
Key Points
-
Apoptosis is characterized by a series of dramatic perturbations to the cellular architecture that contribute not only to cell death, but also prepare cells for removal by phagocytes.
-
Apoptotic cell death is orchestrated by members of the caspase family of cysteine proteases. Caspases target several hundred proteins for restricted proteolysis during this complex process of cell demolition.
-
Although numerous caspase substrates have been linked to specific morphological features of apoptosis, it is likely that many proteins targeted by caspases contribute to death of the cell but have no significant impact on the phenotypic changes that are seen during this process. Because proteins involved in diverse cell functions are cleaved by caspases, it appears that these enzymes use a 'death by a thousand cuts' strategy to ensure that a cell dies. In other words, death by caspase activation probably results from many potentially lethal injuries rather than a single fatal blow.
-
Multiple membrane alterations, most notably the externalization of phosphatidylserine, have been detected on apoptotic cells. These membrane alterations trigger the recognition and engulfment of apoptotic cells by phagocytes.
-
Because apoptotic cells are typically recognized and engulfed by macrophages before leakage of their intracellular contents, apoptosis typically precludes the release of immunostimulatory molecules (called danger signals) that would not normally be present in the extracellular space. Therefore, apart from limiting direct cell damage caused by the release of cytoplasmic contents, one of the main benefits of controlled cell death through apoptosis may be to prevent unwanted immune responses.
Abstract
Apoptosis is characterized by a series of dramatic perturbations to the cellular architecture that contribute not only to cell death, but also prepare cells for removal by phagocytes and prevent unwanted immune responses. Much of what happens during the demolition phase of apoptosis is orchestrated by members of the caspase family of cysteine proteases. These proteases target several hundred proteins for restricted proteolysis in a controlled manner that minimizes damage and disruption to neighbouring cells and avoids the release of immunostimulatory molecules.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$189.00 per year
only $15.75 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others


References
Kerr, J. F. R., Wyllie, A. H. & Currie, A. R. Apoptosis: a basic biological phenomenon with wide-ranging implications in tissue kinetics. Br. J. Cancer 26, 239–275 (1972). This seminal study introduced the term apoptosis, providing a detailed description of the features of apoptotic cells and some of the biological contexts in which this mode of cell death is observed.
Wyllie, A. H., Kerr, J. F. R. & Currie, A. R. Cell death: the significance of apoptosis. Int. Rev. Cytol. 68, 251–305 (1980).
Nicholson, D. W. Caspase structure, proteolytic substrates and function during apoptotic death. Cell Death Differ. 6, 1028–1042 (1999).
Creagh, E. M., Conroy, H. & Martin, S. J. Caspase-activation pathways in apoptosis and immunity. Immunol. Rev. 193, 10–21 (2003).
Savill, J. & Fadok, V. Corpse clearance defines the meaning of cell death. Nature 407, 784–788 (2000).
Robertson, A. M., Bird, C. C., Waddell, A. W. & Currie, A. R. Morphological aspects of glucocorticoid-induced cell death in human lymphoblastoid cells. J. Pathol. 126, 181–187 (1978).
Martin, S. J., Bradley, J. G. & Cotter, T. G. HL-60 cells induced to differentiate towards neutrophils subsequently die via apoptosis. Clin. Exp. Immunol. 79, 448–453 (1990).
Williams, J. R., Little, J. B. & Shipley, W. U. Association of mammalian cell death with a specific endonucleolytic degradation of DNA. Nature 252, 754–755 (1974).
Frank, S. et al. The role of dynamin-related protein 1, a mediator of mitochondrial fission, in apoptosis. Dev. Cell 1, 515–525 (2001).
Lane, J. D. et al. Caspase-mediated cleavage of the stacking protein GRASP65 is required for Golgi fragmentation during apoptosis. J. Cell Biol. 156, 495–509 (2002).
Lüthi, A. U. & Martin, S. J. The CASBAH: a searchable database of caspase substrates. Cell Death Differ. 14, 641–650 (2007). A comprehensive survey of the ∼400 caspase substrates identified to date (which can be found in the caspase substrate database .
Gallucci, S., Lolkema, M. & Matzinger, P. Natural adjuvants: endogenous activators of dendritic cells. Nature Med. 5, 1249–1255 (1999). A provocative study that provides evidence that necrotic, but not apoptotic, cells activate dendritic cells.
