
Abstract
Salmonella infections can become chronic and increase the risk of cancer. The mechanisms by which specific Salmonella organisms contribute to cancer, however, are still unknown. Live and attenuated Salmonella are used as vectors to target cancer cells, but there have been no systematic studies of the oncogenic potential of chronic Salmonella infections in cancer models. AvrA, a pathogenic product of Salmonella, is inserted into host cells during infection and influences eukaryotic cell pathways. In the current study, we colonized mice with Salmonella AvrA-sufficient or AvrA-deficient Salmonella typhimirium strains and induced inflammation-associated colon cancer by azoxymethane/dextran sulfate sodium (AOM/DSS). We confirmed Salmonella persisted in the colon for up to 45 weeks. Salmonella was identified not only in epithelial cells on the colonic luminal surface and base of the crypts but also in invading tumors. Tumor incidence in the AvrA+infected group was 100% compared with 51.4% in the AOM/DSS group without bacterial gavage and 56.3% in mice infected with the AvrA- strain. Infection with AvrA+ strain also altered tumor distribution from the distal to proximal colon that might reflect changes in the microbiome. AvrA-expressing bacteria also upregulated beta-catenin signaling as assessed by decreased beta-catenin ubiquitination, increased nuclear beta-catenin and increased phosphorylated-beta-catenin (Ser552), a marker of proliferating stem-progenitor cells. Other β-catenin targets increased by AvrA included Bmi1, a cancer stem cell marker, matrix metalloproteinase-7, and cyclin D1. In summary, AvrA-expressing Salmonella infection activates β-catenin signals and enhances colonic tumorigenesis. Our findings provide important new mechanistic insights into how a bacterial protein targets proliferating stem-progenitor cells and contributes to cancer development. Our observations also raise a note of caution regarding the use of mutant Salmonella organisms as vectors for anti-cancer therapy. Finally, these studies could suggest biomarkers (such as AvrA level in gut) to assess cancer risk in susceptible individuals and infection-related dysregulation of β-catenin signaling in cancer.
Similar content being viewed by others


Introduction
Colon cancer is a major cause of cancer-related deaths in the Western world. Increasing evidence has implicated chronic inflammation in its causation, and recently attention has focused on the role of the microbiome. Chronic bacterial infection and the ensuing colonic inflammation contribute to tumor initiation and tumor progression.1, 2 The number of microbial cells comprising the commensal microflora is 10 times larger than the number of eukaryotic cells in the human body. More than 1000 species of bacteria inhabit the human colon. Because of the complexity of the gut flora, identification of the specific microbial organisms or bacterial products contributing to colon cancer remains challenging. Deciphering the roles of specific bacterial products is in its infancy. Progress has been made, however. For example, a recent study demonstrated that a human colonic commensal bacterium promoted tumorigenesis via induction of interleukin-17 T helper cell responses.3 In mice, colitis can promote tumorigenesis by altering microbial composition and inducing the expansion of microorganisms with genotoxic capabilities.4 Recent studies suggested that polyketide synthase is an oncogenic factor in that genetic deletion of this element in Escherichia coli NC101 decreased azoxymethane (AOM)-induced tumor multiplicity and invasion in interleukin-10 knockout mice.4
Salmonella infection and colonization in humans can be chronic2 and increase the risk of other gastrointestinal diseases, including chronic cholecystitis and gallbladder cancer.5, 6 Whereas increasing evidence links some Salmonella species to carcinogenesis, other non-invasive Salmonella strains appear promising as vectors to specifically target cancer cells.7 Salmonella and its derivatives invade tumor cells more efficiently than normal cells in animal models.8, 9 The safety of Salmonella as an anti-cancer vector is uncertain, however, because there have been no systematic studies of the effects of chronic infections of non-pathogenic or attenuated Salmonella strains in cancer models.
Bacteria can modulate their eukaryotic host by secreting microbial proteins that modulate host cell function. AvrA is a multifunctional pathogenic gene of enteric bacteria whose encoded protein is inserted into host cells. Our own studies, as well as those of other investigators, have demonstrated that AvrA influences eukaryotic cell pathways by altering ubiquitination and acetylation of target proteins to modulate inflammation, epithelial apoptosis and proliferation.10, 11, 12, 13, 14, 15, 16 For example, AvrA acts as a deubiquintinase to suppress β-catenin ubiquitination thereby stabilizing β-catenin and inhibiting its degradation.15 This results in increased β-catenin signaling, which enhances intestinal epithelial cell proliferation.15
In the current study, our goal was to elucidate the role of AvrA on colon cancer development. We hypothesized that the bacterial effector AvrA would activate the β-catenin pathways to promote colonic tumorigenesis. Other investigators have taken advantage of the colitis induced by Salmonella to study early acute infections or late chronic infections and associated inflammation.17 In previous studies, we have established a mouse model with persistent of Salmonella AvrA+ or AvrA− infection in the gastrointestinal tract.18 In the current study, using this model, we examined the effects of chronic infection with AvrA-expressing bacteria on inflammation-associated colon cancer. We initiated colonic epithelial cell mutations with AOM and accelerated tumorigenesis with dextran sulfate sodium (DSS) treatment. To determine the effects of AvrA, we infected mice with Salmonella strains that differed in AvrA expression level. We used AvrA (parental PhoPC) or AvrA deficient strains (AvrA−) or PhoPC AvrA− strain complemented with a plasmid encoding AvrA (PhoPC AvrA−/AvrA+), as we previously described.13, 14, 15, 19, 20 We used the mutant bacterial strain, PhoPC, for this chronic infection because wild-type (WT) Salmonella induced death of infected mice within a week.18 We therefore focused on mutated bacterial strains for these chronic infection and cancer studies. Previous studies have indicated that PhoPC strain expressing AvrA activates the β-catenin pathway in acute and chronic infection.14, 21 PhoP-PhoQ is a regulatory system that controls the expression of Salmonella virulence and survival. PhoPC is a PhoP-PhoQ constitutive mutation.22 We demonstrate for the first time that bacterial effector AvrA has an essential role in the effects of chronic Salmonella infection on colonic tumorigenesis. Our findings provide important new insights into mechanisms by which the bacterial protein AvrA contributes to colon cancer development.
