
Key Points
-
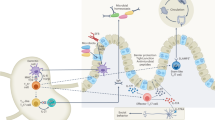
Study of the interleukin-23 (IL-23)–IL-17 pathway has greatly changed our understanding of autoimmune inflammatory mechanisms. This Review highlights the many known and newly identified innate IL-17-producing cell populations found in the skin, gut, lungs and associated lymphoid organs.
-
We describe the cytokine and transcriptional regulation of innate IL-17-producing cells, with an emphasis on the crucial roles of IL-23 receptor signalling and retinoic acid receptor-related orphan receptor-γt (RORγt) in their development and/or function.
-
Innate IL-17 and IL-22 have physiological roles in immune surveillance, which include maintenance of mucosal barrier integrity, induction of antimicrobial protein production by epithelial cells and recruitment of inflammatory myeloid cells to the site of injury.
-
Current evidence suggests that early IL-17 and IL-22 secretion by innate cells shapes the balance of mucosal homeostasis and inflammation; we discuss the potential for autoimmunity if this system is dysregulated.
-
We provide insight into the future direction of research in this area and describe the potential implications for the development of new therapeutic agents.
Abstract
The cytokine interleukin-17 (IL-17) has received considerable attention since the discovery of a distinct CD4+ T helper (TH) cell subset that produces it, known as the TH17 cell subset. Despite the fact that most of the recent literature describes IL-17 as a T cell-secreted cytokine, much of the IL-17 released during an inflammatory response is produced by innate immune cells. In this Review, we explore the many innate immune cell populations that are an early source of IL-17 in response to stress, injury or pathogens. These early sources have been shown to have a central role in the initiation of IL-17-dependent immune responses, even before the first CD4+T cell sees its cognate antigen and initiates the TH17 cell developmental programme.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout



Similar content being viewed by others


Change history
25 June 2010
In the version of the article initially published, the table in BOX 1 incorrectly listed IL-17RB as the receptor for IL-17E (also known as IL-25). The correct receptor should have been IL-17RA-IL-17RB.
References
Rouvier, E., Luciani, M. F., Mattei, M. G., Denizot, F. & Golstein, P. CTLA-8, cloned from an activated T cell, bearing AU-rich messenger RNA instability sequences, and homologous to a herpesvirus saimiri gene. J. Immunol. 150, 5445–5456 (1993).
Weaver, C. T., Hatton, R. D., Mangan, P. R. & Harrington, L. E. IL-17 family cytokines and the expanding diversity of effector T cell lineages. Annu. Rev. Immunol. 25, 821–852 (2007).
Langrish, C. L. et al. IL-23 drives a pathogenic T cell population that induces autoimmune inflammation. J. Exp. Med. 201, 233–240 (2005).
Harrington, L. E. et al. Interleukin 17-producing CD4+ effector T cells develop via a lineage distinct from the T helper type 1 and 2 lineages. Nature Immunol. 6, 1123–1132 (2005).
Park, H. et al. A distinct lineage of CD4 T cells regulates tissue inflammation by producing interleukin 17. Nature Immunol. 6, 1133–1141 (2005).
Veldhoen, M., Hocking, R. J., Atkins, C. J., Locksley, R. M. & Stockinger, B. TGFβ in the context of an inflammatory cytokine milieu supports de novo differentiation of IL-17-producing T cells. Immunity 24, 179–189 (2006).
Wei, L., Laurence, A., Elias, K. M. & O'Shea, J. J. IL-21 is produced by Th17 cells and drives IL-17 production in a STAT3-dependent manner. J. Biol. Chem. 282, 34605–34610 (2007).
Zheng, Y. et al. Interleukin-22 mediates early host defense against attaching and effacing bacterial pathogens. Nature Med. 14, 282–289 (2008). This is the first study to suggest that RORγt+ lymphocytes can produce IL-22, which promotes epithelial cell production of REG3γ required for host defence against extracellular gut pathogens.
Korn, T. et al. IL-21 initiates an alternative pathway to induce proinflammatory TH17 cells. Nature 448, 484–487 (2007).
Ivanov, I. I. et al. The orphan nuclear receptor RORγt directs the differentiation program of proinflammatory IL-17+ T helper cells. Cell 126, 1121–1133 (2006). The data in this report show RORγt is the key transcription factor promoting IL-17A, IL-17F, and IL-22 expression by T H 17 cells. Subsequent studies showed that RORγt also regulates innate lymphocytes.