Shi, Y., Zheng, W. & Rock, K. L. Cell injury releases endogenous adjuvants that stimulate cytotoxic T cell responses. Proc. Natl Acad. Sci. USA 97, 14590–14595 (2000).
Shi, Y., Evans, J. E. & Rock, K. L. Molecular identification of a danger signal that alerts the immune system to dying cells. Nature 425, 516–521 (2003).
Oppenheim, J. J. & Yang, D. Alarmins: chemotactic activators of immune responses. Curr. Opin. Immunol. 17, 359–365 (2005).
Chen, C. J. et al. Identification of a key pathway required for the sterile inflammatory response triggered by dying cells. Nature Med. 13, 851–856 (2007).
Trinchieri, G. & Sher, A. Cooperation of Toll-like receptor signals in innate immune defence. Nature Rev. Immunol. 7, 179–190 (2007).
Matzinger P. The danger model: a renewed sense of self. Science 296, 301–305 (2002). An excellent update of the 'danger model' by its originator.
Scaffidi, P., Misteli, T. & Bianchi, M. E. Release of chromatin protein HMGB1 by necrotic cells triggers inflammation. Nature 418, 191–195 (2002).
Stennicke, H. R. & Salvesen, G. S. Properties of the caspases. Biochim. Biophys Acta. 1387, 17–31 (1998). An excellent overview on the biochemical characteristics of caspases.
Stennicke, H. R. et al. Pro-caspase-3 is a major physiologic target of caspase-8. J. Biol. Chem. 273, 27084–27090 (1998).
Adrain, C. & Martin, S. J. The mitochondrial apoptosome: a killer unleashed by the cytochrome seas. Trends Biochem. Sci. 26, 390–397 (2001).
Communal, C. et al. Functional consequences of caspase activation in cardiac myocytes. Proc. Natl Acad. Sci. USA 99, 6252–6256 (2002).
Gerner, C. et al. The Fas-induced apoptosis analysed by high throughput proteome analysis. J. Biol. Chem. 275, 39018–39026 (2000).
Martin, S. J. et al. Proteolysis of fodrin (non-erythroid spectrin) during apoptosis. J. Biol. Chem. 270, 6425–6428 (1995).
Thiede, B., Treumann, A., Kretschmer, A., Sohike, J. & Rudel, T. Shotgun proteome analysis of protein changes in apoptotic cells. Proteomics 5, 2123–2130 (2005).
Kothakota, S. et al. Caspase-3-generated fragment of gelsolin: effector of morphological change in apoptosis. Science 278, 294–298 (1997).
Browne, K. A., Johnstone, R. W., Jans, D. A. & Trapani, J. A. Filamin (280 kDa actin-binding protein) is a caspase substrate and is also cleaved directly by the cytotoxic T lymphocyte protease granzyme B during apoptosis. J. Biol. Chem. 275, 39262–39266 (2000).
Adrain, C., Brumatti, G., Duriez, P., Delivani, P. & Martin, S. J. The cytotoxic lymphocyte protease, granzyme B, targets the cytoskeleton and perturbs microtubule polymerization dynamics. J. Biol. Chem. 281, 8118–8125 (2006).
Canu, N. et al. Tau cleavage and dephosphorylation in cerebellar granule neurons undergoing apoptosis. J. Neurosci. 18, 7061–7064 (1998).
Lane, J. D., Vergnolle, M. A. S., Woodman, P. G. & Allan, V. J. Apoptotic cleavage of cytoplasmic dynein intermediate chain and p150Glued stops dynein-dependent membrane motility. J. Cell Biol. 153, 1415–1426 (2001).
Morishima, N. Changes in vimentin morphology during apoptosis correlate with vimentin cleavage by different caspases located either upstream or downstream of Bcl-2 action. Genes Cells 4, 401–414 (1999).
Ku, N.-O., Liao, J. & Omary, M. B. Apoptosis generates stable fragments of human type I keratins. J. Biol. Chem. 272, 33197–33203 (1997).
