
Key Points
-
The ultimate goal of regenerative medicine is to replace lost or damaged cells. This can potentially be accomplished using the processes of dedifferentiation, transdifferentiation or reprogramming.
-
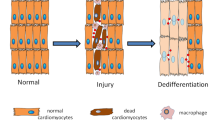
During dedifferentiation, a terminally differentiated cell reverts back to a less-differentiated stage from within its own lineage, which allows it to proliferate. Many regenerative processes have been associated with dedifferentiation.
-
Transdifferentiation sees cells regress to a point when they can switch lineages or can also occur directly between two different cell types.
-
Reprogramming aims to induce differentiated cells into reverting to pluripotency. From here, they can differentiate into almost any cell type.
-
During dedifferentiation and transdifferentiation, well-defined intermediate cell types have been identified. The process of reprogramming seems to be largely stochastic.
-
Dedifferentiation and transdifferentiation can be successfully achieved in vivo, and reprogramming facilitates genetic manipulation such as correcting disease-inducing mutations.
Abstract
The ultimate goal of regenerative medicine is to replace lost or damaged cells. This can potentially be accomplished using the processes of dedifferentiation, transdifferentiation or reprogramming. Recent advances have shown that the addition of a group of genes can not only restore pluripotency in a fully differentiated cell state (reprogramming) but can also induce the cell to proliferate (dedifferentiation) or even switch to another cell type (transdifferentiation). Current research aims to understand how these processes work and to eventually harness them for use in regenerative medicine.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$189.00 per year
only $15.75 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others


References
Brockes, J. P. & Kumar, A. Plasticity and reprogramming of differentiated cells in amphibian regeneration. Nature Rev. Mol. Cell Biol. 3, 566–574 (2002).
Wolff, G. Entwicklungsphysiologische Studien. I. Die Regeneration der. Urodelenlinse. Wilhelm Roux' Arch. Entwickl-Mech. Org. 1, 380–390 (1895).
de Vries, W. N. et al. Reprogramming and differentiation in mammals: motifs and mechanisms. Cold Spring Harb. Symp. Quant. Biol. 73, 33–38 (2008).
Poss, K. D., Wilson, L. G. & Keating, M. T. Heart regeneration in zebrafish. Science 298, 2188–2190 (2002).
Raya, A. et al. Activation of Notch signaling pathway precedes heart regeneration in zebrafish. Proc. Natl Acad. Sci. USA 100, 11889–11895 (2003).
Jopling, C. et al. Zebrafish heart regeneration occurs by cardiomyocyte dedifferentiation and proliferation. Nature 464, 606–609 (2010).
Kikuchi, K. et al. Primary contribution to zebrafish heart regeneration by gata4+ cardiomyocytes. Nature 464, 601–605 (2010).
Sleep, E. et al. Transcriptomics approach to investigate zebrafish heart regeneration. J. Cardiovasc. Med. 11, 369–380 (2010).
Lien, C. L., Schebesta, M., Makino, S., Weber, G. J. & Keating, M. T. Gene expression analysis of zebrafish heart regeneration. PLoS Biol. 4, e260 (2006).
Lepilina, A. et al. A dynamic epicardial injury response supports progenitor cell activity during zebrafish heart regeneration. Cell 127, 607–619 (2006).
Ahuja, P., Sdek, P. & MacLellan, W. R. Cardiac myocyte cell cycle control in development, disease, and regeneration. Physiol. Rev. 87, 521–544 (2007).
Poss, K. D. Advances in understanding tissue regenerative capacity and mechanisms in animals. Nature Rev. Genet. 11, 710–722 (2010).
Burkhart, D. L. & Sage, J. Cellular mechanisms of tumour suppression by the retinoblastoma gene. Nature Rev. Cancer 8, 671–682 (2008).
Tanaka, E. M., Gann, A. A., Gates, P. B. & Brockes, J. P. Newt myotubes reenter the cell cycle by phosphorylation of the retinoblastoma protein. J. Cell Biol. 136, 155–165 (1997).