Results
Persistence of Salmonella colonization in the colon
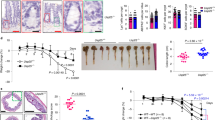
Mice were infected with the indicated Salmonella strains by one oral gavage and then treated with an AOM-DSS protocol to induce colon cancer. The treatment protocol is summarized in Figure 1a. Colonic Salmonella persistence was confirmed by Salmonella culture (pink colonies) and 16S RNA PCR of fecal bacteria DNA (Figures 1b and c). AvrA persistence was also confirmed by western blotting of colon tissue from mice infected with AvrA-expressing bacteria 45 weeks postinfection (Figure 1d). Furthermore, we confirmed persistent colonization of Salmonella in the colon by immunostaining (Figure 1e). Note that Salmonella was identified at the base and surface of the crypts in the infected colon (indicated by arrows) (Figure 1e). We found no significant changes in body weight associated with infection (Supplementary Figure S1). Salmonella levels in the intestine, gallbladder, spleen and liver also correlated with expression of AvrA in infected mice (Supplementary Figure S2).

Persistent Salmonella infection in mice. Mice were infected with the indicated Salmonella strains by oral gavage and treated with AOM-DSS to induce colon cancer. (a) Schema of experimental design. Day-1: Mice were infected by Salmonella gavage 24 h after streptomycin pretreatment (7.5 mg/mouse), ↓azoxymethane (AOM), 10 mg/kg body weight, intraperitoneal injection; ▪ 1% dextran sodium sulfate (DSS) in drinking water, ↓ Salmonella gavage; ▴ mice were killed. Experimental groups: Control, normal WT mice (no Salmonella, no AOM/DSS); AOM/DSS, only (no Salmonella), PhoPC, AOM/DSS-treated mice infected with Salmonella with WT AvrA; PhoPCAvrA−, AOM/DSS-treated mice infected with AvrA− mutant strain derived from PhoPC; PhoPCAvrA−/AvrA+, AOM/DSS-treated mice infected with PhoPCAvrA− transfected with plasmid encoding AvrA. (b) Chronic Salmonella infection was assessed by fecal cultures at week 45. On a BBL CHROMagar plate, intestinal Salmonella species appear mauve (rose to purple) in color, due to metabolic differences in the presence of selected chromogens. Other bacteria are either inhibited or produce blue–green or colorless colonies. (c) Salmonella detected in fecal microbial DNA by 16S rRNA PCR 45 weeks postinfection. Mice were infected with PhoPC expressing AvrA protein or PhoPC AvrA− (AvrA deficient) or AvrA-deficient bacteria transfected with AvrA-expressing plasmid. (d) AvrA western blotting. Protein lysates from colonic tissue were probed for AvrA levels. (e) Immunofluorescence confocal microscopy. Salmonella invading the colonic epithelium 45 weeks postinfection appear green.
Persistent infection with Salmonella expressing AvrA increases tumor incidence and multiplicity and shifts tumor distribution to proximal colon
We collected tissue samples 45 weeks postinfection. Representative colons with tumors are shown in Figure 2a. There was a striking difference in tumor incidence in mice with AvrA+ and AvrA− bacterial colonization (Table 1). The tumor incidence was 51.4% in the AOM/DSS control group (no infection) and 60.6% in the parental PhoPC-infected mice, 56.3% in the AvrA- infected mice and 100% in the PhoPC AvrA−/AvrA+.

AvrA-expressing bacteria significantly increase colonic tumor incidence. (a) Colonic tumors in situ. Representative colons from the indicated groups 45 weeks after bacterial infection. Tumors are indicated by red arrows. (b) Tumor multiplicity. Average number of tumors in each experimental group. *P<0.05, **P<0.01. (c) Tumor location. The distance of each tumor from the anus was measured. *P<0.05. (d) Swiss rolls of representative colons from the indicated groups. Rectangles in the middle panel are shown at higher magnification in the lower panel. (e) Salmonella invasion. Localization of Salmonella in tumor tissue was assessed by immunofluorescence staining.
The average number of tumors in colon (tumor multiplicity) was also significantly higher in the PhoPC AvrA−/AvrA+ compared with the AvrA− group or the AOM/DSS group without infection (Figure 2b). We also analyzed the distribution of tumors. AOM/DSS-treated mice that were not gavaged with Salmonella were located in the distal colon (Figure 2c, AOM/DSS). In contrast, in mice infected with PhoPC or PhoPC AvrA−/AvrA+ Salmonella, the tumors were located more proximally (Figure 2c). We investigated the pathological changes associated with chronic Salmonella infection in AOM/DSS mice using Swiss rolls (Figure 2d). We also found greater number of cancers in AvrA+ compared with AvrA− mice treated with AOM/DSS (Table 1, 40% versus 12.5%, respectively). However, we did not see significant differences in tumor size among the various experimental groups (Supplementary Figure S3).
We detected Salmonella by immunofluorescence invading the colonic mucosa 45 weeks postinfection. Salmonella organisms were present not only on the colonic luminal surface and within the crypt as shown in Figure 1e but also in tumors from infected mice (Figure 2e). Furthermore, the load of invading Salmonella appeared to be higher in tumors, especially in mice colonized with AvrA(+)-expressing Salmonella. Thus, these data demonstrate that a single gavage of Salmonella was sufficient to induce persistent Salmonella colonization, and upregulation of AvrA enhanced tumorigenesis and tumor burden in the AOM/DSS model and altered tumor distribution in the colon.
Activation of the β-catenin signaling in the Salmonella chronically infected colon.
It is known that β-catenin signaling is activated in colonic tumorigenesis.23 In agreement with previous studies, we observed that nuclear β-catenin was upregulated in AOM/DSS-induced tumors (Figure 3a). Nuclear localization suggested that β-catenin is also activated (Figure 3a).