Orth, K., Chinnaiyan, A. M., Garg, M., Froelich, C. J. & Dixit, V. M. The CED-3/ICE-like protease Mch2 is activated during apoptosis and cleaves the death substrate lamin A. J. Biol. Chem. 271, 16443–16446 (1996).
Cotter, T. G., Lennon, S. V., Glynn, J. M. & Green, D. R. Microfilament-disrupting agents prevent the formation of apoptotic bodies in tumour cells undergoing apoptosis. Cancer Res. 52, 997–1005 (1992). The first study to show that actin microfilaments are required for the formation of apoptotic bodies.
Coleman, M. L. et al. Membrane blebbing during apoptosis results from caspase-mediated activation of ROCK I. Nature Cell Biol. 3, 339–345 (2001).
Sebbagh, M. et al. Caspase-3-mediated cleavage of ROCK I induces MLC phosphorylation and apoptotic membrane blebbing. Nature Cell Biol. 3, 346–352 (2001). This study, along with reference 36, strongly implicates ROCK1 in apoptosis-associated membrane blebbing.
Rosenblatt, J., Raff, M.C. & Cramer, L.P. An epithelial cell destined for apoptosis signals its neighbors to extrude it by an actin- and myosin-dependent mechanism. Curr. Biol. 11, 1847–1857 (2001).
Rao, L., Perez, D. & White, E. Lamin proteolysis facilitates nuclear events during apoptosis. J. Cell Biol. 135, 1441–1455 (1996).
Croft, D. R. et al. Actin-myosin-based contraction is responsible for apoptotic nuclear disintegration. J. Cell Biol. 168, 245–255 (2005).
Moss, D. K., Betin, V. M., Malesinski, S. D. & Lane, J. D. A novel role for microtubules in apoptotic chromatin dynamics and cellular fragmentation. J. Cell Sci. 119, 2362–2374 (2006).
Levkau, B., Hrren, B., Koyama, H., Ross, R. & Raines, E. W. Caspase-mediated cleavage of focal adhesion kinase pp125FAK and disassembly of focal adhesions in human endothelial cell apoptosis. J. Exp. Med. 187, 579–586 (1998).
Kim, W., Kook, S., Kim, D. J., Teodorof, C. & Song, W. K. The 31 kDa caspase-generate cleavage product of p130cas functions as a transcriptional repressor of E2A in apoptotic cells. J. Biol. Chem. 279, 8333–8342 (2004).
Kook, S. et al. Caspase-dependent cleavage of tensin induces disruption of actin cytoskeleton during apoptosis. Biochem. Biophys. Res. Comm. 303, 37–45 (2003).
Brancolini, C., Lazarevic, D., Rodriguez, J. & Schneider, C. Dismantling cell–cell contacts during apoptosis is coupled to a caspase-dependent proteolytic cleavage of β-catenin. J. Cell Biol. 139, 759–771 (1997).
Bannerman, D. D., Sathyamoorthy, M. & Goldblum, S. E. Bacterial lipopolysaccharide disrupts endothelial monolayer integrity and survival signalling events through caspase cleavage of adherens junction proteins. J. Biol. Chem. 273, 35371–35380 (1998).
Steinhusen, U. et al. Cleavage and shedding of E-cadherin after induction of apoptosis. J. Biol. Chem. 276, 4872–4980 (2001).
Dusek, R. L. et al. The differentiation-dependent desmosomal cadherin desmoglein 1 is a novel caspase-3 target that regulates apoptosis in keratinocytes. J. Biol. Chem. 281, 3614–3624 (2006).
Weiske, J. et al. The fate of desmosomal proteins in apoptotic cells. J. Biol. Chem. 276, 41175–41181 (2001).
Houge, G., Doskeland, S. O., Boe, R. & Lanotte, M. Selective cleavage of 28S rRNA variable regions V3 and V13 in myeloid leukaemia cell apoptosis. FEBS Lett. 315, 16–20 (1993).
Wyllie, A. H. Glucocorticoid-induced thymocyte apoptosis is associated with endogenous endonuclease activation. Nature 284, 555–556 (1980). The first study to explicitly link internucleosomal DNA fragmentation to apoptosis.
Schulze-Osthoff, K., Walczak, H., Droge, W. & Krammer, P. H. Cell nucleus and DNA fragmentation are not required for apoptosis. J. Cell Biol. 127, 15–20 (1994).
