
Abstract
Ion channels are essential for the regulation of neuronal functions. The significance of plasma membrane, mitochondrial, endoplasmic reticulum and lysosomal ion channels in the regulation of Ca2+ is well established. In contrast, surprisingly little is known about the function of ion channels on the nuclear envelope (NE). Here we demonstrate the presence of functional large-conductance, calcium-activated potassium channels (BK channels) on the NE of rodent hippocampal neurons. Functionally, blockade of nuclear BK channels (nBK channels) induces NE-derived Ca2+ release, nucleoplasmic Ca2+ elevation and cyclic AMP response element binding protein (CREB)-dependent transcription. More importantly, blockade of nBK channels regulates nuclear Ca2+–sensitive gene expression and promotes dendritic arborization in a nuclear Ca2+–dependent manner. These results suggest that the nBK channel functions as a molecular link between neuronal activity and nuclear Ca2+ to convey signals from synapse to nucleus and is a new modulator, operating at the NE, of synaptic activity–dependent neuronal functions.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout





Similar content being viewed by others


References
Wheeler, D.G. et al. CaV1 and CaV2 channels engage distinct modes of Ca2+ signaling to control CREB-dependent gene expression. Cell 149, 1112–1124 (2012).
Jiang, D., Zhao, L. & Clapham, D.E. Genome-wide RNAi screen identifies Letm1 as a mitochondrial Ca2+/H+ antiporter. Science 326, 144–147 (2009).
Calcraft, P.J. et al. NAADP mobilizes calcium from acidic organelles through two-pore channels. Nature 459, 596–600 (2009).
Dong, X.P. et al. The type IV mucolipidosis-associated protein TRPML1 is an endolysosomal iron release channel. Nature 455, 992–996 (2008).
Wegierski, T. et al. TRPP2 channels regulate apoptosis through the Ca2+ concentration in the endoplasmic reticulum. EMBO J. 28, 490–499 (2009).
Bootman, M.D., Fearnley, C., Smyrnias, I., MacDonald, F. & Roderick, H.L. An update on nuclear calcium signalling. J. Cell Sci. 122, 2337–2350 (2009).
Carafoli, E., Nicotera, P. & Santella, L. Calcium signalling in the cell nucleus. Cell Calcium 22, 313–319 (1997).
Gerasimenko, O.V., Gerasimenko, J.V., Tepikin, A.V. & Petersen, O.H. ATP-dependent accumulation and inositol trisphosphate- or cyclic ADP-ribose-mediated release of Ca2+ from the nuclear envelope. Cell 80, 439–444 (1995).
Leite, M.F. et al. Nuclear and cytosolic calcium are regulated independently. Proc. Natl. Acad. Sci. USA 100, 2975–2980 (2003).
Stehno-Bittel, L., Luckhoff, A. & Clapham, D.E. Calcium release from the nucleus by InsP3 receptor channels. Neuron 14, 163–167 (1995).
Bading, H. Nuclear calcium signalling in the regulation of brain function. Nat. Rev. Neurosci. 14, 593–608 (2013).
Zhang, S.J. et al. Nuclear calcium signaling controls expression of a large gene pool: identification of a gene program for acquired neuroprotection induced by synaptic activity. PLoS Genet. 5, e1000604 (2009).
Mauceri, D., Freitag, H.E., Oliveira, A.M., Bengtson, C.P. & Bading, H. Nuclear calcium-VEGFD signaling controls maintenance of dendrite arborization necessary for memory formation. Neuron 71, 117–130 (2011).
Limbäck-Stokin, K., Korzus, E., Nagaoka-Yasuda, R. & Mayford, M. Nuclear calcium/calmodulin regulates memory consolidation. J. Neurosci. 24, 10858–10867 (2004).
Papadia, S., Stevenson, P., Hardingham, N.R., Bading, H. & Hardingham, G.E. Nuclear Ca2+ and the cAMP response element-binding protein family mediate a late phase of activity-dependent neuroprotection. J. Neurosci. 25, 4279–4287 (2005).
Misonou, H. et al. Immunolocalization of the Ca2+-activated K+ channel Slo1 in axons and nerve terminals of mammalian brain and cultured neurons. J. Comp. Neurol. 496, 289–302 (2006).
Douglas, R.M. et al. The calcium-sensitive large-conductance potassium channel (BK/MAXI K) is present in the inner mitochondrial membrane of rat brain. Neuroscience 139, 1249–1261 (2006).
Singh, H. et al. MitoBKCa is encoded by the Kcnma1 gene, and a splicing sequence defines its mitochondrial location. Proc. Natl. Acad. Sci. USA 110, 10836–10841 (2013).
