
Abstract
Cells and tissues can undergo a variety of biological and structural changes in response to mechanical forces. Only a few existing techniques are available for quantification of structural changes at high resolution in response to forces applied along different directions. 3D-magnetic twisting cytometry (3D-MTC) is a technique for applying local mechanical stresses to living cells. Here we describe a protocol for interfacing 3D-MTC with confocal fluorescence microscopy. In 3D-MTC, ferromagnetic beads are bound to the cell surface via surface receptors, followed by their magnetization in any desired direction. A magnetic twisting field in a different direction is then applied to generate rotational shear stresses in any desired direction. This protocol describes how to combine magnetic-field-induced mechanical stimulation with confocal fluorescence microscopy and provides an optional extension for super-resolution imaging using stimulated emission depletion (STED) nanoscopy. This technology allows for rapid real-time acquisition of a living cell's mechanical responses to forces via specific receptors and for quantifying structural and biochemical changes in the same cell using confocal fluorescence microscopy or STED. The integrated 3D-MTC–microscopy platform takes ∼20 d to construct, and the experimental procedures require ∼4 d when carried out by a life sciences graduate student.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$259.00 per year
only $21.58 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout





Similar content being viewed by others

References
Discher, D.E., Mooney, D.J. & Zandstra, P.W. Growth factors, matrices, and forces combine and control stem cells. Science 324, 1673–1677 (2009).
Wang, N., Tytell, J.D. & Ingber, D.E. Mechanotransduction at a distance: mechanically coupling the extracellular matrix with the nucleus. Nat. Rev. Mol. Cell Biol. 1, 75–82 (2009).
Wozniak, M.A. & Chen, C.S. Mechanotransduction in development: a growing role for contractility. Nat. Rev. Mol. Cell Biol. 1, 34–43 (2009).
Humphrey, J.D., Dufresne, E.R. & Schwartz, M.A. Mechanotransduction and extracellular matrix homeostasis. Nat. Rev. Mol. Cell Biol. 12, 802–812 (2014).
Evans, E.A. & Hochmuth, R.M. Membrane viscoelasticity. Biophys. J. 1, 1–11 (1976).
Henderson, E., Haydon, P.G. & Sakaguchi, D.S. Actin filament dynamics in living glial cells imaged by atomic force microscopy. Science 5078, 1944–1946 (1992).
Hoh, J.H. & Schoenenberger, C.A. Surface morphology and mechanical properties of MDCK monolayers by atomic force microscopy. J. Cell Sci. 107, 1105–1114 (1994).
Matzke, R., Jacobson, K. & Radmacher, M. Direct, high-resolution measurement of furrow stiffening during division of adherent cells. Nat. Cell Biol. 6, 607–610 (2001).
Ashkin, A., Dziedzic, J.M. & Yamane, T. Optical trapping and manipulation of single cells using infrared laser beams. Nature 6150, 769–771 (1987).
Block, S.M., Goldstein, L.S. & Schnapp, B.J. Bead movement by single kinesin molecules studied with optical tweezers. Nature 6299, 348–352 (1990).
Dai, J. & Sheetz, M.P. Mechanical properties of neuronal growth cone membranes studied by tether formation with laser optical tweezers. Biophys. J. 3, 988–996 (1995).
Guck, J. et al. The optical stretcher: a novel laser tool to micromanipulate cells. Biophys. J. 2, 767–784 (2001).
Bausch, A.R., Ziemann, F., Boulbitch, A.A., Jacobson, K. & Sackmann, E. Local measurements of viscoelastic parameters of adherent cell surfaces by magnetic bead microrheometry. Biophys. J. 4, 2038–2049 (1998).
Wang, N., Butler, J.P. & Ingber, D.E. Mechanotransduction across the cell surface and through the cytoskeleton. Science 5111, 1124–1127 (1993).
Wang, N. & Ingber, D.E. Control of cytoskeletal mechanics by extracellular matrix, cell shape, and mechanical tension. Biophys. J. 66, 1281–1289 (1994).
Wang, N. & Ingber, D.E. Probing transmembrane mechanical coupling and cytomechanics using magnetic twisting cytometry. Biochem. Cell Biol. 73, 327–335 (1995).
Fabry, B. et al. Signal transduction in smooth muscle selected contribution: time course and heterogeneity of contractile responses in cultured human airway mouse melanoma B16 cells. J. Appl. Physiol. 2, 986–994 (2001).