Wu, Y. C., Stanfield, G. M. & Horvitz, H. R. NUC-1, a Caenorhabditis elegans DNase II homolog, functions in an intermediate step of DNA degradation during apoptosis. Genes Dev. 14, 536–548 (2000).
Kawane, K. et al. Impaired thymic development in mouse embryos deficient in apoptotic DNA degradation. Nature Immunol. 4, 138–144 (2003).
Napirei, M. et al. T. Features of systemic lupus erythematosus in DNase1-deficient mice. Nature Genet. 25, 177–181 (2000). References 54 and 55 suggest that DNase-mediated degradation of genomic DNA from apoptotic cells is essential to ward off inappropriate activation of the immune system, which can lead to autoimmunity.
Samejima, K., Tone, S. & Earnshaw, W. C. CAD/DFF40 nuclease is dispensable for high molecular weight DNA cleavage and stage I chromatin condensation in apoptosis. J. Biol. Chem. 276, 45427–45432 (2001).
Liu, X., Zou, H., Slaughter, C. & Wang, X. DFF, a heterodimeric protein that functions downstream of caspase-3 to trigger DNA fragmentation during apoptosis. Cell 89, 175–184 (1997).
Enari, M. et al. A caspase-activated DNase that degrades DNA during apoptosis, and its inhibitor ICAD. Nature 391, 43–50 (1998). This study, along with reference 57, reported the identification of the nuclease (called CAD or DFF40) that is responsible for internucleosomal DNA fragmentation during apoptosis.
Sakahira, H., Enari, M. & Nagata, S. Cleavage of CAD inhibitor in CAD activation and DNA degradation during apoptosis. Nature 391, 96–99 (1998).
Samejima, K. & Earnshaw, W. C. Trashing the genome: the role of nucleases during apoptosis. Nature Rev. Mol. Cell Biol. 6, 677–688 (2005).
Cheung, W. L. et al. Apoptotic phosphorylation of histone H2B is mediated by mammalian sterile twenty kinase. Cell 113, 507–517 (2003).
Ura, S., Masuyama, N., Graves, J. D. & Gotoh, Y. Caspase cleavage of MST1 promotes nuclear translocation and chromatin condensation. Proc. Natl Acad. Sci. USA 98, 10148–10153 (2001). This study, together with reference 61, links proteolysis of MST1 with apoptotic chromatin condensation.
Ahn, S. H. et al.Sterile 20 kinase phosphorylates histone H2B at serine 10 during hydrogen peroxide-induced apoptosis in S. cerevisiae. Cell 120, 25–36 (2005).
Mancini, M. et al. Caspase-2 is localized at the Golgi complex and cleaves golgin-160 during apoptosis. J. Cell Biol. 149, 603–612 (2000).
Chiu, R., Novikov, L., Mukherjee, S. & Shields, D. A caspase cleavage fragment of p115 induces fragmentation of the Golgi apparatus and apoptosis. J. Cell Biol. 159, 637–648 (2002).
Lowe, M., Lane, J. D., Woodman, P. G. & Allan, V. J. Caspase-mediated cleavage of syntaxin 5 and giantin accompanies inhibition of secretory traffic during apoptosis. J. Cell Sci. 117, 1139–1150 (2003).
Lane, J. D., Allan, V. J. & Woodman, P. G. Active relocation of chromatin and endoplasmic reticulum into blebs in late apoptotic cells. J. Cell Sci. 118, 4059–4071 (2005).
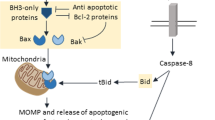
Letai, A. et al. Distinct BH3 domains either sensitize or activate mitochondrial apoptosis, serving as prototype cancer therapeutics. Cancer Cell 2, 183–192 (2002). Provided the first evidence that BH3-only proteins can act either as sensitizers or as direct activators of the BAX–BAK channel.
Kuwana, T. et al. BH3 domains of BH3-only proteins differentially regulate Bax-mediated mitochondrial membrane permeabilization both directly and indirectly. Mol. Cell 17, 525–535 (2005).
Karbowski, M. et al. Spatial and temporal association of Bax with mitochondrial fission sites, Drp1, and Mfn2 during apoptosis. J. Cell Biol. 159, 931–938 (2002).