AvrA regulates β-catenin activation as assessed by phospho (Ser552) β-catenin in AOM/DSS-induced tumors. (a) β-Catenin localization in control mucosa and colonic tumors. (b) P-β-catenin Ser552 localization in control mucosa and colonic tumors. (c) Quantitation of nuclear P-β-catenin Ser552 in control mucosa and colonic tumors. Data are means±s.d. of n=3 control mice or AOM/DSS tumors in each group. *P<0.05, **P<0.01 (d) Expression levels of p-β-catenin, total β-catenin and Akt and P-Akt as assessed by western blotting in control and AOM/DSS-induced tumors. Shown are control mucosa and tumors representative of three mice in each group. (e) Immunostaining of p-Myc, MMP-7, Cyclin D1 and Bmi1 in control colons and AOM/DSS-induced tumors (left panel). Quantitative imaging of nuclear Bmi1 in control mucosa and mucosa adjacent to tumors (right panel). Data are means±s.d. of n=3 control mice or AOM/DSS mice in each group. *P<0.05, **P<0.01.
Residue specific phosphorylations control activation or destabilization of β-catenin and are regulated by several upstream kinases. Inflammation increases phosphorylation of β-catenin on Ser552 in stem progenitor cells inducing proliferation.24 In tumors from mice infected with Salmonella, AvrA expression increased phosphorylated β-catenin Ser552 whereas tumors from mice infected with mutant Salmonella deficient in AvrA had significantly lower levels of phosphorylated β-catenin Ser552 (Figures 3b and c). By western blotting, we confirmed that protein levels of phosphorylated β-catenin Ser552 were higher in tumors from mice infected with AvrA-expressing bacteria, compared with control mice treated only with AOM/DSS alone or also colonized with Salmonella deleted of AvrA (Figure 3d). In contrast, we found no significant change in phosphorylated Ser33/Ser37 or Thr41 associated with AvrA expression in tumors from mice infected with Salmonella (Figure 3d), whereas it was increased slightly in control mice treated with AOM/DSS alone. β-Catenin phosphorylations on residues Ser33/Ser37 and Thr41 are associated with destabilization and degradation of β-catenin. This might explain why we did not find significant AvrA-related differences in total β-catenin in tumors from chronically infected mice.
Akt is an upstream regulator of the β-catenin pathway and phosphorylates β-catenin on Ser552.25 This phosphorylation is implicated in nuclear β-catenin localization.25 Because there was increased β-catenin Ser552 phosphorylation with bacterial AvrA, we investigated Akt activation in tumors from infected mice. We observed that phospho-active Akt was also increased in tumors, especially in the PhoPCAvrA−/AvrA+ group, whereas total Akt levels were not changed in tumors from the PhoPC- and PhoPCAvrA−/AvrA+-infected mice (Figure 3d). The phospho active-β-catenin phosphorylated on Ser552 was also increased in colonocytes 15 and 35 weeks after Salmonella gavage in the PhoPC and PhoPCAvrA−/AvrA+ groups (data not shown). Thus colonic phospho active-β-catenin (phospho-Ser552 β-catenin) is persistently elevated in mice colonized by Salmonella expressing AvrA.
The Bmi1 (B-cell-specific Moloney murine leukemia virus integration site 1) gene was first isolated as an oncogene that cooperates with c-myc in oncogenesis.26 Bmi1 is also implicated as a stem cell marker.27 We found that colonic Bmi1 was significantly increased in AvrA colonized mice (Figure 3e). Although there was no detectable Bmi1 in the tumors by staining, enhanced Bmi1 staining was found in crypts adjacent to tumor of the PhoPC- and PhoPCAvrA−/AvrA+-infected mice (Figure 3e).
We next assessed Akt and β-catenin target genes. Although we did not see significant changes in total c-Myc expression, phospho-c-Myc (P-S62) was upregulated in tumors from mice infected with AvrA-expressing bacteria. Colonic levels of MMP-7 (matrix metalloproteinase 7) and cyclin D1 were also increased in mice colonized with Salmonella expressing AvrA (Figure 3e).
Salmonella effector protein AvrA modulates β-catenin ubiquitination in vitro and in vivo
Our previous study showed that AvrA protein acts as a deubiquitinase to stabilize β-catenin.14 To determine whether AvrA is responsible for the activation of β-catenin signaling through ubiquitination, we investigated the ubiquitinated β-catenin levels in the AOM/DSS mice with or without Salmonella (Figure 4a). The ubiquitinated proteins results in the appearance of multiple higher molecular weight bands above the regular protein band in western blotting. We found that mice colonized with Salmonella strain PhoPC and PhoPC AvrA−/AvrA+ lacked detectable ubiquitinated β-catenin bands (Figure 4a). In contrast, in the PhoPC AvrA− group, ubiquitinated β-catenin levels were similar to the control AOM/DSS mice without Salmonella (Figure 4a).

Bacterial effector protein AvrA modulates β-catenin ubiquitination in colonic epithelial cells. (a) AvrA expression decreased the ub-β-catenin in the colonic epithelial cells. At 6 weeks after Salmonella infection, colonic epithelial cells were collected and immunoprecipitated with anti-β-catenin antibody. Membranes were probed with anti-β-catenin and anti-ubiquitin antibody and visualized using enhanced chemiluminescence. Protein input was normalized by the colonic epithelial marker villin. Ubiquitinated (ub) β-catenin is shown as smear. Colon infected by the PhoPC lacked the ub-β-catenin bands. (b) AvrA expression decreased the ub-β-catenin through ubiquitination in the human colonic epithelial HCT116 cells. Cells were transiently cotransfected with AvrA and the indicated ubiquitin plasmids; 24 h later, the cells were treated with MG262 for 3 h. The ubiquitin plasmids included pRK5-HA-Ub-KO (all lysines mutated), -Ub WT, -Ub-K48 (K48 only, other lysines mutated) and -Ub-K63 (K63 only, other lysines mutated). Equal amounts of total cell lysates were analyzed for protein levels by immunoblot with anti-β-catenin antibody. (c) AvrA expression mostly removes k48-link ubiquitins in the human colonic epithelial HCT116 cells. Cells were transiently cotransfected with AvrA and the indicated ubiquitin plasmids. Equal amounts of total cell lysates were analyzed for protein levels by immunoblot with anti-ub-48 antibody.