Hu, S. et al. Mechanical anisotropy of adherent cells probed by a three dimensional magnetic twisting device. Am. J. Physiol. Cell Physiol. 287, C1184–C1191 (2004).
Tajik, A. et al. Transcription upregulation via force-induced direct stretching of chromatin. Nat. Mater. 15, 1287–1296 (2016).
Hell, S.W. & Wichmann, J. Breaking the diffraction resolution limit by stimulated emission: stimulated emission depletion microscopy. Opt. Lett. 19, 780–782 (1994).
Klar, T.A., Jakobs, S., Dyba, M., Egner, A. & Hell, S.W. Fluorescence microscopy with diffraction resolution barrier broken by stimulated emission. Proc. Natl. Acad. Sci. USA 97, 8206–8210 (2000).
Betzig, E. et al. Imaging intracellular fluorescent proteins at nanometer resolution. Science 313, 1642–1645 (2006).
Rust, M.J., Bates, M. & Zhuang, X. Sub-diffraction-limit imaging by stochastic optical reconstruction microscopy (STORM). Nat. Methods 3, 793–796 (2006).
Sahl, S.J., Leutenegger, M., Hilbert, M., Hell, S.W. & Eggeling, C. Fast molecular tracking maps nanoscale dynamics of plasma membrane lipids. Proc. Natl. Acad. Sci. USA 107, 6829–6834 (2010).
Patterson, G., Davidson, M., Manley, S. & Lippincott-Schwartz, J. Superresolution imaging using single-molecule localization. Annu. Rev. Phys. Chem. 61, 345–367 (2010).
Kittel, R.J. et al. Bruchpilot promotes active zone assembly, Ca2+ channel clustering, and vesicle release. Science 312, 1051–1054 (2006).
Balzarotti, F. et al. Nanometer resolution imaging and tracking of fluorescent molecules with minimal photon fluxes. Science. 355, 606–612 (2017).
Fabry, B. et al. Scaling the microrheology of living cells. Phys. Rev. Lett. 87, 148102 (2001).
Chowdhury, F. et al. Is cell rheology governed by nonequilibrium-to-equilibrium transition of noncovalent bonds? Biophys. J. 95, 5719–5727 (2008).
Heller, I. et al. STED nanoscopy combined with optical tweezers reveals protein dynamics on densely covered DNA. Nat. Methods 10, 910–916 (2013).
Neuman, K.C. & Block, S.M. Optical trapping. Rev. Sci. Instrum. 75, 2787–809 (2004).
Gutiérrez-Medina, B., Andreasson, J.O., Greenleaf, W.J., Laporta, A. & Block, S.M. An optical apparatus for rotation and trapping. Methods Enzymol. 475, 377–404 (2010).
Shroff, S.G., Saner, D.R. & Lal, R. Dynamic micromechanical properties of cultured rat atrial myocytes measured by atomic force microscopy. Am. J. Physiol. 269, C286–C292 (1995).
Raman, A. et al. Mapping nanomechanical properties of live cells using multi-harmonic atomic force microscopy. Nat. Nanotechnol. 6, 809–814 (2011).
Cartagena, A. & Raman, A. Local viscoelastic properties of live cells investigated using dynamic and quasi-static atomic force microscopy methods. Biophys. J. 106, 1033–1043 (2014).
Hubmayr, R.D. et al. Pharmacological activation changes stiffness of cultured human airway smooth muscle cells. Am. J. Physiol. Cell Physiol. 271, C1660–C1668 (1996).
Pourati, J. et al. Is cytoskeletal tension a major determinant of cell deformability in adherent endothelial cells? Am. J. Physiol. Cell Physiol. 274, C1283–C1289 (1998).
Wang, N. et al. Cell prestress. I. Stiffness and prestress are closely associated in adherent contractile cells. Am. J. Physiol. Cell Physiol. 282, C606–C616 (2002).
Puig-de-Morales, M. et al. Cytoskeletal mechanics in adherent human airway smooth muscle cells: probe specificity and scaling of protein-protein dynamics. Am. J. Physiol. Cell Physiol. 287, C643–C654 (2004).
An, S.S., Fabry, B., Trepat, X., Wang, N. & Fredberg, J.J. Do biophysical properties of the airway smooth muscle in culture predict airway hyperresponsiveness? Am. J. Respir. Cell Mol. Biol. 35, 55–64 (2006).