Happel, K. I. et al. Cutting edge: roles of Toll-like receptor 4 and IL-23 in IL-17 expression in response to Klebsiella pneumoniae infection. J. Immunol. 170, 4432–4436 (2003).
Happel, K. I. et al. Divergent roles of IL-23 and IL-12 in host defense against Klebsiella pneumoniae. J. Exp. Med. 202, 761–769 (2005).
Ferretti, S., Bonneau, O., Dubois, G. R., Jones, C. E. & Trifilieff, A. IL-17, produced by lymphocytes and neutrophils, is necessary for lipopolysaccharide-induced airway neutrophilia: IL-15 as a possible trigger. J. Immunol. 170, 2106–2112 (2003).
Fossiez, F. et al. T cell interleukin-17 induces stromal cells to produce proinflammatory and hematopoietic cytokines. J. Exp. Med. 183, 2593–2603 (1996).
Kennedy, J. et al. Mouse IL-17: a cytokine preferentially expressed by αβ TCR + CD4−CD8− T cells. J. Interferon Cytokine Res. 16, 611–617 (1996).
Lochner, M. et al. In vivo equilibrium of proinflammatory IL-17+ and regulatory IL-10+ Foxp3+ RORγt+ T cells. J. Exp. Med. 205, 1381–1393 (2008). This study used RORγteGFP reporter mice to determine the proportion of 'type 17' [Au: T H 17 type?] αβ and γδ T cell subsets in skin, lungs and gut. It shows the expansion of IL-17+RORγt+ cells in situ during inflammatory responses.
Sutton, C. E. et al. Interleukin-1 and IL-23 induce innate IL-17 production from γδ T cells, amplifying Th17 responses and autoimmunity. Immunity 31, 331–341 (2009). This report shows that γδ T cells have an early role in promoting CNS inflammation. The authors suggest that innate cell-produced IL-17 directly enhances development of MOG-specific T H 17 cells.
Veldhoen, M. et al. The aryl hydrocarbon receptor links TH17-cell-mediated autoimmunity to environmental toxins. Nature 453, 106–109 (2008). The data presented here suggest that AHR is a transcriptional regulator associated with regulatory T cells and IL-17-producing cells. Subsequent studies showed that AHR promotes IL-22 production in both adaptive and innate cell populations.
Korn, T., Bettelli, E., Oukka, M. & Kuchroo, V. K. IL-17 and Th17 Cells. Annu. Rev. Immunol. 27, 485–517 (2009).
McGeachy, M. J. & Cua, D. J. Th17 cell differentiation: the long and winding road. Immunity 28, 445–453 (2008).
Aggarwal, S., Ghilardi, N., Xie, M. H., de Sauvage, F. J. & Gurney, A. L. Interleukin-23 promotes a distinct CD4 T cell activation state characterized by the production of interleukin-17. J. Biol. Chem. 278, 1910–1914 (2003).
Murphy, C. A. et al. Divergent pro- and antiinflammatory roles for IL-23 and IL-12 in joint autoimmune inflammation. J. Exp. Med. 198, 1951–1957 (2003). This early study shows that IL-23-deficient mice lack IL-17-producing cells following antigenic challenge, which suggests a role for IL-17+ T H cells in promoting autoimmune inflammation.
Uhlig, H. H. et al. Differential activity of IL-12 and IL-23 in mucosal and systemic innate immune pathology. Immunity 25, 309–318 (2006). This study shows that IL-23 can activate innate IL-17-producing cells in RAG-deficient mice. IL-23 acts locally in gut mucosal tissues whereas IL-12 has a dominant role in promoting systemic inflammation.
Cua, D. J. et al. Interleukin-23 rather than interleukin-12 is the critical cytokine for autoimmune inflammation of the brain. Nature 421, 744–748 (2003). IL-23 directly promotes innate cell production of inflammatory cytokines. This is the first suggestion that IL-23 acts on both innate and adaptive populations.
Aujla, S. J. et al. IL-22 mediates mucosal host defense against Gram-negative bacterial pneumonia. Nature Med. 14, 275–281 (2008).
Cella, M. et al. A human natural killer cell subset provides an innate source of IL-22 for mucosal immunity. Nature 457, 722–725 (2009). This study shows NK-like cells that produce IL-22 are essential for host defence against gut bacterial infection. In this mouse system, the NKp46+ IL-22-producing cells do not seem to produce IL-17.