The protein modifications can be either a single ubiquitin protein or chains of ubiquitin. Ubiquitin has seven lysines (K6, K11, K27, K29, K33, K48 and K63), all of which can be conjugated to another ubiquitin to form a polyubiquitin chain.28 K63 and K48 linkage-specific ubiquitinations are the most predominant forms of ubiquitination.29 To explore the molecular mechanism of AvrA on ubiquitination, we then transiently cotransfected AvrA and different ubiquitin plasmids in the human colonic epithelial HCT116 cells. The ubiquitin plasmids included pRK5-HA-Ub-KO (all lysines mutated to arginines), -Ub WT, -Ub-K48 (K48 only, other lysines mutated to arginines) and -Ub-K63 (K63 only, other lysines mutated to arginines). As expected, AvrA expression decreased the ubiquitinated β-catenin (ub-β-catenin) in the ub-WT group (Figure 4b, lanes 3 and 4). In the presence of AvrA expression and ub-K48, the ubiquitinated β-catenin (ub-β-catenin) was decreased (Figure 4b, lane 7, indicated by an arrow), whereas ub-β-catenin levels were more in the absence of AvrA (Figure 4b, lane 8). In contrast, there is no difference in the ub-K63 group with or without AvrA (Figure 4b, lanes 5 and 6). As a negative control, the ub-KO group with or without AvrA (Figure 4b, lanes 1 and 2) did not show any difference of ubiquitinated β-catenin due to the mutation of the all lysines to arginines. Immunobloting with the anti-Ub-K48 further showed that AvrA protein mostly decreased UbK48 linkage-specific ubiquitination (Figure 4c, lane 5). However, we did not find significant change of Ub K6, K11, K27, K29 or K33 (data not shown). These in vivo and in vitro data indicate that bacterial effector AvrA prevents β-catenin ubiquitination in epithelial cells.
AvrA activates β-catenin signaling pathways in the Salmonella-infected cells in vitro
To further investigate the role of bacterial AvrA in modulating the host signaling pathways in vitro, we transfected human HEK293 cells with AvrA and TCF lucifierase reporter plasmid. We found that AvrA could increase the β-catenin activity as assessed by increased luciferase activity in cells transfected with a PGL3-OT plasmid with WT TCF-binding site (Figure 5a). In contrast, AvrA did not increase activation of mutated PGL3-OF that is unresponsive to β-catenin–TCF transactivation.

Bacterial AvrA activates β-catenin signaling. (a) AvrA induces β-catenin signaling in HEK293 cells. HEK293 cells were transiently co-transfected with a pcmv-myc-AvrA plasmid and a pGL3-OT plasmid (TCF-responsive reporter with WT TCF binding site) or pGL3-OF (mutant TCF binding site), using lipofectin reagent according to the manufacturer’s instructions. LiCl treatment (20 mmol/l) was used as the positive control. Each experiment was assayed in triplicate. N=3 independent platings *P<0.05. (b) Colonic BrdU was detected in normal mucosa and tumors in control mice (AOM/DSS alone) or mice also gavaged with the indicated bacteria 45 weeks postinfection. (c) Quantitation of BrdU staining in normal tissue and tumors from the indicated groups. *P<0.05, **P<0.01. (d) Model of AvrA regulation of β-catenin signaling in colonic tumorigenesis. In mice chronically infected with Salmonella, AvrA activates β-catenin, which upregulates its target genes to promote colonic tumorigenesis.
β-Catenin signals drive proliferation that is mediated by several target genes, including c-myc and cyclin D1.30, 31, 32 AvrA upregulated colonic Myc and cyclin D1 expression in Salmonella-colonized mice (Figure 3e). In the ‘normal’ tissue close to tumors, we also found proliferation marker BrdU (bromodeoxyuridine) incorporation was increased in control colon and tumors from mice infected with AvrA-expressing bacteria (Figure 5b). Taken together, our data indicate that AvrA activates the β-catenin signals in vitro and in vivo in Salmonella-infected murine colonocytes and tumor cells.
Discussion
In the current study, we investigated the effects of chronic Salmonella infection on tumor development in the AOM/DSS model. We used a genetic approach to dissect the effects of AvrA on tumorigenesis, comparing Salmonella sufficient in AvrA to microbes deficient in AvrA or AvrA-deficient microbes stably transfected to express AvrA. We showed for the first time that AvrA enhanced proliferation and promoted colonic tumorigenesis and tumor progression and concomitantly activated proto-oncogene β-catenin and its down-stream targets c-Myc and cyclin D1. In part, upregulated β-catenin signals may be mediated by increased phosphorylation of Ser552 and decreased β-catenin ubiquitination. AvrA expression also increased the abundance of Salmonella organisms in tumors, confirming a direct effect of AvrA on Salmonella invasiveness (data in Figure 2e, summarized in Figure 5d). This study provides novel and potentially important insights into bacterial mechanisms promoting colonic tumorigenesis and tumor promotion. Specifically, our studies identify for the first time a novel tumor-promoting role for AvrA in promoting tumorigenesis.
WT mice rarely develop spontaneous colon cancers but require a carcinogen such as AOM to induce tumorigenesis. As C57B/6 mice are relatively resistant to AOM alone, we used the AOM/DSS model to promote robust colonic tumorigenesis. Because the AOM/DSS model is driven by β-catenin signals and AvrA was shown to activate the β-catenin pathway in colonocytes, we speculated that AvrA might enhance tumorigenesis. Infections with Salmonella expressing AvrA, however, were insufficient to induce colonic tumors in the absence of AOM or AOM/DSS. When combined with AOM and DSS, however, AvrA enhanced tumor initiation and progression, suggesting AvrA possesses co-carcinogen activity.
Salmonella AvrA altered tumor distribution
The PhoPC AvrA−/AvrA+ Salmonella strain is known to express high levels of AvrA.25 Supporting the oncogenic role of AvrA, the PhoPC AvrA−/AvrA+ group had the highest tumor incidence and cancer burden. In contrast, chronic infection with AvrA-negative Salmonella was not sufficient to increase tumor incidence. These data indicate that bacterial protein AvrA promotes oncogenesis in this model.
