
Abstract
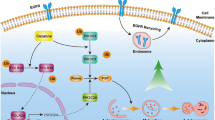
Nucleus accumbens-1 (NAC1), a nuclear factor belonging to the BTB/POZ gene family, is known to have important roles in proliferation and growth of tumor cells and in chemotherapy resistance. Yet, the mechanisms underlying how NAC1 contributes to drug resistance remain largely unclear. We report here that autophagy was involved in NAC1-mediated resistance to cisplatin, a commonly used chemotherapeutic drug in the treatment of ovarian cancer. We found that treatment with cisplatin caused an activation of autophagy in ovarian cancer cell lines, A2780, OVCAR3 and SKOV3. We further demonstrated that knockdown of NAC1 by RNA interference or inactivation of NAC1 by inducing the expression of a NAC1 deletion mutant that contains only the BTB/POZ domain significantly inhibited the cisplatin-induced autophagy, resulting in increased cisplatin cytotoxicity. Moreover, inhibition of autophagy and sensitization to cisplatin by NAC1 knockdown or inactivation were accompanied by induction of apoptosis. To confirm that the sensitizing effect of NAC1 inhibition on the cytotoxicity of cisplatin was attributed to suppression of autophagy, we assessed the effects of the autophagy inhibitors 3-methyladenosine and chloroquine, and small interfering RNAs (siRNAs) targeting beclin 1 or Atg5 on the cytotoxicity of cisplatin. Treatment with 3-methyladenosine, chloroquine or beclin 1 and Atg5-targeted siRNA also enhanced the sensitivity of SKOV3, A2780 and OVCAR3 cells to cisplatin, indicating that suppression of autophagy indeed renders tumor cells more sensitive to cisplatin. Regulation of autophagy by NAC1 was mediated by the high-mobility group box 1 (HMGB1), as the functional status of NAC1 was associated with the expression, translocation and release of HMGB1. The results of our study not only revealed a new mechanism determining cisplatin sensitivity but also identified NAC1 as a novel regulator of autophagy. Thus, the NAC1-mediated autophagy may be exploited as a new target for enhancing the efficacy of cisplatin against ovarian cancer and other types of malignancies.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 50 print issues and online access
$259.00 per year
only $5.18 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout






Similar content being viewed by others
References
Borst P, Rottenberg S, Jonkers J . (2008). How do real tumors become resistant to cisplatin? Cell Cycle 7: 1353–1359.
Cheng Y, Li H, Ren X, Niu T, Hait WN, Yang J . (2010). Cytoprotective effect of the elongation factor-2 kinase-mediated autophagy in breast cancer cells subjected to growth factor inhibition. PLoS One 5: e9715.
Davidson B, Berner A, Trope CG, Wang TL, Shih Ie M . (2007). Expression and clinical role of the bric-a-brac tramtrack broad complex/poxvirus and zinc protein NAC-1 in ovarian carcinoma effusions. Hum Pathol 38: 1030–1036.
Ellerman JE, Brown CK, de Vera M, Zeh HJ, Billiar T, Rubartelli A et al. (2007). Masquerader: high mobility group box-1 and cancer. Clin Cancer Res 13: 2836–2848.
Fanzani A, Zanola A, Rovetta F, Rossi S, Aleo MF . (2011). Cisplatin triggers atrophy of skeletal C2C12 myotubes via impairment of Akt signalling pathway and subsequent increment activity of proteasome and autophagy systems. Toxicol Appl Pharmacol 250: 312–321.
Harhaji-Trajkovic L, Vilimanovich U, Kravic-Stevovic T, Bumbasirevic V, Trajkovic V . (2009). AMPK-mediated autophagy inhibits apoptosis in cisplatin-treated tumour cells. J Cell Mol Med 13: 3644–3654.
He Q, Liang CH, Lippard SJ . (2000). Steroid hormones induce HMG1 overexpression and sensitize breast cancer cells to cisplatin and carboplatin. Proc Natl Acad Sci USA 97: 5768–5772.
Jinawath N, Vasoontara C, Yap KL, Thiaville MM, Nakayama K, Wang TL et al. (2009). NAC-1, a potential stem cell pluripotency factor, contributes to paclitaxel resistance in ovarian cancer through inactivating Gadd45 pathway. Oncogene 28: 1941–1948.
Kaushal GP, Kaushal V, Herzog C, Yang C . (2008). Autophagy delays apoptosis in renal tubular epithelial cells in cisplatin cytotoxicity. Autophagy 4: 710–712.
Kim J, Chu J, Shen X, Wang J, Orkin SH . (2008). An extended transcriptional network for pluripotency of embryonic stem cells. Cell 132: 1049–1061.
Liu D, Yang Y, Liu Q, Wang J . (2009). Inhibition of autophagy by 3-MA potentiates cisplatin-induced apoptosis in esophageal squamous cell carcinoma cells. Med Oncol 28: 105–111.
Liu L, Yang M, Kang R, Wang Z, Zhao Y, Yu Y et al. (2011). HMGB1-induced autophagy promotes chemotherapy resistance in leukemia cells. Leukemia 25: 23–31.
Livesey KM, Tang D, Zeh HJ, Lotze MT . (2009). Autophagy inhibition in combination cancer treatment. Curr Opin Investig Drugs 10: 1269–1279.
Muller FJ, Laurent LC, Kostka D, Ulitsky I, Williams R, Lu C et al. (2008). Regulatory networks define phenotypic classes of human stem cell lines. Nature 455: 401–405.