Crellin, N. K., Trifari, S., Kaplan, C. D., Cupedo, T. & Spits, H. Human NKp44+IL-22+ cells and LTi-like cells constitute a stable RORC+ lineage distinct from conventional natural killer cells. J. Exp. Med. 207, 281–290 (2010). This study shows that human RORγt+ NK-like cells can be isolated from peripheral blood mononuclear cells. These cells produce both IL-17 and IL-22 and are probably the counterparts of mouse NKp46+ IL-22-producing cells. A subsequent study from the same group showed that NKp46+ cells may develop from LTi-like cells in the human system.
Takatori, H. et al. Lymphoid tissue inducer-like cells are an innate source of IL-17 and IL-22. J. Exp. Med. 206, 35–41 (2009). The is the first study to show that KIT+LIN− LTi-like cells are an innate source of IL-17 and IL-22 during an immune response to fungal antigens.
Roark, C. L. et al. Exacerbation of collagen-induced arthritis by oligoclonal, IL-17-producing γδ T cells. J. Immunol. 179, 5576–5583 (2007).
Ito, Y. et al. γδ T cells are the predominant source of interleukin-17 in affected joints in collagen-induced arthritis, but not in rheumatoid arthritis. Arthritis Rheum. 60, 2294–2303 (2009).
Cui, Y. et al. Major role of γδ T cells in the generation of IL-17+ uveitogenic T cells. J. Immunol. 183, 560–567 (2009).
Fenoglio, D. et al. Vδ1 T lymphocytes producing IFN-γ and IL-17 are expanded in HIV-1-infected patients and respond to Candida albicans. Blood 113, 6611–6618 (2009).
Aujla, S. J., Dubin, P. J. & Kolls, J. K. Th17 cells and mucosal host defense. Semin. Immunol. 19, 377–382 (2007).
Lockhart, E., Green, A. M. & Flynn, J. L. IL-17 production is dominated by γδ T cells rather than CD4 T cells during Mycobacterium tuberculosis infection. J. Immunol. 177, 4662–4669 (2006).
Hayday, A. C. γδ T cells and the lymphoid stress-surveillance response. Immunity 31, 184–196 (2009).
Michel, M. L. et al. Identification of an IL-17-producing NK1.1neg iNKT cell population involved in airway neutrophilia. J. Exp. Med. 204, 995–1001 (2007).
Passos, S. T. et al. IL-6 promotes NK cell production of IL-17 during toxoplasmosis. J. Immunol. 184, 1776–1783 (2010).
Jensen, K. D. et al. Thymic selection determines γδ T cell effector fate: antigen-naive cells make interleukin-17 and antigen-experienced cells make interferon γ. Immunity 29, 90–100 (2008).
Ribot, J. C. et al. CD27 is a thymic determinant of the balance between interferon-γ- and interleukin 17-producing γδ T cell subsets. Nature Immunol. 10, 427–436 (2009).
Haas, J. D. et al. CCR6 and NK1.1 distinguish between IL-17A and IFN-γ-producing γδ effector T cells. Eur. J. Immunol. 39, 3488–3497 (2009).
Shibata, K. et al. Identification of CD25+ γδ T cells as fetal thymus-derived naturally occurring IL-17 producers. J. Immunol. 181, 5940–5947 (2008).
Laurence, A. et al. Interleukin-2 signaling via STAT5 constrains T helper 17 cell generation. Immunity 26, 371–381 (2007).
Kisielow, J., Kopf, M. & Karjalainen, K. SCART scavenger receptors identify a novel subset of adult γδ T cells. J. Immunol. 181, 1710–1716 (2008).
Sun, Z. et al. Requirement for RORγ in thymocyte survival and lymphoid organ development. Science 288, 2369–2373 (2000).
Eberl, G. et al. An essential function for the nuclear receptor RORγt in the generation of fetal lymphoid tissue inducer cells. Nature Immunol. 5, 64–73 (2004).
Fukuyama, S. et al. Initiation of NALT organogenesis is independent of the IL-7R, LTβR, and NIK signaling pathways but requires the Id2 gene and CD3−CD4+CD45+ cells. Immunity 17, 31–40 (2002).
Adachi, S. et al. Essential role of IL-7 receptor α in the formation of Peyer's patch anlage. Int. Immunol. 10, 1–6 (1998).