Gong, L.W., Gao, T.M., Huang, H., Zhuang, Z.Y. & Tong, Z. Transient forebrain ischemia induces persistent hyperactivity of large conductance Ca2+-activated potassium channels via oxidation modulation in rat hippocampal CA1 pyramidal neurons. Eur. J. Neurosci. 15, 779–783 (2002).
Hu, H. et al. Presynaptic Ca2+-activated K+ channels in glutamatergic hippocampal terminals and their role in spike repolarization and regulation of transmitter release. J. Neurosci. 21, 9585–9597 (2001).
Lee, U.S. & Cui, J. BK channel activation: structural and functional insights. Trends Neurosci. 33, 415–423 (2010).
Salkoff, L., Butler, A., Ferreira, G., Santi, C. & Wei, A. High-conductance potassium channels of the SLO family. Nat. Rev. Neurosci. 7, 921–931 (2006).
Fricker, M., Hollinshead, M., White, N. & Vaux, D. Interphase nuclei of many mammalian cell types contain deep, dynamic, tubular membrane-bound invaginations of the nuclear envelope. J. Cell Biol. 136, 531–544 (1997).
Quesada, I. et al. Nuclear KATP channels trigger nuclear Ca2+ transients that modulate nuclear function. Proc. Natl. Acad. Sci. USA 99, 9544–9549 (2002).
Wu, B., Yamaguchi, H., Lai, F.A. & Shen, J. Presenilins regulate calcium homeostasis and presynaptic function via ryanodine receptors in hippocampal neurons. Proc. Natl. Acad. Sci. USA 110, 15091–15096 (2013).
Deisseroth, K., Mermelstein, P.G., Xia, H. & Tsien, R.W. Signaling from synapse to nucleus: the logic behind the mechanisms. Curr. Opin. Neurobiol. 13, 354–365 (2003).
Han, M.H. et al. Role of cAMP response element-binding protein in the rat locus ceruleus: regulation of neuronal activity and opiate withdrawal behaviors. J. Neurosci. 26, 4624–4629 (2006).
Sheng, M., Thompson, M.A. & Greenberg, M.E. CREB: a Ca2+-regulated transcription factor phosphorylated by calmodulin-dependent kinases. Science 252, 1427–1430 (1991).
Bito, H., Deisseroth, K. & Tsien, R.W. CREB phosphorylation and dephosphorylation: a Ca2+- and stimulus duration-dependent switch for hippocampal gene expression. Cell 87, 1203–1214 (1996).
Xing, J., Ginty, D.D. & Greenberg, M.E. Coupling of the RAS-MAPK pathway to gene activation by RSK2, a growth factor-regulated CREB kinase. Science 273, 959–963 (1996).
Hardingham, G.E., Arnold, F.J. & Bading, H. Nuclear calcium signaling controls CREB-mediated gene expression triggered by synaptic activity. Nat. Neurosci. 4, 261–267 (2001).
Rodrigues, M.A. et al. Nucleoplasmic calcium is required for cell proliferation. J. Biol. Chem. 282, 17061–17068 (2007).
Lin, Y. et al. Activity-dependent regulation of inhibitory synapse development by Npas4. Nature 455, 1198–1204 (2008).
Ramamoorthi, K. et al. Npas4 regulates a transcriptional program in CA3 required for contextual memory formation. Science 334, 1669–1675 (2011).
Meredith, A.L., Thorneloe, K.S., Werner, M.E., Nelson, M.T. & Aldrich, R.W. Overactive bladder and incontinence in the absence of the BK large conductance Ca2+-activated K+ channel. J. Biol. Chem. 279, 36746–36752 (2004).
Meredith, A.L. et al. BK calcium-activated potassium channels regulate circadian behavioral rhythms and pacemaker output. Nat. Neurosci. 9, 1041–1049 (2006).
Sausbier, M. et al. Cerebellar ataxia and Purkinje cell dysfunction caused by Ca2+-activated K+ channel deficiency. Proc. Natl. Acad. Sci. USA 101, 9474–9478 (2004).
Xu, W. et al. Cytoprotective role of Ca2+- activated K+ channels in the cardiac inner mitochondrial membrane. Science 298, 1029–1033 (2002).
Mazzanti, M., Bustamante, J.O. & Oberleithner, H. Electrical dimension of the nuclear envelope. Physiol. Rev. 81, 1–19 (2001).
Dick, D.A. The distribution of sodium, potassium and chloride in the nucleus and cytoplasm of Bufo bufo oocytes measured by electron microprobe analysis. J. Physiol. (Lond.) 284, 37–53 (1978).
Garner, M.H. Na,K-ATPase in the nuclear envelope regulates Na+: K+ gradients in hepatocyte nuclei. J. Membr. Biol. 187, 97–115 (2002).