Nye, H. L., Cameron, J. A., Chernoff, E. A. & Stocum, D. L. Regeneration of the urodele limb: a review. Dev. Dyn. 226, 280–294 (2003).
Kim, K. et al. Epigenetic memory in induced pluripotent stem cells. Nature 467, 285–290 (2010).
Kragl, M. et al. Cells keep a memory of their tissue origin during axolotl limb regeneration. Nature 460, 60–65 (2009).
Chen, Z. L., Yu, W. M. & Strickland, S. Peripheral regeneration. Annu. Rev. Neurosci. 30, 209–233 (2007).
Mirsky, R. et al. Novel signals controlling embryonic Schwann cell development, myelination and dedifferentiation. J. Peripher. Nerv. Syst. 13, 122–135 (2008).
Woodhoo, A. et al. Notch controls embryonic Schwann cell differentiation, postnatal myelination and adult plasticity. Nature Neurosci. 12, 839–847 (2009).
Wolff, T. & Ready, D. F. The beginning of pattern formation in the Drosophila compound eye: the morphogenetic furrow and the second mitotic wave. Development 113, 841–850 (1991).
Harvey, K. & Tapon, N. The Salvador–Warts–Hippo pathway — an emerging tumour-suppressor network. Nature Rev. Cancer 7, 182–191 (2007).
Nicolay, B. N., Bayarmagnai, B., Moon, N. S., Benevolenskaya, E. V. & Frolov, M. V. Combined inactivation of pRB and Hippo pathways induces dedifferentiation in the Drosophila retina. PLoS Genet. 6, e1000918 (2010).
Monje, P. V., Soto, J., Bacallao, K. & Wood, P. M. Schwann cell dedifferentiation is independent of mitogenic signaling and uncoupled to proliferation: role of cAMP and JNK in the maintenance of the differentiated state. J. Biol. Chem. 285, 31024–31036 (2010).
McGann, C. J., Odelberg, S. J. & Keating, M. T. Mammalian myotube dedifferentiation induced by newt regeneration extract. Proc. Natl Acad. Sci. USA 98, 13699–13704 (2001).
Camarda, G. et al. A pRb-independent mechanism preserves the postmitotic state in terminally differentiated skeletal muscle cells. J. Cell Biol. 167, 417–423 (2004).
Sherr, C. J. et al. p53-dependent and -independent functions of the Arf tumor suppressor. Cold Spring Harb. Symp. Quant. Biol. 70, 129–137 (2005).
Pajcini, K. V., Corbel, S. Y., Sage, J., Pomerantz, J. H. & Blau, H. M. Transient inactivation of Rb and ARF yields regenerative cells from postmitotic mammalian muscle. Cell Stem Cell 7, 198–213 (2010).
Bicknell, K. A., Coxon, C. H. & Brooks, G. Can the cardiomyocyte cell cycle be reprogrammed? J. Mol. Cell Cardiol. 42, 706–721 (2007).
MacLellan, W. R. et al. Overlapping roles of pocket proteins in the myocardium are unmasked by germ line deletion of p130 plus heart-specific deletion of Rb. Mol. Cell Biol. 25, 2486–2497 (2005).
Lee, J. et al. Activation of p38 MAPK induces cell cycle arrest via inhibition of Raf/ERK pathway during muscle differentiation. Biochem. Biophys. Res. Commun. 298, 765–771 (2002).
Engel, F. B. et al. p38 MAP kinase inhibition enables proliferation of adult mammalian cardiomyocytes. Genes Dev. 19, 1175–1187 (2005).
Rumyantsev, P. P. Interrelations of the proliferation and differentiation processes during cardiact myogenesis and regeneration. Int. Rev. Cytol. 51, 186–273 (1977).
Gassmann, M. et al. Aberrant neural and cardiac development in mice lacking the ErbB4 neuregulin receptor. Nature 378, 390–394 (1995).
Lee, K. F. et al. Requirement for neuregulin receptor ErbB2 in neural and cardiac development. Nature 378, 394–398 (1995).
Meyer, D. & Birchmeier, C. Multiple essential functions of neuregulin in development. Nature 378, 386–390 (1995).