Yoshida, H. et al. IL-7 receptor alpha+ CD3− cells in the embryonic intestine induces the organizing center of Peyer's patches. Int. Immunol. 11, 643–655 (1999).
Meier, D. et al. Ectopic lymphoid-organ development occurs through interleukin 7-mediated enhanced survival of lymphoid-tissue-inducer cells. Immunity 26, 643–654 (2007).
Cupedo, T., Kraal, G. & Mebius, R. E. The role of CD45+CD4+CD3− cells in lymphoid organ development. Immunol. Rev. 189, 41–50 (2002).
Cupedo, T. & Mebius, R. E. Cellular interactions in lymph node development. J. Immunol. 174, 21–25 (2005).
Finke, D., Acha-Orbea, H., Mattis, A., Lipp, M. & Kraehenbuhl, J. CD4+CD3− cells induce Peyer's patch development: role of α4β1 integrin activation by CXCR5. Immunity 17, 363–373 (2002).
Mebius, R. E. Organogenesis of lymphoid tissues. Nature Rev. Immunol. 3, 292–303 (2003).
Cupedo, T. et al. Human fetal lymphoid tissue-inducer cells are interleukin 17-producing precursors to RORC+ CD127+ natural killer-like cells. Nature Immunol. 10, 66–74 (2009). Human NK-like cells produce both IL-17 and IL-22 and may share a common lineage with LTi-like cells. Their ability to produce both IL-17 and IL-22 is in contrast to mouse NKp46+ cells, which only produce IL-22.
Lane, P. et al. Lymphoid tissue inducer cells in adaptive CD4 T cell dependent responses. Semin. Immunol. 20, 159–163 (2008).
Mebius, R. E., Rennert, P. & Weissman, I. L. Developing lymph nodes collect CD4+CD3− LTβ+ cells that can differentiate to APC NK cells, and follicular cells but not T or B cells. Immunity 7, 493–504 (1997).
Kronenberg, M. & Engel, I. On the road: progress in finding the unique pathway of invariant NKT cell differentiation. Curr. Opin. Immunol. 19, 186–193 (2007).
Michel, M. L. et al. Critical role of ROR-γt in a new thymic pathway leading to IL-17-producing invariant NKT cell differentiation. Proc. Natl Acad. Sci. USA 105, 19845–19850 (2008).
Campillo-Gimenez, L. et al. AIDS progression is associated with the emergence of IL-17-producing cells early after simian immunodeficiency virus infection. J. Immunol. 184, 984–992 (2010).
Grajewski, R. S. et al. Activation of invariant NKT cells ameliorates experimental ocular autoimmunity by a mechanism involving innate IFN-γ production and dampening of the adaptive Th1 and Th17 responses. J. Immunol. 181, 4791–4797 (2008).
Luci, C. et al. Influence of the transcription factor RORγt on the development of NKp46+ cell populations in gut and skin. Nature Immunol. 10, 75–82 (2009). One of the first reports suggesting NKp46+ cells are the early innate IL-22-producing cells that have a role in skin and mucosal tissue repair and homeostasis.
Satoh-Takayama, N. et al. IL-7 and IL-15 independently program the differentiation of intestinal CD3−NKp46+ cell subsets from Id2-dependent precursors. J. Exp. Med. 207, 273–280 (2010).
Dhiman, R. et al. IL-22 produced by human NK cells inhibits growth of Mycobacterium tuberculosis by enhancing phagolysosomal fusion. J. Immunol. 183, 6639–6645 (2009).
Sanos, S. L. & Diefenbach, A. Isolation of NK cells and NK-like cells from the intestinal lamina propria. Methods Mol. Biol. 612, 505–517 (2010).
Satoh-Takayama, N. et al. Microbial flora drives interleukin 22 production in intestinal NKp46+ cells that provide innate mucosal immune defense. Immunity 29, 958–970 (2008).
Satoh-Takayama, N. et al. The natural cytotoxicity receptor NKp46 is dispensable for IL-22-mediated innate intestinal immune defense against Citrobacter rodentium. J. Immunol. 183, 6579–6587 (2009).
Ayabe, T. et al. Secretion of microbicidal α-defensins by intestinal Paneth cells in response to bacteria. Nature Immunol. 1, 113–118 (2000).
Wilson, C. L. et al. Regulation of intestinal α-defensin activation by the metalloproteinase matrilysin in innate host defense. Science 286, 113–117 (1999).