Berntsen, P. et al. Biomechanical effects of environmental and engineered particles on human airway smooth muscle cells. J. R. Soc. Interface 7, S331–S340 (2010).
Na, S. et al. Rapid signal transduction in living cells is a unique feature of mechanotransduction. Proc. Natl. Acad. Sci. USA 105, 6626–6631 (2008).
Na, S. & Wang, N. Application of fluorescence resonance energy transfer and magnetic twisting cytometry to quantify mechanochemical signaling activities in a living cell. Sci. Signal. 1, pl1 (2008).
Poh, Y.C. et al. Rapid activation of Rac GTPase in living cells by force is independent of Src. PLoS One 4, e7886 (2009).
Poh, Y.C. et al. Dynamic force-induced direct dissociation of protein complexes in a nuclear body in living cells. Nat. Commun. 3, 866 (2012).
Mijailovich, S.M., Kojic, M., Zivkovic, M., Fabry, B. & Fredberg, J.J. A finite element model of cell deformation during magnetic bead twisting. J. Appl. Physiol. 93, 1429–1436 (2002).
Craig, W.S., Cheng, S., Mullen, D.G., Blevitt, J. & Pierschbacher, M.D. Concept and progress in the development of RGD-containing peptide pharmaceuticals. Biopolymers 37, 157–175 (1995).
Tolić-Nørrelykke, I.M., Butler, J.P., Chen, J. & Wang, N. Spatial and temporal traction response in human airway smooth muscle cells. Am. J. Physiol. Cell Physiol. 283, C1254–C1266 (2002).
Hu, S., Chen, J., Butler, J.P. & Wang, N. Prestress mediates force propagation into the nucleus. Biochem. Biophys. Res. Commun. 329, 423–428 (2005).
Acknowledgements
We thank Y. Tan, S. Zhang, and H. Yi for technical assistance. This work was supported by the US National Institutes of Health (NIH) grant GM072744, Ministry of Science and Technology of China grant 2016YFA0101100, and the Fundamental Research Fund for the Central Universities (No. 2017KFQWJX002) from the Huazhong University of Science and Technology.
Author information
Authors and Affiliations
Contributions
N.W. conceived the project. N.W., Yuejin Z., and F.W. designed the experiments and analyzed the data. Y.-C.P. developed some analytical tools. Yuejin Z., F.W., Q.J., Junjian C., Junwei C., J.L., W.Y., W.Z., W.H., F.Y., and Yao Z. carried out the experiments. N.W., Yuejin Z., and F.W. wrote the manuscript. All authors discussed the results and implications and commented on the manuscript at all stages.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Integrated supplementary information
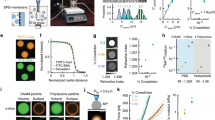
Supplementary Figure 1 Photographs of the 3D-MTC device and the confocal fluorescence microscopy.
I: The system consists of a Leica SP8-Confocal microscope (with an option of STED), a confocal PC, a 3D-MTC PC and a 3D-MTC; II to V: the steps of loading a cell-containing sample; VI: the current (magnetic field) control box for 3D-MTC.
Supplementary Figure 2 Schematic for the three-dimensional (3D) coils.
(a) Schematic for the auto temperature controller of 3D coils. The auto temperature controller provides a low flow rate of 37 ℃ air to mitigate the potential local heating problem. (b) The size of 3D coils and stage. X and Y coils’ internal diameters are 4.1 cm and their external diameters are 6 cm; Z coils’ internal diameters are 5.1 cm and their external diameters are 7 cm. The coils’ thicknesses are 1 cm each. The stage top dimension are 16 cm x 11 cm x 0.5 cm, bottom dimensions are 15 cm x 10 cm x 0.5 cm. (c) Top view of the Z coils and dish. The diameter of the cell culture dish is 3.5 cm. In the top view, the lateral distance from the dish boundary to the Z coil is 0.8 cm.
Supplementary Figure 3 Displacement quantification method.
(a) Flow chart of the image correlation method (ICM). (b) At each level the displacement field is estimated by using image A and image B. Image B is deformed by the displacement field calculated at the previous level. Succeeding levels have finer spatial resolution and smaller window size. Corr, calculation of cross-correlation; check, checkpoint; interp, interpolation48. (c) GFP-H2B fluorescence image. Nucleus is outlined with a dashed line. (d) Displacement map image of the same nucleus.