In addition to AvrA-enhanced tumor multiplicity and tumor progression, another novel finding in the current study was that this pathogenicity factor altered tumor distribution. In agreement with previous studies,33, 34 uninfected mice treated with AOM/DSS developed tumors in the distal colon (Figure 2c, AOM/DSS). In contrast, in mice infected with AvrA-expressing bacteria, tumors were found more in the proximal colon. We conclude that AvrA alters the colonic milieu so as to enhance tumorigenesis in the right colon. Compared with the left colon, the cecum has a greater bacterial load and increased bacterial fermentation that we speculate contributes to this rightward shift in tumors (Figure 1e). Increasing incidence in right-sided tumors has also been reported in the Western world. While increased endoscopic screening that probably clears distal colonic lesions more effectively than proximal colonic lesions, based on our studies, this shift might also reflect changes in the microbiome.
AvrA-expressing Salmonella strains enhance β-catenin signaling and upregulate BMI1 stem cell marker.
A growing body of literature suggests that chronic inflammation leads to tissue damage, stem cell expansion and a predisposition to transforming mutations.35 In this regard, colonic stem cells are known to have a critical role in the pathogenesis of colon cancer.36 We found that Bmi1, a marker of stem cells, was increased in an AvrA-dependent manner. AvrA also increased levels of phosphorylated Ser552 β-catenin and colonic crypt cell proliferation. β-Catenin is involved in inflammation-induced stem progenitor cell expansion.24, 25 In previous publications, we demonstrated that AvrA inhibited β-catenin degradation and increased β-catenin transcriptional activity through the Wnt pathway.15, 16, 21 As β-catenin activation is a critical step in stem cell proliferation and colonic tumorigenesis, our current findings regarding AvrA induction of β-catenin activation place our previous observations in a pathophysiological context relevant to cancer and highlights the potential synergistic role of aberrant Wnt signaling (driven by APC or β-catenin mutations) and AvrA to increase β-catenin activation. Thus AvrA upregulation of β-catenin Ser552 phosphorylation provides new insights into the mechanism by which AvrA can expand stem cells by activating the β-catenin pathway in vivo.
AvrA modulates β-catenin ubiquitination
Ubiquitination is a key posttranslational protein modification that controls cellular protein levels. In our previous study, we found that AvrA can regulate colonic epithelial cell inflammation by deubiquitination.15 Here we found that AvrA deubiquitinates β-catenin and thus blocks degradation of this key oncogenic Wnt effector protein during colonic tumorigenesis. We also investigated which ubiquitin lysine residues are controlled by AvrA. We demonstrated that AvrA stabilized β-catenin at the ubiquitination level by blocking lysine 48 ubiquitination. Without degradation, β-catenin can accumulate in the cytoplasm. And in the presence of activated AKT, phospho-active β-catenin translocates into the nucleus. Nuclear β-catenin heterodimerizes with TCF to stimulates transcription of target genes, including c-Myc and cyclin D1 (Figure 5d).
In summary, we have uncovered novel effector functions of Salmonella-associated AvrA that activates β-catenin pathways to promote tumorigenesis. Salmonella expressing AvrA significantly increased tumor multiplicity and tumor progression and shifted tumor distribution more proximally. This study increases our understanding of the role of bacteria in colon cancer. Our observations of Salmonella effects on tumorigenesis also raise a note of caution regarding the use of mutant Salmonella organisms as vectors for anti-cancer therapy. Further studies are needed to confirm whether AvrA is expressed in these strains and the safety of such approaches. Finally, our results bring us one step closer to unraveling infection-related dysregulation of β-catenin signaling in cancer. These studies, moreover, could suggest biomarkers (such as AvrA level in gut) to assess colon cancer risk in susceptible individuals.
Materials and methods
Ethics statement
This study was carried out in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health. The protocol was approved by the University of Rochester University Committee on Animal Resources (UCAR) committee (UCAR 2007-065) and Rush University Institutional Animal Care and Use Committee (IACUC Number 12-018). All efforts were made to minimize suffering.
Bacterial strains and growth condition
Salmonella typhimurium strains used in this study included Salmonella mutant strains PhoPC,22 PhoPCAvrA−, and PhoPCAvrA−/AvrA+.14 Non-agitated microaerophilic bacterial cultures were prepared by inoculating 10 ml of Luria–Bertani broth with 0.01 ml of a stationary-phase culture followed by overnight incubation (∼18 h) at 37 °C, as previously described.14
Salmonella-infected colon cancer mouse model
Animal experiments were performed using specific pathogen-free female C57BL/6 mice (Taconic, Albany, NY, USA) that were 6–7-weeks old as previously described.20 Water and food were withdrawn 4 h before oral gavage with 7.5 mg/mouse of streptomycin (100 μl of sterile solution). Afterwards, animals were supplied with water and food ad libitum. Twenty hours after streptomycin treatment, water and food were withdrawn again for 4 h before the mice were infected with 1 × 106 colony-forming units of Salmonella (100-μl suspension in HBSS (Hank’s Balanced Salt Solution)) or treated with sterile HBSS (control) by oral gavage, as previously described.37 After Salmonella gavage, the mice were treated with 10 mg/kg of AOM (Sigma-Aldrich, Milwaukee, WI, USA) by intraperitoneal injection as described by Greten and Karin.38 After a 7-day recovery period, the animals received 1% DSS ad libitum (see schematic in Figure 1a). The initial sample size was n=40 mice in the control group with no treatment and n=50 in each experimental group. Sixteen weeks later, mice received second AOM injection and another three cycles of 1% DSS in the drinking water. Tissue samples were collected at 1, 15, 25, 35 and 45 weeks after Salmonella gavage. Tumor counts and measurements were performed in a blinded fashion under a stereo-dissecting microscope (Nikon SMZ1000, Melville, NY, USA). Microscopic analysis was performed for severity of inflammation and dysplasia39 on hematoxylin and eosin-stained ‘Swiss rolled’ colons by a gastrointestinal pathologist blinded to treatment conditions.