Bersell, K., Arab, S., Haring, B. & Kühn, B. Neuregulin1/ErbB4 signaling induces cardiomyocyte proliferation and repair of heart injury. Cell 138, 257–270 (2009).
Tsonis, P. A., Madhavan, M., Tancous, E. E. & Del Rio-Tsonis, K. A newt's eye view of lens regeneration. Int. J. Dev. Biol. 48, 975–980 (2004).
Thitoff, A. R., Call, M. K., Del Rio-Tsonis, K. & Tsonis, P. A. Unique expression patterns of the retinoblastoma (Rb) gene in intact and lens regeneration-undergoing newt eyes. Anat. Rec. A Discov. Mol. Cell. Evol. Biol. 271, 185–188 (2003).
Maki, N. et al. Expression profiles during dedifferentiation in newt lens regeneration revealed by expressed sequence tags. Mol. Vis. 16, 72–78 (2010).
Tsonis, P. A. et al. Effects of a CDK inhibitor on lens regeneration. Wound Repair Regen. 12, 24–29 (2004).
Xie, H., Ye, M., Feng, R. & Graf, T. Stepwise reprogramming of B cells into macrophages. Cell 117, 663–676 (2004).
McKercher, S. R. et al. Targeted disruption of the PU.1 gene results in multiple hematopoietic abnormalities. EMBO J. 15, 5647–5658 (1996).
Nutt, S. L., Heavey, B., Rolink, A. G. & Busslinger, M. Commitment to the B-lymphoid lineage depends on the transcription factor Pax5. Nature 401, 556–562 (1999).
Offield, M. F. et al. PDX-1 is required for pancreatic outgrowth and differentiation of the rostral duodenum. Development 122, 983–995 (1996).
Meivar-Levy, I. et al. Pancreatic and duodenal homeobox gene 1 induces hepatic dedifferentiation by suppressing the expression of CCAAT/enhancer-binding protein β. Hepatology 46, 898–905 (2007).
Zhou, Q., Brown, J., Kanarek, A., Rajagopal, J. & Melton, D. A. In vivo reprogramming of adult pancreatic exocrine cells to β-cells. Nature 455, 627–632 (2008).
Gradwohl, G., Dierich, A., LeMeur, M. & Guillemot, F. neurogenin3 is required for the development of the four endocrine cell lineages of the pancreas. Proc. Natl Acad. Sci. USA 97, 1607–1611 (2000).
Bernardo, A. S., Hay, C. W. & Docherty, K. Pancreatic transcription factors and their role in the birth, life and survival of the pancreatic β cell. Mol. Cell. Endocrinol. 294, 1–9 (2008).
Kataoka, K. et al. MafA is a glucose-regulated and pancreatic β-cell-specific transcriptional activator for the insulin gene. J. Biol. Chem. 277, 49903–49910 (2002).
Thorel, F. et al. Conversion of adult pancreatic α-cells to β-cells after extreme β-cell loss. Nature 464, 1149–1154 (2010).
Vierbuchen, T. et al. Direct conversion of fibroblasts to functional neurons by defined factors. Nature 463, 1035–1041 (2010).
Romm, E., Nielsen, J. A., Kim, J. G. & Hudson, L. D. Myt1 family recruits histone deacetylase to regulate neural transcription. J. Neurochem. 93, 1444–1453 (2005).
Ieda, M. et al. Direct reprogramming of fibroblasts into functional cardiomyocytes by defined factors. Cell 142, 375–386 (2010).
Ghosh, T. K. et al. Physical interaction between TBX5 and MEF2C is required for early heart development. Mol. Cell. Biol. 29, 2205–2218 (2009).
Garg, V. et al. GATA4 mutations cause human congenital heart defects and reveal an interaction with TBX5. Nature 424, 443–447 (2003).
Lin, Q., Schwarz, J., Bucana, C. & Olson, E. N. Control of mouse cardiac morphogenesis and myogenesis by transcription factor MEF2C. Science 276, 1404–1407 (1997).