Delivani, P., Taylor, R. C., Adrain, C., Duriez, P. & Martin, S. J. Role for CED-9 and Egl-1 as regulators of mitochondrial fission and fusion dynamics. Mol. Cell 21, 761–773 (2006).
Arnoult, D., Grodet, A., Lee, Y. J., Estaquier, J. & Blackstone, C. Release of OPA1 during apoptosis participates in the rapid and complete release of cytochrome c and subsequent mitochondrial fragmentation. J. Biol. Chem. 280, 35742–35750 (2005).
Karbowski, M., Norris, K. L., Cleland, M. M., Jeong, S.-Y. & Youle, R. J. Role of Bax and Bak in mitochondrial morphogenesis. Nature 443, 658–662 (2006). This provocative study, along with reference 71, suggests that certain BCL-2 family proteins may have roles in mitochondrial fission and fusion dynamics that are unrelated to their roles within the cell death machinery.
Ricci. J. E. et al. Disruption of mitochondrial function during apoptosis is mediated by caspase cleavage of the p75 subunit of complex I of the electron transport chain. Cell 117, 773–786 (2004).
Sun, X. M. et al. Caspase activation inhibits proteasome function during apoptosis. Mol. Cell 14, 81–93 (2004).
Adrain, C., Creagh, E. M., Cullen, S. P. & Martin, S. J. Caspase-dependent inactivation of proteasome function during programmed cell death in Drosophila and man. J. Biol. Chem. 279, 36923–36930 (2004).
Voll, R. E, Hermann, M., Roth, E. A., Stach, C. & Kalden, J. R. Immunosupressive effects of apoptotic cells. Nature 390, 350–351 (1997).
Fadok, V. et al. Macrophages that have ingested apoptotic cells in vitro inhibit proinflammatory cytokine production through autocrine/paracrine mechanisms involving TGF-β, PGE2, and PAF. J. Clin. Invest. 101, 890–898 (1998).
Bellone, M. et al. Processing of engulfed apoptotic bodies yields T-cell epitopes. J. Immunol. 159, 5391–5399 (1997).
Albert, M. I., Sauter, B. & Bhardwaj, N. Dendritic cells acquire antigen from apoptotic cells and induce class-restricted CTLs. Nature 392, 86–89 (1998). This study presents the first evidence that peptides from apoptotic cells may be cross-presented to class-I-restricted cytotoxic T lymphocytes.
Steinman, R. M., Turley, S., Mellman, I. & Inaba, K. The induction of tolerance by dendritic cells that have captured apoptotic cells. J. Exp. Med. 191, 411–416 (2000).
Orlando, K. A., Stone, N. L. & Pittman, R. N. Rho kinase regulates fragmentation and phagocytosis of apoptotic cells. Exp. Cell Res. 312, 5–15 (2006).
Lauber, K. et al. Apoptotic cells induce migration of phagocytes via caspase-3-mediated release of a lipid attraction signal. Cell 113, 717–730 (2003). This intriguing study suggested that apoptotic cells can secrete molecules with chemotactic properties.
Horino, K. et al. A monocyte chemotactic factor, S19 ribosomal protein dimer, in phagocytic clearance of apoptotic cells. Lab. Invest. 78, 603–617 (1998).
Wakasugi, K. & Schimmel, P. Two distinct cytokines released from a human aminoacyl-tRNA synthetase. Science 284, 147–151 (1999).
Fadok, V. A. et al. Exposure of phosphatidylserine on the surface of apoptotic lymphocytes triggers specific recognition and removal by macrophages. J. Immunol. 148, 2207–2216 (1992). This was the first report to suggest that phosphatidylserine might be exposed on the surface of apoptotic cells.
Martin, S. J. et al. Early redistribution of plasma membrane phosphatidylserine is a general feature of apoptosis regardless of the initiating stimulus: inhibition by overexpression of Bcl-2 and Abl. J. Exp. Med. 182, 1545–1556 (1995). This report directly demonstrates that phosphatidylserine exposure is a universal event during apoptosis of various cell types. It also introduced the annexin-5 labelling method.