Bultinck, J., Brouckaert, P. & Cauwels, A. The in vivo contribution of hematopoietic cells to systemic TNF and IL-6 production during endotoxemia. Cytokine 36, 160–166 (2006).
Keshav, S. et al. Tumor necrosis factor mRNA localized to Paneth cells of normal murine intestinal epithelium by in situ hybridization. J. Exp. Med. 171, 327–332 (1990).
Lala, S. et al. Crohn's disease and the NOD2 gene: a role for paneth cells. Gastroenterology 125, 47–57 (2003).
Rumio, C. et al. Degranulation of paneth cells via toll-like receptor 9. Am. J. Pathol. 165, 373–381 (2004).
Salzman, N. H. et al. Enteric defensins are essential regulators of intestinal microbial ecology. Nature Immunol. 11, 76–83 (2010).
Takahashi, N. et al. IL-17 produced by Paneth cells drives TNF-induced shock. J. Exp. Med. 205, 1755–1761 (2008).
Li, L. et al. IL-17 produced by neutrophils regulates IFN-γ-mediated neutrophil migration in mouse kidney ischemia-reperfusion injury. J. Clin. Invest. 120, 331–342 (2010).
Hoshino, A. et al. MPO-ANCA induces IL-17 production by activated neutrophils in vitro via classical complement pathway-dependent manner. J. Autoimmun. 31, 79–89 (2008).
Ivanov, I. I. et al. Induction of intestinal Th17 cells by segmented filamentous bacteria. Cell 139, 485–498 (2009). Comparative analysis of microbiota from T H 17 cell-deficient and T H 17 cell-sufficient mice identified SFB as inducers of T H 17 cells in the gut. This study suggests that commensal bacteria can shape the balance of immune cell subsets.
Ivanov, I. I. & Littman, D. R. Segmented filamentous bacteria take the stage. Mucosal Immunol. 3, 209–212 (2010).
Buonocore, S. et al. Innate lymphoid cells drive interleukin-23-dependent innate intestinal pathology. Nature 464, 1371–1375 (2010). This study identified THY1hiSCA1+ innate lymphoid cells that produce IL-17 and IL-22. In contrast to LTi-like and NKp46+ cells, this newly described subset also produces IFNγ and is regulated by both RORγt and T-bet.
Gaffen, S. L. Structure and signalling in the IL-17 receptor family. Nature Rev. Immunol. 9, 556–567 (2009).
Tanaka, S. et al. Natural occurring IL-17 producing T cells regulate the initial phase of neutrophil mediated airway responses. J. Immunol. 183, 7523–7530 (2009).
Rachitskaya, A. V. et al. Cutting edge: NKT cells constitutively express IL-23 receptor and RORγt and rapidly produce IL-17 upon receptor ligation in an IL-6-independent fashion. J. Immunol. 180, 5167–5171 (2008).
He, Y. W., Deftos, M. L., Ojala, E. W. & Bevan, M. J. RORγt, a novel isoform of an orphan receptor, negatively regulates Fas ligand expression and IL-2 production in T cells. Immunity 9, 797–806 (1998).
Doisne, J. M. et al. Skin and peripheral lymph node invariant NKT cells are mainly retinoic acid receptor-related orphan receptor γt+ and respond preferentially under inflammatory conditions. J. Immunol. 183, 2142–2149 (2009).
Esser, C., Rannug, A. & Stockinger, B. The aryl hydrocarbon receptor in immunity. Trends Immunol. 30, 447–454 (2009).
Schecter, A., Birnbaum, L., Ryan, J. J. & Constable, J. D. Dioxins: an overview. Environ. Res. 101, 419–428 (2006).
Rannug, A. et al. Certain photooxidized derivatives of tryptophan bind with very high affinity to the Ah receptor and are likely to be endogenous signal substances. J. Biol. Chem. 262, 15422–15427 (1987).
Quintana, F. J. et al. Control of Treg and TH17 cell differentiation by the aryl hydrocarbon receptor. Nature 453, 65–71 (2008).
Martin, B., Hirota, K., Cua, D. J., Stockinger, B. & Veldhoen, M. Interleukin-17-producing γδ T cells selectively expand in response to pathogen products and environmental signals. Immunity 31, 321–330 (2009).
Zhou, L. & Littman, D. R. Transcriptional regulatory networks in Th17 cell differentiation. Curr. Opin. Immunol. 21, 146–152 (2009).