Cirillo, L. A. et al. Opening of compacted chromatin by early developmental transcription factors HNF3 (FoxA) and GATA-4. Mol. Cell 9, 279–289 (2002).
Yamanaka, S. & Blau, H. M. Nuclear reprogramming to a pluripotent state by three approaches. Nature 465, 704–712 (2010).
Takahashi, K. & Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 126, 663–676 (2006).
van den Berg, D. L. et al. An Oct4-centered protein interaction network in embryonic stem cells. Cell Stem Cell 6, 369–381 (2010).
Pardo, M. et al. An expanded Oct4 interaction network: implications for stem cell biology, development, and disease. Cell Stem Cell 6, 382–395 (2010).
Nichols, J. et al. Formation of pluripotent stem cells in the mammalian embryo depends on the POU transcription factor Oct4. Cell 95, 379–391 (1998).
Ellis, P. et al. SOX2, a persistent marker for multipotential neural stem cells derived from embryonic stem cells, the embryo or the adult. Dev. Neurosci. 26, 148–165 (2004).
Kondoh, H. & Kamachi, Y. SOX-partner code for cell specification: regulatory target selection and underlying molecular mechanisms. Int. J. Biochem. Cell Biol. 42, 391–399 (2010).
Masui, S. et al. Pluripotency governed by Sox2 via regulation of Oct3/4 expression in mouse embryonic stem cells. Nature Cell Biol. 9, 625–635 (2007).
Niwa, H., Miyazaki, J. & Smith, A. G. Quantitative expression of Oct-3/4 defines differentiation, dedifferentiation or self-renewal of ES cells. Nature Genet. 24, 372–376 (2000).
Silva, J. et al. Nanog is the gateway to the pluripotent ground state. Cell 138, 722–737 (2009).
Chambers, I. et al. Nanog safeguards pluripotency and mediates germline development. Nature 450, 1230–1234 (2007).
Jaenisch, R. & Young, R. Stem cells, the molecular circuitry of pluripotency and nuclear reprogramming. Cell 132, 567–582 (2008).
Niwa, H. How is pluripotency determined and maintained? Development 134, 635–646 (2007).
Loh, Y. H. et al. The Oct4 and Nanog transcription network regulates pluripotency in mouse embryonic stem cells. Nature Genet. 38, 431–440 (2006).
Kuroda, T. et al. Octamer and Sox elements are required for transcriptional cis regulation of Nanog gene expression. Mol. Cell. Biol. 25, 2475–2485 (2005).
Kashyap, V. et al. Regulation of stem cell pluripotency and differentiation involves a mutual regulatory circuit of the NANOG, OCT4, and SOX2 pluripotency transcription factors with polycomb repressive complexes and stem cell microRNAs. Stem Cells Dev. 18, 1093–1108 (2009).
Hochedlinger, K. & Plath, K. Epigenetic reprogramming and induced pluripotency. Development 136, 509–523 (2009).
Sridharan, R. et al. Role of the murine reprogramming factors in the induction of pluripotency. Cell 136, 364–377 (2009).
Nakagawa, M. et al. Generation of induced pluripotent stem cells without Myc from mouse and human fibroblasts. Nature Biotech. 26, 101–106 (2008).
Giorgetti, A. et al. Generation of induced pluripotent stem cells from human cord blood using OCT4 and SOX2. Cell Stem Cell 5, 353–357 (2009).
Kim, J. B. et al. Direct reprogramming of human neural stem cells by OCT4. Nature 461, 649–643 (2009).
Heng, J. C. et al. The nuclear receptor Nr5a2 can replace Oct4 in the reprogramming of murine somatic cells to pluripotent cells. Cell Stem Cell 6, 167–174 (2010).
Hanna, J. et al. Direct cell reprogramming is a stochastic process amenable to acceleration. Nature 462, 595–601 (2009).
Mikkelsen, T. S. et al. Dissecting direct reprogramming through integrative genomic analysis. Nature 454, 49–55 (2008).
Lowry, W. E. et al. Generation of human induced pluripotent stem cells from dermal fibroblasts. Proc. Natl Acad. Sci. USA 105, 2883–2888 (2008).