Martin, S. J., Finucane, D. M., Amarante-Mendes, G. P., O'Brien, G. A. & Green, D. R. Phosphatidylserine externalization during CD95-induced apoptosis of cells and cytoplasts requires ICE/CED-3 protease activity. J. Biol. Chem. 271, 28753–28756 (1996).
Hamon, Y. et al. ABC1 promotes engulfment of apoptotic cells and transbilayer redistribution of phosphatidlyserine. Nature Cell Biol. 2, 399–406 (2000).
Fadok, V. A. et al. A receptor for phosphatidylserine-specific clearance of apoptotic cells. Nature 405, 85–90 (2000).
Mitchell, J. E. et al. The presumptive phosphatidylserine receptor is dispensable for innate anti-inflammatory recognition and clearance of apoptotic cells. J. Biol. Chem. 281, 5718–5725 (2006).
Hanayama, R. et al. Identification of a factor that links apoptotic cells to phagocytes. Nature 417, 182–187 (2002).
Manodori, A. B., Barabino, G. A., Lubin, B. H. & Kuypers, F. A. Adherence of phosphatidylserine-exposing erythrocytes to endothelial matrix thrombospondin. Blood 95, 1293–1300 (2000).
Greenberg, M. E. et al. Oxidized phosphatidylserine-CD36 interactions play an essential role in macrophage-dependent phagocytosis of apoptotic cells. J. Exp. Med. 203, 2613–2625 (2006).
Chang, M.-K. et al. Monoclonal antibodies against oxidised low-density lipoprotein bind to apoptotic cells and inhibit their phagocytosis by elicited macrophages: evidence that oxidation-specific epitopes mediate macrophage recognition. Proc. Natl Acad. Sci. USA 96, 6353–6358 (1999).
Oka, K. et al. Lectin-like oxidised low-density lipoprotein receptor 1 mediates phagocytosis of aged/apoptotic cells in endothelial cells. Proc. Natl Acad. Sci. USA 95, 9535–9540 (1998).
Platt, N., Suzuki, H., Kurihara, Y., Kodama, T. & Gordon, S. Role for the class A macrophage scavenger receptor in the phagocytosis of apoptotic thymocytes in vitro. Proc. Natl Acad. Sci. USA 93, 12456–12460 (1996).
Gardai, S. J. et al. Cell-surface calreticulin initiates clearance of viable or apoptotic cells through trans-activation of LRP on the phagocyte. Cell 123, 321–334 (2005).
Moffatt, O. D., Devitt, A., Bell, E. D., Simmons, D. L. & Gregory, C. D. Macrophage recognition of ICAM-3 on apoptotic leukocytes. J. Immunol. 162, 6800–6810 (1999).
Arur, S. et al. Annexin I is an endogenous ligand that mediates apoptotic cell engulfment. Dev. Cell 4, 587–598 (2003).
Hall, S. E., Savill, J. S., Henson, P. M. & Haslett, C. Apoptotic neutrophils are phagocytosed by fibroblasts with participation of the fibroblast vitronectin receptor and involvement of a mannose/fucose-specific lectin. J. Immunol. 153, 3218–3227 (1994).
Ogden, C. A. et al. C1q and mannose-binding lectin engagement of cell surface calreticulin and CD91 initiates macropinocytosis and uptake of apoptotic cells. J. Exp. Med. 194, 781–795 (2001).
Savill, J., Hogg, N., Ren, Y. & Haslett, C. Thrombospondin cooperates with CD36 and the vitronectin receptor in macrophage recognition of neutrophils undergoing apoptosis. J. Clin. Invest. 90, 1513–1522 (1992).
Takizawa, F., Tsuji, S. & Nagawa, S. Enhancement of macrophage phagocytosis upon iC3b deposition on apoptotic cells. FEBS Lett. 397, 269–272 (1996).
Acknowledgements
Work in the Martin laboratory is supported by an award from Science Foundation Ireland. We thank P. Delivani for providing the images for Figure 1. We apologize to colleagues for citing reviews instead of primary papers in several places owing to space constraints.
Author information
Authors and Affiliations
Corresponding author
Supplementary information
Supplementary information S1 (movie) | Time-lapse movie of human HeLa cells undergoing apoptosis.