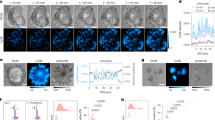
Supplementary Figure 4 Nuclear H2B displacements influenced by stress directionality.
A different cell from that in Fig. 4. From left to right: bright-field images of the cell, GFP-Histone 2B fluorescence images, and displacement map images of the same murine melanoma B16 cell. Nucleus is outlined with a dashed line. The thick white arrows point to the bead; the red arrows indicate direction of the bead displacement; the color bar indicates the displacement magnitude. (a) No applied stress (non-magnetized); (b) Magnetizing in Z axis and apply a 15 Pa torque (0.1Hz) in Y axis bright-field images; (c) Magnetizing in Z axis and apply a 15 Pa torque (0.1 Hz) in X axis bright-field images; (d) Magnetizing along Z axis and apply a 9 Pa along X axis and 12 Pa along Y axis (the resultant stress is 15 Pa and stress angle is 36.9°). (e) Magnetizing along Z axis and apply a 10.6 Pa along X axis and 10.6 Pa along Y axis (the resultant stress is 15 Pa and stress angle is 45°). (f) Magnetizing along Z axis and apply a 12 Pa along X axis and 9 Pa along Y axis (the resultant stress is 15 Pa and stress angle is 53.1°). The white square in each displacement map is enlarged and presented on the right (the 4th image in each row) to show the differences of the displacements between each loading direction (a-f) at the same loading magnitude. It is apparent that stress angles (loading directions) are important in resulting major differences in GFP-H2B displacements. The thin white arrow (150 nm) provides a scale for the magnitudes of the red arrows. Image acquisition time is 1 second per image. Scale bars, 7.5 μm. (g) Computed cell stiffnesses (complex moduli) for this cell at different forcing directions.at different forcing directions.
Supplementary Figure 5 H2B strains depend on force directions.
This is the same cell as in Supplementary Fig. 4. The bulk strains and shear strains were computed from displacement maps of the nucleus in Supplementary Fig. 4. (a) Stress angle of 0°. (b) Stress angle of 36.9°. (c) Stress angle of 45°. (d) Stress angle of 53.1°. (e) Stress angle of 90°. It is obvious that strain maps are different, with the 90°-stress angle generating highest strains.
Supplementary Figure 6 Improved displacements quantification via 3D-MTC with STED.
A living CHO cell was plated on collagen-1 coated dish overnight. A magnetizing field was applied along Z-axis and a twisting field was applied along X-axis (the short axis of the cell). The top panel is the data of GFP-H2B of the nucleus via a confocal microscope. The bottom panel is the data of the same cell under the same stress conditions quantified with a STED. Parameters for STED are: STED laser power (90 %) is 810 mW; the STED laser is CW (continuous wave) phase pattern; the excitation laser wavelength is 488nm; the STED depletion laser wavelength is 592 nm; scanning method is beam scanning. The scanning speed is 1000 Hz (1000 lines per sec). The 90 % STED power was estimated to be reduced to ~64.8 mW at the objective lens. (a) The GFP-H2B image of whole nucleus. The white box is the enlarged area. The top left of confocal image is the brightfield image of the same cell, the black dot is an RGD-coated magnetic bead. (b) Enlarged GFP-H2B images. (c) Full Width at Half Maximum (FWHM) of the fluorescence images. Confocal FWHM is 251 nm; STED FWHM is 118 nm. (d) Histograms of the fluorescence counts (with a Gaussian fit) at various FWHMs using confocal or STED. FWHM of confocal is 251.3 ± 1.03 nm, of STED is 118.04 ± 0.55 nm; mean ± s.e.m., 100 GFP-H2B spots were measured that were near the GFP-LacI spots next to the DHFR gene locus19. P<0.001 when comparing FWHM of confocal with that of STED. (e) A 15-Pa stress was applied at 0.3 Hz and peak displacement map was computed (displacement). The white box is the enlarged area. (f) Enlarged displacement maps. The black arrow (150 nm) represents the scale for the displacement magnitude. Image acquisition time is 320 millisecond per image. Comparing the white box of H2B fluorescence via STED with that via confocal, one could see better spatial resolution of the GFP-H2B images. FWHM of the PSF is 118 nm, better than the ~185 nm resolution using the re-scan confocal microscopy1. Comparing the white box of the displacement map via STED with that via confocal, improved displacements were also noticed. Several other cells showed similar trends.