Ying, Q. L. et al. The ground state of embryonic stem cell self-renewal. Nature 453, 519–523 (2008).
Smith, Z. D., Nachman, I., Regev, A. & Meissner, A. Dynamic single-cell imaging of direct reprogramming reveals an early specifying event. Nature Biotech. 28, 521–526 (2010).
Stadtfeld, M., Maherali, N., Breault, D. T. & Hochedlinger, K. Defining molecular cornerstones during fibroblast to iPS cell reprogramming in mouse. Cell Stem Cell 2, 230–240 (2008).
Huangfu, D. et al. Induction of pluripotent stem cells by defined factors is greatly improved by small-molecule compounds. Nature Biotech. 26, 795–797 (2008).
Wernig, M. et al. A drug-inducible transgenic system for direct reprogramming of multiple somatic cell types. Nature Biotech. 26, 916–924 (2008).
Osawa, M., Hanada, K., Hamada, H. & Nakauchi, H. Long-term lymphohematopoietic reconstitution by a single CD34-low/negative hematopoietic stem cell. Science 273, 242–245 (1996).
Yin, T. & Li, L. The stem cell niches in bone. J. Clin. Invest. 116, 1195–1201 (2006).
Arai, F. et al. Tie2/angiopoietin-1 signaling regulates hematopoietic stem cell quiescence in the bone marrow niche. Cell 118, 149–161 (2004).
Wilson, A. et al. Dormant and self-renewing hematopoietic stem cells and their niches. Ann. N. Y. Acad. Sci. 1106, 64–75 (2007).
Kopp, H. G., Avecilla, S. T., Hooper, A. T. & Rafii, S. The bone marrow vascular niche: home of HSC differentiation and mobilization. Physiology 20, 349–356 (2005).
Flanagan, J. G., Chan, D. C. & Leder, P. Transmembrane form of the kit ligand growth factor is determined by alternative splicing and is missing in the Sld mutant. Cell 64, 1025–1035 (1991).
Broudy, V. C. Stem cell factor and hematopoiesis. Blood 90, 1345–1364 (1997).
Heissig, B. et al. Recruitment of stem and progenitor cells from the bone marrow niche requires MMP-9 mediated release of kit-ligand. Cell 109, 625–637 (2002).
Weber, J. M. & Calvi, L. M. Notch signaling and the bone marrow hematopoietic stem cell niche. Bone 46, 281–285 (2010).
Meshorer, E. & Misteli, T. Chromatin in pluripotent embryonic stem cells and differentiation. Nature Rev. Mol. Cell Biol. 7, 540–546 (2006).
Mallanna, S. K. & Rizzino, A. Emerging roles of microRNAs in the control of embryonic stem cells and the generation of induced pluripotent stem cells. Dev. Biol. 344, 16–25 (2010).
Boyer, L. A. et al. Polycomb complexes repress developmental regulators in murine embryonic stem cells. Nature 441, 349–353 (2006).
Fouse, S. D. et al. Promoter CpG methylation contributes to ES cell gene regulation in parallel with Oct4/Nanog, PcG complex, and histone H3 K4/K27 trimethylation. Cell Stem Cell 2, 160–169 (2008).
Francis, N. J., Kingston, R. E. & Woodcock, C. L. Chromatin compaction by a polycomb group protein complex. Science 306, 1574–1577 (2004).
Nagy, P. L., Griesenbeck, J., Kornberg, R. D. & Cleary, M. L. A trithorax-group complex purified from Saccharomyces cerevisiae is required for methylation of histone H3. Proc. Natl Acad. Sci. USA 99, 90–94 (2002).
Mikkelsen, T. S. et al. Genome-wide maps of chromatin state in pluripotent and lineage-committed cells. Nature 448, 553–560 (2007).
Zhao, X. D. et al. Whole-genome mapping of histone H3 Lys4 and 27 trimethylations reveals distinc genomic compartments in human embryonic stem cells. Cell Stem Cell 1, 286–298 (2007).
Lister, R. et al. Human DNA methylomes at base resolution show widespread epigenomic differences. Nature 462, 315–322 (2009).