Detection of Salmonella in intestine
Mouse feces (about 100 mg) were collected, vortexed in phosphate-buffered saline (PBS), and centrifuged for 10 min at 800 r.p.m. The sample was transferred to a clean microfuge tube and centrifuged again at 6000 r.p.m. for 5 min. The supernatant was discarded, and 200 μl PBS was added to the pellet, which was then vortexed. Colonic Salmonella was detected by culturing fecal content at 37 °C overnight on a BBL CHROMagar plate (BD Biosciences, San Jose, CA, USA). Salmonella species appeared mauve (rose to purple).
Microbial analysis
After euthanasia, 1-ml samples from the cecal contents of colonized mice were serially diluted in PBS. Total bacterial concentrations were determined using counting chambers. Colonic Salmonella distribution was determined by immunostaining colon sections using anti-Salmonella lipopolysaccharide antibody (Santa Cruz Biotechnology, Santa Cruz, CA, USA).16, 18, 25 The presence of Salmonella existence was also confirmed by real-time PCR measurements of Salmonella 16S rRNA. DNA was extracted from colonic tissues after washing to remove residual feces. Primers specific to 18S rRNA from intestinal cells in fecal samples were used as an endogenous control to normalize loading between samples. For the quantification of Salmonella, a pair of primers specific for Salmonella species of 16S rRNA were used to amplify this bacterial gene: 16S rRNA Fw 5′-TATAGCCCCATCGTGTAGTCAGAAC-3′, Rv 5′-TGCGGCTGGATCACCTCCTT-3′; and 18S rRNA Fw 5′-AGGGGAGAGCGGGTAAGAGA-3′, Rv 5′-GGACAGGACTAGGCGGAACA-3′. The relative amount of 16S rRNA in each sample was estimated using the ΔΔCT method.
Salmonella burden in the intestine, gallbladder, spleen and liver
Intestine, gallbladder, spleen and liver were harvested from each mouse, placed into 14-ml tubes with 5 ml sterile PBS, minced with scissors and homogenized using a Polytron PT2100 (Luzern, Switzerland). Tissue homogenates were diluted 1000x to 10 000x with LB, plated (100 μl) on MacConkey agar plates, and incubated overnight at 37 °C. Colony-forming units were quantified.
Histological testing
Tissues were fixed in 10% neutral buffered formaldehyde for 4–12 h, transferred into 70% ethanol and processed by standard techniques. Sections (5 μm) were stained with hematoxylin and eosin. For immunostaining, antigens were retrieved by 10-min boiling in 10 mM citrate (pH 6.0). Blinded histological inflammatory scores were performed by a validated scoring system.40 Histological assessment of intestinal tumors in the rodent was performed using the recommended standards.41 Immunostaining, including BrdU staining, was performed as previously described.15, 42 The slides were stained with anti-BrdU, anti-Bmi1, anti-phospho-c-Myc, anti-cyclin D1, anti-MMP-7 (Abcam, Cambridge, MA, USA), anti-phospho-β-catenin (Ser552), anti-β-catenin (BD, Transduction Laboratories, Lexington, KY, USA).
Immunoblotting
Mouse colonic mucosa was collected by scraping proximal and distal regions as previously described.20 Tumors were collected separately. Mouse epithelial cells were lysed in lysis buffer (1% Triton X-100, 150 mM NaCl, 10 mM Tris pH 7.4, 1 mM EDTA, 1 mM EGTA pH 8.0, 0. 2 mM sodium orthovanadate, protease inhibitor cocktail). Immunoblotting was performed with primary antibodies: anti-AvrA (custom-made in Sub lab13), anti-β-catenin (BD, Transduction Laboratories), anti-Bmi 1 (Abcam), and anti-villin (Santa Cruz Biotechnology Inc.), anti-GSK-3β (BD), anti-phospho-GSK-3β (Ser9), anti-phospho-β-catenin (Ser552), anti-phospho-β-catenin (Ser33, Ser37, Thr41) and anti-phospho-Akt (T308) (Cell Signaling, Beverly, MA, USA), or anti-β-actin (Sigma-Aldrich) antibodies, and visualized by enhanced chemiluminescence.43, 44
Immunofluorescence
Colonic tissue and colonic tumors were freshly isolated fixed in 10% neutral buffered formalin and embedded in paraffin. Immunohistochemistry was performed on formalin-fixed paraffin-embedded sections (5 μm) of mouse colons. After tissue preparation, as described previously,15 sections were stained with anti-Salmonella lipopolysaccharide antibody (Santa Cruz Biotechnology).16, 18, 25 Specimens were examined with a Leica SP5 Laser Scanning confocal microscope (Buffalo Grove, IL, USA).
Transfection and luciferase assay for β-catenin transcriptional activity
The transcriptional activity of β-catenin was measured using the TCF Reporter Assay.14 Briefly, HEK293 cell line was co-transfected with a WT TCF/LEF-responsive firefly luciferase reporter (pGL3-OT) or a mutant firefly luciferase reporter (pGL3-OF) as a negative control, as well as specific pCMV-AvrA or pCMV. LiCl treatment (20 mM for 24 h after transfection) was used as the positive control. PRL-TK vector was used to control for transfection efficiency. Luciferase activity was monitored using the dual luciferase assay system (Promega, Madison, WI, USA). Experiments were done in triplicate with three independent platings.
AvrA and ubiquitin assay in vitro
HCT116 cells were transiently cotransfected with AvrA and the indicated ubiquitin plasmids, including pRK5-HA-Ub WT, -Ub K48, -UbK63 (K48/63/33 only, other lysines mutated to arginines) and -Ub KO (all lysines mutated to arginines). Plasmids were purchased from Addgene (Cambridge, MA, USA). After 24 h, cells were treated with MG262 for 3 h. Total cell lysates were analyzed by immunoblotting.14, 15
Coimmunoprecipitation assay
Animal experiments were performed using 6–7-weeks-old specific pathogen-free female C57BL/6 mice (Taconic). Mice were infected with the indicated Salmonella strains by oral gavage. At 6 weeks after Salmonella infection, colonic mucosal scrapings were rinsed twice in ice-cold HBSS and lysed in cold immunoprecipitation buffer (1% Triton X-100, 150 mM NaCl, 10 mM Tris pH 7.4, 1 mM EDTA, 1 mM EGTA pH 8.0, 0.2 mM sodium orthovanadate and protease inhibitor cocktail (Boehringer, Mannheim, Germany)). Samples were immunoprecipitated with anti-β-catenin antibody. Membranes were probed with anti-β-catenin and anti-ubiquitin antibody and visualized using enhanced chemiluminescence. Protein input was normalized by villin, a colonic epithelial cell marker.
