
Abstract
How do we time hand closure to catch a ball? Binocular disparity and optical looming provide two sources of information about an object's motion in depth, but the relative effectiveness of the two cues depends on ball size. Based on results from a virtual reality ball–catching task, we derive a simple model that uses both cues. The model is sensitive to the relative effectiveness of size and disparity and implicitly switches its response to the cue that specifies the earliest arrival and away from a cue that is lost or below threshold. We demonstrate the model's robustness by predicting the response of participants to some very unusual ball trajectories in a virtual reality task.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others
References
Sun, H. & Frost, B. J. Computation of different optical variables of looming objects in pigeon nucleus rotundus neurons. Nat. Neurosci. 4, 296–303 (1998).
Regan, D. & Beverley, K. I. Binocular and monocular stimuli for motion in depth: Changing–disparity and changing–size feed the same motion in depth stage. Vision Res. 19, 1331–1342 (1979).
Lee, D. N. A theory of visual control of braking based on information about time–to–collision. Perception 5, 437–459 (1976).
Regan, D. & Hamstra, S. J. Dissociation of discrimination thresholds for time to contact and for rate of angular expansion. Vision Res. 33, 447–462 (1993).
Heuer, H. Estimates of time to contact based on changing size and changing target vergence. Perception 22, 549–563 (1993).
Gray, R. & Regan, D. Accuracy of estimating time to collision using binocular and monocular information. Vision Res. 38, 499–512 (1997).
Wann, J. P. Anticipating arrival: is the tau–margin a specious theory? J. Exp Psychol. Hum. Percept. Perform. 22, 1031– 1048 (1996).
Tresilian, J. R. Perceptual and motor processes in interceptive timing. Hum. Mov. Sci 13, 335–373 ( 1994).
Laurent, M., Montagne, G. & Durey, A. Binocular invariants in interception tasks: a directed perception approach. Perception 25, 1437 –1450 (1996).
Regan, D., Erkelens C. J. & Collewijn, H. Necessary conditions for the perception of motion in depth. Invest. Ophthalmol. Vision Sci. 27, 584–597 (1986).
Milner, A. D. & Goodale, M. A. The Visual Brain in Action (Oxford Univ. Press, Oxford, 1996).
Alderson, G. J. K., Sully, D. J. & Sully, H. G. An operational analysis of a one–handed catching task using high–speed photography. J. Motor Behav. 6, 217–226 (1974).
Tresilian, J. R. Perceptual and cognitive processes in time–to–contact estimation: Analysis of prediction–motion and relative judgment tasks. Percept. Psychophys. 57, 231–245 (1995).
Landy, M. S., Maloney, L. T., Johnston, E. B. & Young, M. J. Measurement and modeling of depth cue combination: In defense of weak fusion. Vision Res. 35, 389–412 (1995).
Dobbins, A. C., Jeo, R. M., Fiser, J. & Allman, J. M. Distance modulation of neural activity in visual cortex. Science 281, 552–555 (1998).
Julesz, B. Textons, the elements of texture perception and their interactions. Nature 290, 91–97 ( 1981).
Regan, D. Visual judgements and misjudgements in cricket, and the art of flight. Perception 21, 91–115 ( 1992).
Peper, C. L., Bootsma, R. J., Mestre, D. R. & Bakker, F. C. Catching balls: How to get the hand to the right place at the right time. J. Exp Psychol. Hum. Percept. Perform. 20, 591–612 (1994).
Acknowledgements
We thank Anna Plooy and Martin Smyth for their input to the second experiment and also Julie Harris, Mike Harris and Graham Schafer for incisive comments. This research was supported by the UK EPSRC GR/L18693 and GR/L16125.
Author information
Authors and Affiliations
Corresponding author
Supplementary information
Supplementary Information
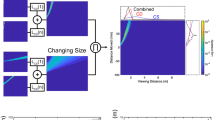
Three models that combine binocular and monocular information to estimate time to contact (TTC). (a) Fixed weighting of optic size and disparity. TTC is estimated from each input and then averaged on the basis of some weighting (0 <β < 1). This approach, however, is not robust if one input is perturbed or lost, and so it will produce large errors of estimation. (b) Variable weighting of optic size and disparity. Each modular estimate is first summated (Σ) and then a cross ratio from the opposing input is used to calculate the β weightings (see Results). (c) The dipole model. Binocular and monocular motion is combined before estimating TTC. If one of the inputs is perturbed or drops out, this approach has the advantages of model (b) without the explicit iterative calculation of β weights in flight. Note that in all three models the recovery of the binocular angular subtense φ would be required for a veridical estimate of TTC from disparity, but for many natural environments relative disparity α is approximately equal to φ (see Mathematical Appendix). (GIF 26 kb)
Rights and permissions
About this article
Cite this article
Rushton, S., Wann, J. Weighted combination of size and disparity: a computational model for timing a ball catch. Nat Neurosci 2, 186–190 (1999). https://doi.org/10.1038/5750
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1038/5750
This article is cited by
-
Image statistics determine the integration of visual cues to motion-in-depth
Scientific Reports (2022)
-
Emotional effects on time-to-contact judgments: arousal, threat, and fear of spiders modulate the effect of pictorial content
Experimental Brain Research (2014)
-
Eye movements influence estimation of time-to-contact in prediction motion
Experimental Brain Research (2010)
-
Visuo-motor coordination and internal models for object interception
Experimental Brain Research (2009)
-
Estimating time to contact during pursuit eye movements: Comparison between geometric model prediction and human performance
Optical Review (2008)

{kind=link}