Loh, Y. H., Zhang, W., Chen, X., George, J. & Ng, H. H. Jmjd1a and Jmjd2c histone H3 Lys 9 demethylases regulate self-renewal in embryonic stem cells. Genes Dev. 21, 2545–2557 (2007).
Kloosterman, W. P. & Plasterk, R. H. The diverse functions of microRNAs in animal development and disease. Dev. Cell 11, 441–450 (2006).
Sinkkonen, L. et al. MicroRNAs control de novo DNA methylation through regulation of transcriptional repressors in mouse embryonic stem cells. Nature Struct. Mol. Biol. 15, 259–267 (2008).
Acknowledgements
We apologize to our colleagues whose research could not be included in this article owing to size limitations. We thank A. Faucherre and M. J. Barrero for constructive criticism of the manuscript and the members of the laboratory for discussions. Work in the laboratory of J.C.I.B. was supported by grants from MICINN, Fundacion Cellex, Sanofi-Aventis and the G. Harold and Leila Y. Mathers Charitable Foundation.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
Chris Jopling, Stephanie Boue and Juan Carlos Izpisua Belmonte
Dedifferentiation, transdifferentiation and reprogramming: three routes to regeneration
The authors have received funding from Sanofi-Aventis.
Related links
Glossary
- Pluripotency
-
The ability of a cell to give rise to all cells of the embryo. Cells of the inner cell mass and its derivative embryonic stem cells are pluripotent.
- Totipotent
-
Pertaining to the ability of a cell to give rise to all cells of an organism, including embryonic and extra-embryonic tissues. Zygotes are totipotent.
- Hyperplasia
-
Growth that is due to an increase in the number of cells.
- Hypertrophic growth
-
Growth that is due to an increase in the size of cells.
- Multipotent
-
Pertaining to the ability of a cell to give rise to different cell types of a given cell lineage. Most adult stem cells, such as gut stem cells, skin stem cells, haematopoietic stem cells and neural stem cells, are multipotent.
- Schwann cell
-
A cell type that wraps around the axons of peripheral nerves and forms the insulating myelin sheath.
- Neural crest cell
-
A cell type that migrates during neurulation and forms most of the peripheral nervous system (and many other structures) in the embryo.
- Myotube
-
An elongated multinucleate cell (with three or more nuclei) that contains some peripherally located myofibrils. Myotubes are formed in vivo or in vitro by the fusion of myoblasts; they eventually develop into mature muscle fibres that have peripherally located nuclei and most of their cytoplasm filled with myofibrils.
- β-cell
-
A cell type in the pancreas, found in the islets of Langerhans, that produces insulin.
- Pancreatic exocrine cell
-
A pancreatic cell type that is responsible for the secretion of bicarbonate ions and digestive enzymes.
- DNA methyltransferase
-
An enzyme that catalyses the addition of a methyl group to C or A. DNMT1 is a maintenance DNA methyltransferase, DNMT3a and DNMT3b are de novo DNA methyltransferases.
Rights and permissions
About this article
Cite this article
Jopling, C., Boue, S. & Belmonte, J. Dedifferentiation, transdifferentiation and reprogramming: three routes to regeneration. Nat Rev Mol Cell Biol 12, 79–89 (2011). https://doi.org/10.1038/nrm3043
Published:
Issue Date:
DOI: https://doi.org/10.1038/nrm3043
This article is cited by
-
Tissue-engineered bone construct promotes early osseointegration of implants with low primary stability in oversized osteotomy
BMC Oral Health (2024)
-
Epigenetic OCT4 regulatory network: stochastic analysis of cellular reprogramming
npj Systems Biology and Applications (2024)
-
Homeostatic regulation of renewing tissue cell populations via crowding control: stability, robustness and quasi-dedifferentiation
Journal of Mathematical Biology (2024)
-
Current view of liver cancer cell-of-origin and proposed mechanisms precluding its proper determination
Cancer Cell International (2023)
-
Myc-dependent dedifferentiation of Gata6+ epidermal cells resembles reversal of terminal differentiation
Nature Cell Biology (2023)