Statistical analysis
Data were expressed as mean±s.d. All statistical tests were two-sided. P-values <0.05 were considered to be statistically significant. For cecum data, four areas of mouse body were checked to obtain the average infection score. Then, multiple comparisons of mean infection scores were performed using analysis of variance. Following analysis of overall group differences on the mean scores at P<0.05, αlevels were adjusted using the Tukey’s correction for multiple comparisons. As the study involved a longitudinal design, the generalized estimating equations were utilized to provide statistical inference.45 The analysis was conducted using the Genmod procedure in SAS. The overall treatment effects were modeled using the time by treatment interaction term through intake ( week 0 ) to week 45.18 Statistical analyses were performed using the SAS version 9.2 (SAS Institute, Inc., Cary, NC, USA).
References
Coussens LM, Werb Z . Inflammation and cancer. Nature 2002; 420: 860–867.
Mantovani A . Cancer: inflaming metastasis. Nature 2009; 457: 36–37.
Wu S, Rhee KJ, Albesiano E, Rabizadeh S, Wu X, Yen HR et al. A human colonic commensal promotes colon tumorigenesis via activation of T helper type 17 T cell responses. Nat Med 2009; 15: 1016–1022.
Arthur JC, Perez-Chanona E, Muhlbauer M, Tomkovich S, Uronis JM, Fan TJ et al. Intestinal inflammation targets cancer-inducing activity of the microbiota. Science 2012; 338: 120–123.
Gradel KO, Nielsen HL, Schonheyder HC, Ejlertsen T, Kristensen B, Nielsen H . Increased short- and long-term risk of inflammatory bowel disease after salmonella or campylobacter gastroenteritis. Gastroenterology 2009; 137: 495–501.
Kumar S . Infection as a risk factor for gallbladder cancer. J Surg Oncol 2006; 93: 633–639.
Zhao M, Geller J, Ma H, Yang M, Penman S, Hoffman RM . Monotherapy with a tumor-targeting mutant of Salmonella typhimurium cures orthotopic metastatic mouse models of human prostate cancer. Proc Natl Acad Sci USA 2007; 104: 10170–10174.
Arrach N, Zhao M, Porwollik S, Hoffman RM, McClelland M . Salmonella promoters preferentially activated inside tumors. Cancer Res 2008; 68: 4827–4832.
Zhao M, Yang M, Li XM, Jiang P, Baranov E, Li S et al. Tumor-targeting bacterial therapy with amino acid auxotrophs of GFP-expressing Salmonella typhimurium. Proc Natl Acad Sci USA 2005; 102: 755–760.
Collier-Hyams LS, Zeng H, Sun J, Tomlinson AD, Bao ZQ, Chen H et al. Cutting edge: Salmonella AvrA effector inhibits the key proinflammatory, anti-apoptotic NF-kappa B pathway. J Immunol 2002; 169: 2846–2850.
Du F, Galan JE . Selective inhibition of type III secretion activated signaling by the Salmonella effector AvrA. PLoS Pathog 2009; 5: e1000595.
Hardt WD, Galan JE . A secreted Salmonella protein with homology to an avirulence determinant of plant pathogenic bacteria. Proc Natl Acad Sci USA 1997; 94: 9887–9892.
Liao AP, Petrof EO, Kuppireddi S, Zhao Y, Xia Y, Claud EC et al. Salmonella type III effector AvrA stabilizes cell tight junctions to inhibit inflammation in intestinal epithelial cells. PLoS ONE 2008; 3: e2369.
Sun J, Hobert ME, Rao AS, Neish AS, Madara JL . Bacterial activation of beta-catenin signaling in human epithelia. Am J Physiol Gastrointest Liver Physiol 2004; 287: G220–G227.
Ye Z, Petrof EO, Boone D, Claud EC, Sun J . Salmonella effector AvrA regulation of colonic epithelial cell inflammation by deubiquitination. Am J Pathol 2007; 171: 882–892.
Liu X, Lu R, Wu S, Sun J . Salmonella regulation of intestinal stem cells through the Wnt/beta-catenin pathway. FEBS Lett 2010; 584: 911–916.
Grassl GA, Valdez Y, Bergstrom KS, Vallance BA, Finlay BB . Chronic enteric salmonella infection in mice leads to severe and persistent intestinal fibrosis. Gastroenterology 2008; 134: 768–780.
Lu R, Wu S, Liu X, Xia Y, Zhang YG, Sun J . Chronic effects of a Salmonella type III secretion effector protein AvrA in vivo. PLoS ONE 2010; 5: e10505.
Sun J, Hobert ME, Duan Y, Rao AS, He TC, Chang EB et al. Crosstalk between NF-kappaB and beta-catenin pathways in bacterial-colonized intestinal epithelial cells. Am J Physiol Gastrointest Liver Physiol 2005; 289: G129–G137.
Duan Y, Liao AP, Kuppireddi S, Ye Z, Ciancio MJ, Sun J . beta-Catenin activity negatively regulates bacteria-induced inflammation. Lab Invest 2007; 87: 613–624.
Lu R, Liu X, Wu S, Xia Y, Zhang YG, Petrof EO et al. Consistent activation of the beta-catenin pathway by Salmonella type-three-secretion effector protein AvrA in chronically infected intestine. Am J Physiol Gastrointest Liver Physiol 2012; 303: G1113–G1125.
Miller SI, Mekalanos JJ . Constitutive expression of the phoP regulon attenuates Salmonella virulence and survival within macrophages. J Bacteriol 1990; 172: 2485–2490.
Sellin JH, Umar S, Xiao J, Morris AP . Increased beta-catenin expression and nuclear translocation accompany cellular hyperproliferation in vivo. Cancer Res 2001; 61: 2899–2906.
He XC, Yin T, Grindley JC, Tian Q, Sato T, Tao WA et al. PTEN-deficient intestinal stem cells initiate intestinal polyposis. Nat Genet 2007; 39: 189–198.