HeLa cells were induced to undergo apoptosis by exposure to 10 μM daunorubicin. Images were taken every minute over ∼12 hours and were animated at 10 frames/second. The first image represents healthy cells 2 h after treatment. By 4 h, one of the cells begins to round up and detach from the substratum, and this is followed 30 min later by retraction of the neighbouring cell. Dynamic plasma membrane blebbing is then evident in both cells and this continues for several hours. Later, membrane blebs become larger and more stable until, ∼8.5 hours after the cells were exposed to the pro-apoptotic drug, secondary necrosis begins. During secondary necrosis, cells cease to bleb and large balloon-like swellings can be seen as cells lose plasma membrane integrity and release their contents into the surroundings. Secondary necrosis is a highly undesirable endpoint and this is normally prevented in vivo through removal of apoptotic cells by phagocytes early in the process. (MOV 3995 kb)
Related links
Related links
DATABASES
OMIM
FURTHER INFORMATION
Glossary
- Phagocyte
-
A cell that can engulf and ingest foreign material, such as an apoptotic cell corpse, which then undergoes digestion within lysosomes.
- Caspase
-
One of a family of proteases that have an essential Cys residue in their active site and a requirement for an Asp residue in the substrate cleavage site. Initiator caspases are typically activated in response to particular stimuli, whereas effector caspases are particularly important for the ordered dismantling of vital cellular structures.
- Apoptotic body
-
During apoptosis, cells collapse into small intact fragments that exclude vital dyes. Such fragments are termed apoptotic bodies.
- Apoptosome
-
A large protein complex that comprises cytochrome c and apoptotic protease-activating factor-1 (APAF1), and forms in the presence of ATP or dATP. The apoptosome recruits pro-caspase-9 and results in the allosteric activation of caspase-9.
- Granzyme B
-
A Ser protease contained within the secretory granules of cytotoxic lymphocytes and natural killer cells. Granzyme B cleaves its protein substrates after Asp residues, and can promote caspase activation and apoptosis.
- Myosin
-
One of a family of actin-associated motor proteins that bind to actin fibres by their head domain and use ATP hydrolysis to move along fibres. The tail domain of myosin associates with cargo or other myosin molecules.
- Nuclear lamin
-
One of a family of intermediate filament proteins that form the proteinaceous nuclear lamina structure, which surrounds the nucleus.
- ROCK1
-
An effector of the small GTPase Rho, ROCK1 is a kinase that can phosphorylate myosin light chain, which alters the dynamics of the actin cytoskeleton.
- 14-3-3 protein
-
One of a family of small phosphopeptide-binding dimeric proteins that typically act as adaptors or localize components of signalling pathways.
- BH3-only protein
-
A member of a class of pro-apoptotic proteins in the larger B-cell lymphoma-2 (BCL-2) family of proteins. BH3-only proteins share a short (∼12 amino acid) motif known as the BH3 domain that exhibits homology with a region within BCL-2.
- Phosphatidylserine
-
A membrane phospholipid that is usually localized to the inner leaflet of the plasma membrane, but which is translocated to the outer leaflet during apoptosis.
- Scavenger receptor
-
A type of cell-surface receptor that recognizes modified low-density lipoprotein or other negatively charged macromolecules, and often functions to remove foreign substances or waste materials from the body.
Rights and permissions
About this article
Cite this article
Taylor, R., Cullen, S. & Martin, S. Apoptosis: controlled demolition at the cellular level. Nat Rev Mol Cell Biol 9, 231–241 (2008). https://doi.org/10.1038/nrm2312
Issue Date:
DOI: https://doi.org/10.1038/nrm2312
This article is cited by
-
Exosomes derived from programmed cell death: mechanism and biological significance
Cell Communication and Signaling (2024)
-
Mutation and apoptosis are well-coordinated for protecting against DNA damage-inducing toxicity in Drosophila
Genes and Environment (2023)
-
Elucidating the molecular mechanisms underlying the induction of autophagy by antidepressant-like substances in C57BL/6J mouse testis model upon LPS challenge
Cell Communication and Signaling (2023)
-
CD33 as a leukocyte-associated marker expressed on human spermatozoa
BMC Research Notes (2023)
-
Optimized allotopic expression of mitochondrial ND6 transgene restored complex I and apoptosis deficiencies caused by LHON-linked ND6 14484T > C mutation
Journal of Biomedical Science (2023)