Kimura, A., Naka, T., Nohara, K., Fujii-Kuriyama, Y. & Kishimoto, T. Aryl hydrocarbon receptor regulates Stat1 activation and participates in the development of Th17 cells. Proc. Natl Acad. Sci. USA 105, 9721–9726 (2008).
Duan, J., Chung, H., Troy, E. & Kasper, D. L. Microbial colonization drives expansion of IL-1 receptor 1-expressing and IL-17-producing γ/δ T cells. Cell Host Microbe 7, 140–150 (2010).
Yoshiga, Y. et al. Invariant NKT cells produce IL-17 through IL-23-dependent and -independent pathways with potential modulation of Th17 response in collagen-induced arthritis. Int. J. Mol. Med. 22, 369–374 (2008).
Riol-Blanco, L. et al. IL-23 receptor regulates unconventional IL-17-producing T cells that control bacterial infections. J. Immunol. 184, 1710–1720 (2010).
Curtis, M. M. & Way, S. S. Interleukin-17 in host defence against bacterial, mycobacterial and fungal pathogens. Immunology 126, 177–185 (2009).
Iwakura, Y., Nakae, S., Saijo, S. & Ishigame, H. The roles of IL-17A in inflammatory immune responses and host defense against pathogens. Immunol. Rev. 226, 57–79 (2008).
Kolls, J. K. & Linden, A. Interleukin-17 family members and inflammation. Immunity 21, 467–476 (2004).
Ye, P. et al. Interleukin-17 and lung host defense against Klebsiella pneumoniae infection. Am. J. Respir. Cell. Mol. Biol. 25, 335–340 (2001).
Umemura, M. et al. IL-17-mediated regulation of innate and acquired immune response against pulmonary Mycobacterium bovis bacille Calmette-Guerin infection. J. Immunol. 178, 3786–3796 (2007).
Peng, M. Y. et al. Interleukin 17-producing γδ T cells increased in patients with active pulmonary tuberculosis. Cell. Mol. Immunol. 5, 203–208 (2008).
D'Souza, C. D. et al. An anti-inflammatory role for γδ T lymphocytes in acquired immunity to Mycobacterium tuberculosis. J. Immunol. 158, 1217–1221 (1997).
Khader, S. A. et al. IL-23 and IL-17 in the establishment of protective pulmonary CD4+ T cell responses after vaccination and during Mycobacterium tuberculosis challenge. Nature Immunol. 8, 369–377 (2007).
Molne, L., Corthay, A., Holmdahl, R. & Tarkowski, A. Role of γ/δ T cell receptor-expressing lymphocytes in cutaneous infection caused by Staphylococcus aureus. Clin. Exp. Immunol. 132, 209–215 (2003).
Cho, J. S. et al. IL-17 is essential for host defense against cutaneous Staphylococcus aureus infection in mice. J. Clin. Invest. 120, 1762–1773 (2010).
Shibata, K., Yamada, H., Hara, H., Kishihara, K. & Yoshikai, Y. Resident Vδ1+ γδ T cells control early infiltration of neutrophils after Escherichia coli infection via IL-17 production. J. Immunol. 178, 4466–4472 (2007).
Siegemund, S. et al. Differential IL-23 requirement for IL-22 and IL-17A production during innate immunity against Salmonella enterica serovar Enteritidis. Int. Immunol. 21, 555–565 (2009).
Meeks, K. D., Sieve, A. N., Kolls, J. K., Ghilardi, N. & Berg, R. E. IL-23 is required for protection against systemic infection with Listeria monocytogenes. J. Immunol. 183, 8026–8034 (2009).
Kelly, M. N. et al. Interleukin-17/interleukin-17 receptor-mediated signaling is important for generation of an optimal polymorphonuclear response against Toxoplasma gondii infection. Infect. Immun. 73, 617–621 (2005).
Chen, Y., Chou, K., Fuchs, E., Havran, W. L. & Boismenu, R. Protection of the intestinal mucosa by intraepithelial γδ T cells. Proc. Natl Acad. Sci. USA 99, 14338–14343 (2002).
Kuhl, A. A. et al. Aggravation of intestinal inflammation by depletion/deficiency of γδ T cells in different types of IBD animal models. J. Leukoc. Biol. 81, 168–175 (2007).