Wu S, Ye Z, Liu X, Zhao Y, Xia Y, Steiner A et al. Salmonella typhimurium infection increases p53 acetylation in intestinal epithelial cells. Am J Physiol Gastrointest Liver Physiol 2010; 298: G784–G794.
Haupt Y, Alexander WS, Barri G, Klinken SP, Adams JM . Novel zinc finger gene implicated as myc collaborator by retrovirally accelerated lymphomagenesis in E mu-myc transgenic mice. Cell 1991; 65: 753–763.
Sangiorgi E, Capecchi MR . Bmi1 is expressed in vivo in intestinal stem cells. Nat Genet 2008; 40: 915–920.
Liu S, Chen ZJ . Expanding role of ubiquitination in NF-kappaB signaling. Cell Res 2011; 21: 6–21.
Dammer EB, Na CH, Xu P, Seyfried NT, Duong DM, Cheng D et al. Polyubiquitin linkage profiles in three models of proteolytic stress suggest the etiology of Alzheimer disease. J Biol Chem 2011; 286: 10457–10465.
Shtutman M, Zhurinsky J, Simcha I, Albanese C, D'Amico M, Pestell R et al. The cyclin D1 gene is a target of the beta-catenin/LEF-1 pathway. Proc Natl Acad Sci USA 1999; 96: 5522–5527.
Moumen M, Chiche A, Decraene C, Petit V, Gandarillas A, Deugnier MA et al. Myc is required for beta-catenin-mediated mammary stem cell amplification and tumorigenesis. Mol Cancer 2013; 12: 132.
Hu D, Fang W, Han A, Gallagher L, Davis RJ, Xiong B et al. c-Jun N-terminal kinase 1 interacts with and negatively regulates Wnt/beta-catenin signaling through GSK3beta pathway. Carcinogenesis 2008; 29: 2317–2324.
Dougherty U, Mustafi R, Wang Y, Musch MW, Wang CZ, Konda VJ et al. American ginseng suppresses Western diet-promoted tumorigenesis in model of inflammation-associated colon cancer: role of EGFR. BMC Complement Altern Med 2011; 11: 111.
Tao Y, Hart J, Lichtenstein L, Joseph LJ, Ciancio MJ, Hu S et al. Inducible heat shock protein 70 prevents multifocal flat dysplastic lesions and invasive tumors in an inflammatory model of colon cancer. Carcinogenesis 2009; 30: 175–182.
Houghton J, Stoicov C, Nomura S, Rogers AB, Carlson J, Li H et al. Gastric cancer originating from bone marrow-derived cells. Science 2004; 306: 1568–1571.
Ricci-Vitiani L, Pagliuca A, Palio E, Zeuner A, De Maria R . Colon cancer stem cells. Gut 2008; 57: 538–548.
McCormick BA, Colgan SP, Delp-Archer C, Miller SI, Madara JL . Salmonella typhimurium attachment to human intestinal epithelial monolayers: transcellular signalling to subepithelial neutrophils. J Cell Biol 1993; 123: 895–907.
Greten FR, Karin M . The IKK/NF-kappaB activation pathway-a target for prevention and treatment of cancer. Cancer Lett 2004; 206: 193–199.
Dieleman LA, Palmen MJ, Akol H, Bloemena E, Pena AS, Meuwissen SG et al. Chronic experimental colitis induced by dextran sulphate sodium (DSS) is characterized by Th1 and Th2 cytokines. Clin Exp Immunol 1998; 114: 385–391.
Sellon RK, Tonkonogy S, Schultz M, Dieleman LA, Grenther W, Balish E et al. Resident enteric bacteria are necessary for development of spontaneous colitis and immune system activation in interleukin-10-deficient mice. Infect Immun 1998; 66: 5224–5231.
Boivin GP, Washington K, Yang K, Ward JM, Pretlow TP, Russell R et al. Pathology of mouse models of intestinal cancer: consensus report and recommendations. Gastroenterology 2003; 124: 762–777.
Wu S, Liao AP, Xia Y, Li YC, Li JD, Sartor RB et al. Vitamin D receptor negatively regulates bacterial-stimulated NF-kappaB activity in intestine. Am J Pathol 2010; 177: 686–697.
Sun J, Kong J, Duan Y, Szeto FL, Liao A, Madara JL et al. Increased NF-kappaB activity in fibroblasts lacking the vitamin D receptor. Am J Physiol Endocrinol Metab 2006; 291: E315–E322.
Sun J, Mustafi R, Cerda S, Chumsangsri A, Xia YR, Li YC et al. Lithocholic acid down-regulation of NF-kappaB activity through vitamin D receptor in colonic cancer cells. J Steroid Biochem Mol Biol 2008; 111: 37–40.
Zeger SL, Liang KY . Longitudinal data analysis for discrete and continuous outcomes. Biometrics 1986; 42: 121–130.
Acknowledgements
This work was supported by the American Cancer Society Research Scholar Award and NIDDK KO1 DK075386 to JS.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Supplementary Information accompanies this paper on the Oncogenesis website
Rights and permissions
Oncogenesis is an open-access journal published by Nature Publishing Group. This work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 3.0 Unported License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/3.0/
About this article

Cite this article
Lu, R., Wu, S., Zhang, Yg. et al. Enteric bacterial protein AvrA promotes colonic tumorigenesis and activates colonic beta-catenin signaling pathway. Oncogenesis 3, e105 (2014). https://doi.org/10.1038/oncsis.2014.20
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/oncsis.2014.20
This article is cited by
-
Intratumoral microorganisms in tumors of the digestive system
Cell Communication and Signaling (2024)
-
Intratumoral microbiota: roles in cancer initiation, development and therapeutic efficacy
Signal Transduction and Targeted Therapy (2023)
-
Bacterial Peptides and Bacteriocins as Novel Treatment for Prostate Cancer
International Journal of Peptide Research and Therapeutics (2023)
-
A Systemic Review on Fitness and Survival of Salmonella in Dynamic Environment and Conceivable Ways of Its Mitigation
Indian Journal of Microbiology (2023)