Nagatani G, Nomoto M, Takano H, Ise T, Kato K, Imamura T et al. (2001). Transcriptional activation of the human HMG1 gene in cisplatin-resistant human cancer cells. Cancer Res 61: 1592–1597.
Nakayama K, Nakayama N, Davidson B, Katabuchi H, Kurman RJ, Velculescu VE et al. (2006a). Homozygous deletion of MKK4 in ovarian serous carcinoma. Cancer Biol Ther 5: 630–634.
Nakayama K, Nakayama N, Davidson B, Sheu JJ, Jinawath N, Santillan A et al. (2006b). A BTB/POZ protein, NAC-1, is related to tumor recurrence and is essential for tumor growth and survival. Proc Natl Acad Sci USA 103: 18739–18744.
Nakayama K, Nakayama N, Wang TL, Shih Ie M . (2007). NAC-1 controls cell growth and survival by repressing transcription of Gadd45GIP1, a candidate tumor suppressor. Cancer Res 67: 8058–8064.
Periyasamy-Thandavan S, Jiang M, Wei Q, Smith R, Yin XM, Dong Z . (2008). Autophagy is cytoprotective during cisplatin injury of renal proximal tubular cells. Kidney Int 74: 631–640.
Ren JH, He WS, Nong L, Zhu QY, Hu K, Zhang RG et al. (2010). Acquired cisplatin resistance in human lung adenocarcinoma cells is associated with enhanced autophagy. Cancer Biother Radiopharm 25: 75–80.
Sharma A, Ramanjaneyulu A, Ray R, Rajeswari MR . (2009). Involvement of high mobility group B proteins in cisplatin-induced cytotoxicity in squamous cell carcinoma of skin. DNA Cell Biol 28: 311–318.
Sun Y, Liu JH, Jin L, Lin SM, Yang Y, Sui YX et al. (2010). Over-expression of the Beclin1 gene upregulates chemosensitivity to anti-cancer drugs by enhancing therapy-induced apoptosis in cervix squamous carcinoma CaSki cells. Cancer Lett 294: 204–210.
Tang D, Kang R, Cheh CW, Livesey KM, Liang X, Schapiro NE et al. (2010a). HMGB1 release and redox regulates autophagy and apoptosis in cancer cells. Oncogene 29: 5299–5310.
Tang D, Kang R, Livesey KM, Cheh CW, Farkas A, Loughran P et al. (2010b). Endogenous HMGB1 regulates autophagy. J Cell Biol 190: 881–892.
Thorburn J, Frankel AE, Thorburn A . (2009a). Regulation of HMGB1 release by autophagy. Autophagy 5: 247–249.
Thorburn J, Horita H, Redzic J, Hansen K, Frankel AE, Thorburn A . (2009b). Autophagy regulates selective HMGB1 release in tumor cells that are destined to die. Cell Death Differ 16: 175–183.
Wang J, Rao S, Chu J, Shen X, Levasseur DN, Theunissen TW et al. (2006). A protein interaction network for pluripotency of embryonic stem cells. Nature 444: 364–368.
Wang J, Zhou JY, Wu GS . (2007). ERK-dependent MKP-1-mediated cisplatin resistance in human ovarian cancer cells. Cancer Res 67: 11933–11941.
Wei M, Burenkova O, Lippard SJ . (2003). Cisplatin sensitivity in Hmbg1-/- and Hmbg1+/+ mouse cells. J Biol Chem 278: 1769–1773.
White E, DiPaola RS . (2009). The double-edged sword of autophagy modulation in cancer. Clin Cancer Res 15: 5308–5316.
Wu H, Zhu H, Liu DX, Niu TK, Ren X, Patel R et al. (2009). Silencing of elongation factor-2 kinase potentiates the effect of 2-deoxy-D-glucose against human glioma cells through blunting of autophagy. Cancer Res 69: 2453–2460.
Yeasmin S, Nakayama K, Ishibashi M, Katagiri A, Iida K, Purwana IN et al. (2008). Expression of the bric-a-brac tramtrack broad complex protein NAC-1 in cervical carcinomas seems to correlate with poorer prognosis. Clin Cancer Res 14: 1686–1691.
Acknowledgements
This study was supported by grants from the US Public Health Service (R01CA135038 and RO1CA103937) and the Penn State College of Medicine
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Supplementary Information accompanies the paper on the Oncogene website
Rights and permissions
About this article
Cite this article
Zhang, Y., Cheng, Y., Ren, X. et al. NAC1 modulates sensitivity of ovarian cancer cells to cisplatin by altering the HMGB1-mediated autophagic response. Oncogene 31, 1055–1064 (2012). https://doi.org/10.1038/onc.2011.290
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/onc.2011.290
Keywords
This article is cited by
-
HERPUD1 promotes ovarian cancer cell survival by sustaining autophagy and inhibit apoptosis via PI3K/AKT/mTOR and p38 MAPK signaling pathways
BMC Cancer (2022)
-
miR-18a-5p derived from mesenchymal stem cells-extracellular vesicles inhibits ovarian cancer cell proliferation, migration, invasion, and chemotherapy resistance
Journal of Translational Medicine (2022)
-
FUS-induced circRHOBTB3 facilitates cell proliferation via miR-600/NACC1 mediated autophagy response in pancreatic ductal adenocarcinoma
Journal of Experimental & Clinical Cancer Research (2021)
-
Autophagy modulating agents as chemosensitizers for cisplatin therapy in cancer
Investigational New Drugs (2021)
-
Autophagy inhibition enhances PD-L1 expression in gastric cancer
Journal of Experimental & Clinical Cancer Research (2019)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}