Kinugasa, T., Sakaguchi, T., Gu, X. & Reinecker, H. C. Claudins regulate the intestinal barrier in response to immune mediators. Gastroenterology 118, 1001–1011 (2000).
Ogawa, A., Andoh, A., Araki, Y., Bamba, T. & Fujiyama, Y. Neutralization of interleukin-17 aggravates dextran sulfate sodium-induced colitis in mice. Clin. Immunol. 110, 55–62 (2004).
Liang, S. C. et al. Interleukin (IL)-22 and IL-17 are coexpressed by Th17 cells and cooperatively enhance expression of antimicrobial peptides. J. Exp. Med. 203, 2271–2279 (2006).
Chan, Y. R. et al. Lipocalin 2 is required for pulmonary host defense against Klebsiella infection. J. Immunol. 182, 4947–4956 (2009).
Ismail, A. S., Behrendt, C. L. & Hooper, L. V. Reciprocal interactions between commensal bacteria and γδ intraepithelial lymphocytes during mucosal injury. J. Immunol. 182, 3047–3054 (2009).
Cash, H. L., Whitham, C. V., Behrendt, C. L. & Hooper, L. V. Symbiotic bacteria direct expression of an intestinal bactericidal lectin. Science 313, 1126–1130 (2006). REG3γ is a potent antimicrobial agent that kills Gram-positive bacteria. Subsequent studies showed that REG3γ is strongly induced by IL-22.
Brandl, K., Plitas, G., Schnabl, B., DeMatteo, R. P. & Pamer, E. G. MyD88-mediated signals induce the bactericidal lectin RegIIIγ and protect mice against intestinal Listeria monocytogenes infection. J. Exp. Med. 204, 1891–1900 (2007).
Littman, D. R. & Rudensky, A. Y. Th17 and regulatory T cells in mediating and restraining inflammation. Cell 140, 845–858 (2010).
Ely, L. K., Fischer, S. & Garcia, K. C. Structural basis of receptor sharing by interleukin 17 cytokines. Nature Immunol. 10, 1245–1251 (2009).
Ishigame, H. et al. Differential roles of interleukin-17A and -17F in host defense against mucoepithelial bacterial infection and allergic responses. Immunity 30, 108–119 (2009).
Wucherpfennig, K. W. et al. γδ T-cell receptor repertoire in acute multiple sclerosis lesions. Proc. Natl Acad. Sci. USA 89, 4588–4592 (1992).
Shimonkevitz, R., Colburn, C., Burnham, J. A., Murray, R. S. & Kotzin, B. L. Clonal expansions of activated γ/δ T cells in recent-onset multiple sclerosis. Proc. Natl Acad. Sci. USA 90, 923–927 (1993).
Kang, Z. et al. Astrocyte-restricted ablation of interleukin-17-induced Act1-mediated signaling ameliorates autoimmune encephalomyelitis. Immunity 32, 414–425 (2010).
Duerr, R. H. et al. A genome-wide association study identifies IL23R as an inflammatory bowel disease gene. Science 314, 1461–1463 (2006). First study to show that IL23R gene polymorphism is linked to human inflammatory bowel disease susceptibility. Numerous studies have since confirmed and extended this initial observation.
Capon, F. et al. Sequence variants in the genes for the interleukin-23 receptor (IL23R) and its ligand (IL12B) confer protection against psoriasis. Hum. Genet. 122, 201–206 (2007).
Rueda, B. et al. The IL23R Arg381Gln non-synonymous polymorphism confers susceptibility to ankylosing spondylitis. Ann. Rheum. Dis. 67, 1451–1454 (2008).
Griffiths, C. E. et al. Comparison of ustekinumab and etanercept for moderate-to-severe psoriasis. N. Engl. J. Med. 362, 118–128 (2010).
Steinman, L. Mixed results with modulation of TH-17 cells in human autoimmune diseases. Nature Immunol. 11, 41–44 (2010).
Hurst, S. D. et al. New IL-17 family members promote Th1 or Th2 responses in the lung: in vivo function of the novel cytokine IL-25. J. Immunol. 169, 443–453 (2002).
Guo, P. et al. Dual nature of the adaptive immune system in lampreys. Nature 459, 796–801 (2009).
Franke, A. et al. Genome-wide association study for ulcerative colitis identifies risk loci at 7q22 and 22q13 (IL17REL). Nature Genet. 42, 292–294 (2010).