Supplementary Figure 7 Lateral resolutions of confocal microscopy and STED.
Full Width at Half Maximum (FWHM) of Point Spread Function (PSF) is defined as Lateral resolution. 0%, confocal fluorescence microscopy only; 30%, 30 % depletion laser power of 270 mW (reduced to ~21.6 mW at the objective lens); 60 %, 60 % depletion laser power of 540 mW (reduced to ~43.2 mW the objective lens); 90%, 90 % depletion laser power of 810 mW (reduced to ~64.8 mW at the objective lens). Mean + s.e.m.; n= 11 cells (each data point is an average value from multiple GFP-H2B measurements near the DHFR gene locus in a cell); the same cells were imaged by confocal microscopy first and then by STED. 3 independent experiments. P <0.001 between data from any two laser powers. The line is a fit from the inverse square root law.
Supplementary Figure 8 Nuclear H2B fluorescence images and displacement maps under different conditions.
This cell is a representative cell in Supplementary Fig. 7. First row: brightfield images. White dashed lines are outlines of the nucleus. White arrow points to the magnetic bead. Second row: H2B fluorescence images. Note that the magnetic bead did not block the optical path because the bead was not in the optical path between the laser and the sample. Third row: H2B displacement maps when 15 Pa stress at 0.3125 Hz was applied. The pink arrow represents the magnitude and direction of the magnetic bead displacement. Bottom row: enlarged displacement maps corresponding to the white boxes above. (a) No-applied-stress (non-magnetized) confocal images. (b-e) Magnetizing in Z axis and applying a twisting field to induce 15 Pa stress at 0.3125 Hz along X-axis (the short axis of the cell); (b) image obtained with confocal microscopy. Full Width Half Maximum (FWHM) of the fluorescence image PSF is 279 nm; (c) image with 30% STED power (~21.6 mW). The FWHM is 198 nm; (d) image with 60% STED power (~43.2 mW). The FWHM is 157 nm; (e) image with 90% STED power (810 mW) (reduced to ~64.8 mW at the objective lens). The FWHM is 124 nm. Image acquisition time is 320 ms per image. Scale bars, 5 μm.
Supplementary Figure 9 STED power did not induce magnetic bead displacement.
From left to right: bright-field images of the cell, GFP-Histone 2B fluorescence images, displacement map images and enlarged displacement maps. Nucleus is outlined with a dashed line. The thick white arrows point to the magnetized bead; the pink dot indicates the magnitude and direction of the bead displacement (<5 nm, suggesting no additional displacements by STED); the color bar indicates the displacement magnitude. (a) No applied stress in confocal images. The FWHM is 236 nm; (b) the same nucleus in 90% STED power (~64.8 mW) images. The FWHM is 120 nm. Image acquisition time is 320 ms per image. Scale bars, 5 μm.
Supplementary Figure 10 System description of 3D-MTC with confocal fluorescence microscopy.
The platform of interfacing 3D-Magnetic Twisting Cytometry (3D-MTC) with the confocal fluorescence microscopy (with an option of stimulated emission depletion (STED) nanoscopy) includes three parts: magnetic field generator subsystem, monitoring and analysis subsystem, and observing subsystem. Each subsystem includes hardware equipment, interface board, control interface program or analysis modules.
Supplementary information
Supplementary Text and Figures
Supplementary Figures 1–10 and the Supplementary Method. (PDF 1666 kb)
Rights and permissions
About this article

Cite this article
Zhang, Y., Wei, F., Poh, YC. et al. Interfacing 3D magnetic twisting cytometry with confocal fluorescence microscopy to image force responses in living cells. Nat Protoc 12, 1437–1450 (2017). https://doi.org/10.1038/nprot.2017.042
Published:
Issue Date:
DOI: https://doi.org/10.1038/nprot.2017.042
This article is cited by
-
Brillouin microscopy monitors rapid responses in subcellular compartments
PhotoniX (2024)
-
Mapping mechanical properties of biological materials via an add-on Brillouin module to confocal microscopes
Nature Protocols (2021)
-
Optoregulated force application to cellular receptors using molecular motors
Nature Communications (2021)
-
Applications of atomic force microscopy in immunology
Frontiers of Medicine (2021)
-
Miniaturized magnetic stir bars for controlled agitation of aqueous microdroplets
Scientific Reports (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
