSF of newborn infants with asphyxia, PVL, and PIVH, as well as in the CSF of control infants without signs of CNS injury. The levels of nestin in the CSF decrease in asphyxiated newborn infants. CSF-derived nestin appears as a lighter protein than brain-derived nestin, and we show that deglycosylation can mimic the transition from the heavy to the light form of nestin.
We also detected nestin protein in the periventricular brain tissue of a 1-wk-old full-term infant. This finding corroborates previous observations of nestin expression in rodent CNS progenitor cells located in the neuroepithelium lining the neural tube(6, 7, 10). Furthermore, a small population of nestin-positive CNS stem cells is present in the subependyma of adult mice(1), and a small number of nestin-positive cells was also found in the subependyma of a 40-wk GA human fetus(12). Because these nestin-positive cells are juxtaposed to the ventricles, it seems reasonable to assume that nestin in the CSF, at least in part, comes from the nestin-positive CNS progenitor cells lining the ventricles, and it is possible that some of these cells proliferate and undergo cell death, which has been previously shown in the mouse(2). When these cells are damaged or die, they disintegrate and leak constituents into the extracellular compartment and subsequently into the CSF. An additional source of nestin in the CSF may be the plexus chorioideus, which produces CSF and has been shown to express nestin mRNA(10). At present, we do not know the relative contribution from CNS progenitor cells and cells from the plexus chorioideus to the pool of nestin found in the CSF.
We find here that nestin levels were significantly decreased in the CSF in the asphyxia group. This may reflect a reduced release of nestin into the CSF from cells surrounding the ventricles or, alternatively, a reduced level of nestin expression in these cells. A reduced release may be the result of the nestin-positive cells' migration away from the ventricular area toward the site of damage in response to hypoxic-ischemic injury, a situation which would actually lower the number of nestin-positive cells in the vicinity of the ventricle. This type of migratory behavior, i.e. away from the ventricular area, has been suggested to occur after mechanical CNS injury(14–16). The idea that the cellular levels of nestin protein are reduced after asphyxia is also possible, but this would be in contrast to previous reports on increased expression of nestin in CNS tumor cells(11, 12) and after mechanical CNS injury(14–16). The levels of nestin in CSF were, however, not analyzed in the cases of CNS tumors and mechanical or chemical CNS injury(11, 12, 14–16), which precludes a direct comparison. We therefore cannot establish the mechanism for the reduced levels of nestin after asphyxia, but to address the issue in further detail we are currently analyzing whether the expression of nestin is altered after experimental asphyxia in an animal model.
An unexpected finding in this study is that brain- and CSF-derived nestin have different apparent molecular masses. An approximately 240-kD form is found in the protein extract from the periventricular brain tissue and from the cell line G6, whereas two nestin bands of approximately 200 and 220 kD appear in all the analyzed CSF samples. Because removal of sugar groups reduces the apparent molecular mass from 240 to 220 kD, i.e. the same molecular mass as one of the nestin bands in the CSF, we suggest that deglycosylation at least in part explains the difference in apparent molecular masses. Thus, we assume that deglycosylation takes place when nestin is released into the CSF. We cannot at present explain the mechanism for generation of the 200-kD band. Further deglycosylation is conceivable, because 200 kD is the molecular mass deduced from the coding sequence(19). Alternatively, the shorter form may be the result of proteolytic cleavage, because proteolytic enzymes are known to reside in the CSF(25). Screening of the CSF proteins with a digoxigenin glycan/protein double labeling kit revealed that not all proteins were deglycosylated (data not shown), suggesting that only a subset of proteins, including nestin, is subject to deglycosylation. It is an interesting, although speculative, idea that the deglycosylated form of nestin could be functionally distinct from the glycosylated form and less readily forms large aggregates, which otherwise could lead to obstruction of narrow ducts of the CSF. It will be interesting to learn whether other proteins acquire different characteristics by specific deglycosylation in the CSF.