Fort, M. M. et al. IL-25 induces IL-4, IL-5, and IL-13 and Th2-associated pathologies in vivo. Immunity 15, 985–995 (2001).
Colonna, M. Interleukin-22-producing natural killer cells and lymphoid tissue inducer-like cells in mucosal immunity. Immunity 31, 15–23 (2009).
Acknowledgements
We thank M. McGeachy and K. Boniface for insightful discussion and R. Kastelein for critical reading of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Glossary
- Recombination-activating gene
-
(RAG). Expressed by developing lymphocytes, mice that are deficient for either Rag1 or Rag2 fail to produce B or T cells owing to a developmental block in the gene rearrangement that is necessary for receptor expression.
- γδ T cell
-
A T cell that expresses a TCR consisting of a γ-chain and a δ-chain. These T cells are present in the intestinal epithelium as intraepithelial lymphocytes. γδ T cells are also present in the lungs, skin, liver and other tissue sites during inflammation. γδ T cells are thought to have an important role in innate immune responses, particularly in mucosal tissues.
- Invariant natural killer T (iNKT) cell
-
A T cell that expresses a particular variable gene segment, Vα14 (in mice) and Vα 24 (in humans), precisely rearranged to a particular Jα (joining) gene segment to yield T cell receptor α-chains with an invariant sequence. Typically, these cells co-express cell-surface markers that are encoded by the natural killer (NK) locus, and they are activated by recognition of CD1d, particularly when α-galactosylceramide is bound in the groove of CD1d.
- Common cytokine-receptor γ-chain
-
A chain common to type I cytokine receptors. It was first discovered as the γ-chain of the IL-2 receptor and was subsequently shown also to be present in the receptors for IL-4, IL-7, IL-9, IL-15 and IL-21. It is mutated in humans with X-linked severe combined immunodeficiency.
- Ischaemia–reperfusion injury
-
An injury in which the tissue first suffers from hypoxia as a result of severely decreased, or completely arrested, blood flow. Restoration of normal blood flow then triggers inflammation, which exacerbates the tissue damage.
- Tight junction
-
A belt-like region of adhesion between adjacent epithelial or endothelial cells that regulates paracellular flux. Tight junction proteins include the integral membrane proteins occludin and claudin, in association with cytoplasmic zonula occludens proteins.
- Blood–brain barrier
-
A physiological barrier between blood vessels and brain parenchyma. It is formed by specialized tight junctions between endothelial cells of the blood vessel wall, which is surrounded by a basement membrane and an additional membrane formed from astrocyte feet and microglial cells, known as the glial limitin.
- IL-23ReGFP and RORγteGFP reporter mice
-
Mice expressing enhanced green fluorescent proteins (eGFP) under the control of the IL-23R (encoded by Il23a) or RORγt (encoded by Rorc) promoter. These mice can be used to track the differentiation, proliferation, migration and effector function of IL-17-producing cells. Gene targeting vectors are generated by introducing eGFP sequences into bacterial artificial chromosome (BAC) clones containing the Il23a or Rorc gene. The targeting constructs are introduced into embryonic stem cells and injected into mouse blastocysts to generate eGFP transgenic mice. The eGFP is co-expressed with IL-23R or RORγt, which allows cell fate analysis of IL-17-producing cells during an immune response.
Rights and permissions
About this article
Cite this article
Cua, D., Tato, C. Innate IL-17-producing cells: the sentinels of the immune system. Nat Rev Immunol 10, 479–489 (2010). https://doi.org/10.1038/nri2800
Published:
Issue Date:
DOI: https://doi.org/10.1038/nri2800
This article is cited by
-
Ribosomal protein S6 kinase 2 (RPS6KB2) is a potential immunotherapeutic target for cancer that upregulates proinflammatory cytokines
Molecular Biology Reports (2024)
-
IL-17A-mediated mitochondrial dysfunction induces pyroptosis in colorectal cancer cells and promotes CD8 + T-cell tumour infiltration
Journal of Translational Medicine (2023)
-
IL-17A promotes the progression of Alzheimer’s disease in APP/PS1 mice
Immunity & Ageing (2023)
-
Mechanisms of obesity- and diabetes mellitus-related pancreatic carcinogenesis: a comprehensive and systematic review
Signal Transduction and Targeted Therapy (2023)
-
Central role for neurally dysregulated IL-17A in dynamic networks of systemic and local inflammation in combat casualties
Scientific Reports (2023)