Abbreviations
- CSF:
-
cerebrospinal fluid
- IF:
-
intermediate filament
- PVL:
-
periventricular leukomalacia
- PIVH:
-
peri(intra)ventricular hemorrhage
- GA:
-
gestational age
- PCR:
-
polymerase chain reaction
References
Morshead CM, Reynolds BA, Craig CG, McBurney MW, Staines WA, Morassutti D, Weiss S, van der Kooy D 1994 Neural stem cells in the adult mammalian forebrain: a relatively quiescent subpopulation of subependymal cells. Neuron 13: 1071–1082
Morshead CM, van der Kooy D 1992 Postmitotic death is the fate of constitutively proliferating cells in the subependymal layer of the adult mouse brain. J Neurosci 12: 249–256
Lois C, Alvarez-Buylla A 1994 Long-distance neuronal migration in the adult mammalian brain. Science 264: 1145–1148
Reynolds BA, Weiss S 1992 Generation of neurons and astrocytes from isolated cells of the adult mammalian central nervous system. Science 255: 1707–1710
Lois C, Alvarez-Buylla A 1993 Proliferating subventricular zone cells in the adult mammalian forebrain can differentiate into neurons and glia. Proc Natl Acad Sci USA 90: 2074–2077
Frederiksen K, McKay RDG 1988 Proliferation and differentiation of rat neuroepithelial precursor cells in vivo. J Neurosci 8: 1144–1151
Lendahl U, Zimmerman LB, McKay RDG 1990 CNS stem cells express a new class of intermediate filament protein. Cell 60: 585–595
Fishell G, Mason CA, Hatten ME 1993 Dispersion of neural progenitors within the germinal zones of the forebrain. Nature 362: 636–638
Williams BP, Price J 1995 Evidence for multiple precursor cell types in the embryonic rat cerebral cortex. Neuron 14: 1181–1188
Dahlstrand J, Lardelli M, Lendahl U 1995 Nestin mRNR expression correlates with the central nervous system progenitor cell state in many, but not all, regions of developing central nervous system. Dev Brain Res 84: 109–129
Dahlstrand J, Collins VP, Lendahl U 1992 Expression of the class VI intermediate filament nestin in human central nervous system tumors. Cancer Res 52: 5334–5341
Tohyama T, Lee VM-Y, Rorke LB, Marvin M, McKay RDG, Trojanowsky JQ 1992 Nestin expression in embryonic human neuroepithelium and in human neuroepithelial tumor cells. Lab Invest 66: 303–313
Redies C, Lendahl U, McKay RDG 1991 Differentiation and heterogeneity in T-antigen immortalized precursor cell lines from mouse cerebellum. J Neurosci Res 30: 601–615
Clarke SR, Shetty AK, Bradley JL, Turner DA 1994 Reactive astrocytes express the embryonic intermediate neurofilament nestin. NeuroReport 5: 1885–1888
Lin RCS, Matesic DF, Marvin M, McKay RDG, Brustle O 1995 Reexpression of the intermediate filament nestin in reactive astrocytes. Neurobiol Dis 2: 79–85
Frisén J, Johansson CB, Török C, Risling M, Lendahl U 1995 Rapid, widespread and long-lasting induction of nestin contributes to the generation of glial scar tissue after CNS injury. J Cell Biol 131: 453–464
Zimmerman L, Lendahl U, Cunningham M, McKay R, Parr B, Gavin B, Mann J, Vassileva G, McMahon A 1994 Independent regulatory elements in the nestin gene direct transgene expression to neural stem cells or muscle precursors. Neuron 12: 11–24
Sarnat HB, Sarnat MS 1976 Neonatal encephalopathy following fetal distress. Arch Neurol 33: 696–705
Dahlstrand J, Zimmerman LB, McKay RDG, Lendahl U 1992 Characterization of human nestin gene reveals a close evolutionary relationship to neurofilaments. J Cell Sci 103: 589–597
Löwenadler B, Jansson B, Paleus S, Holmgren E, Nilsson B, Moks T, Palm G, Josephson S, Philipson L, Uhlén M 1987 A gene fusion system for generating antibodies against short peptides. Gene 58: 87–97
Jin P, Farmer K, Ringertz N, Sejersen T 1993 Proliferation and differentiation of human fetal myoblasts is regulated by PDGF-BB. Differentiation 54: 47–54
Sejersen T, Lendahl U 1993 Transient expression of the intermediate filament nestin during skeletal muscle development. J Cell Sci 106: 1291–1300
Frederiksen K, Jat PS, Valtz N, Levy D, McKay R 1988 Immortalisation of precursor cells from the mammalian CNS. Neuron 1: 439–448
Heins S, Aebi U 1994 Making heads and tails of intermediate filament assembly, dynamics and networks. Curr Opin Cell Biol 6: 25–33
Nyberg F, Nordström K, Terenius L 1985 Endopeptidase in human cerebrospinal fluid which cleaves proenkephalin B opioid peptides at consecutive basic amino acids. Biochem Biophys Res Commun 131: 1069–1074
Acknowledgements
The authors thank Dr. Björn Löwenadler for generating the anti-nestin antiserum 4350, Hilary Brown for advise on Western blot analysis, Dr. Ron McKay and Dr. Tomas Sejersen for cells, Dr. Bronius Jurevicius for providing us with human brain tissue, and Dr. Giovanna Marchini for valuable comments on the manuscript.
Author information
Authors and Affiliations
Additional information
Supported by grants from Sahléns Stiftelse, the Swedish Medical Research Council 5234 (H.I.,) and 7164 (G.F.), the Swedish Cancer Society, Margaret och Axel Ax:son Johnsons Stiftelse, Kjell och Märta Beijers Stiftelse, Åke Wibergs Stiftelse and Magn. Bergvalls Stiftelse (U.L.), and ESPR Young Investigator Exchange Program (G.G.).
Rights and permissions
About this article
Cite this article
Grigelioniené, G., Blennow, M., Török, C. et al. Cerebrospinal Fluid of Newborn Infants Contains a Deglycosylated Form of the Intermediate Filament Nestin. Pediatr Res 40, 809–814 (1996). https://doi.org/10.1203/00006450-199612000-00006
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1203/00006450-199612000-00006