Gifford, J.D., Galla, J.H., Luke, R.G. & Rick, R. Ion concentrations in the rat CCD: differences between cell types and effect of alkalosis. Am. J. Physiol. 259, F778–F782 (1990).
Rick, R. pHi determines rate of sodium transport in frog skin: results of a new method to determine pHi. Am. J. Physiol. 266, F367–F374 (1994).
Endo, M. Calcium-induced calcium release in skeletal muscle. Physiol. Rev. 89, 1153–1176 (2009).
Chen, S.R., Li, X., Ebisawa, K. & Zhang, L. Functional characterization of the recombinant type 3 Ca2+ release channel (ryanodine receptor) expressed in HEK293 cells. J. Biol. Chem. 272, 24234–24246 (1997).
Zahradníková, A. & Meszaros, L.G. Voltage change-induced gating transitions of the rabbit skeletal muscle Ca2+ release channel. J. Physiol. (Lond.) 509, 29–38 (1998).
van Welie, I. & du Lac, S. Bidirectional control of BK channel open probability by CAMKII and PKC in medial vestibular nucleus neurons. J. Neurophysiol. 105, 1651–1659 (2011).
Loane, D.J., Hicks, G.A., Perrino, B.A. & Marrion, N.V. Inhibition of BK channel activity by association with calcineurin in rat brain. Eur. J. Neurosci. 24, 433–441 (2006).
Shamloo, M. et al. Npas4, a novel helix-loop-helix PAS domain protein, is regulated in response to cerebral ischemia. Eur. J. Neurosci. 24, 2705–2720 (2006).
Huang, L., Li, B., Li, W., Guo, H. & Zou, F. ATP-sensitive potassium channels control glioma cells proliferation by regulating ERK activity. Carcinogenesis 30, 737–744 (2009).
Avdonin, V., Tang, X.D. & Hoshi, T. Stimulatory action of internal protons on Slo1 BK channels. Biophys. J. 84, 2969–2980 (2003).
Tang, X.D. et al. Haem can bind to and inhibit mammalian calcium-dependent Slo1 BK channels. Nature 425, 531–535 (2003).
Taylor, C.W. et al. IP3 receptors: some lessons from DT40 cells. Immunol. Rev. 231, 23–44 (2009).
Cao, X. et al. Astrocyte-derived ATP modulates depressive-like behaviors. Nat. Med. 19, 773–777 (2013).
Woo, R.S. et al. Neuregulin-1 enhances depolarization-induced GABA release. Neuron 54, 599–610 (2007).
Chen, Y.J. et al. ErbB4 in parvalbumin-positive interneurons is critical for neuregulin 1 regulation of long-term potentiation. Proc. Natl. Acad. Sci. USA 107, 21818–21823 (2010).
Li, X.M. et al. Contribution of downregulation of L-type calcium currents to delayed neuronal death in rat hippocampus after global cerebral ischemia and reperfusion. J. Neurosci. 27, 5249–5259 (2007).
Hristov, K.L., Chen, M., Kellett, W.F., Rovner, E.S. & Petkov, G.V. Large-conductance voltage- and Ca2+-activated K+ channels regulate human detrusor smooth muscle function. Am. J. Physiol. Cell Physiol. 301, C903–C912 (2011).
Zhuang, X., Semenova, E., Maric, D. & Craigie, R. Dephosphorylation of barrier-to-autointegration factor by protein phosphatase 4 and its role in cell mitosis. J. Biol. Chem. 289, 1119–1127 (2014).
Muller, F.L. et al. Passenger deletions generate therapeutic vulnerabilities in cancer. Nature 488, 337–342 (2012).
Cameron, P.H., Chevet, E., Pluquet, O., Thomas, D.Y. & Bergeron, J.J. Calnexin phosphorylation attenuates the release of partially misfolded alpha1-antitrypsin to the secretory pathway. J. Biol. Chem. 284, 34570–34579 (2009).
Obexer, P. et al. p16INK4A sensitizes human leukemia cells to FAS- and glucocorticoid-induced apoptosis via induction of BBC3/Puma and repression of MCL1 and BCL2. J. Biol. Chem. 284, 30933–30940 (2009).
Zaru, R., Ronkina, N., Gaestel, M., Arthur, J.S. & Watts, C. The MAPK-activated kinase Rsk controls an acute Toll-like receptor signaling response in dendritic cells and is activated through two distinct pathways. Nat. Immunol. 8, 1227–1235 (2007).
Gehani, S.S. et al. Polycomb group protein displacement and gene activation through MSK-dependent H3K27me3S28 phosphorylation. Mol. Cell 39, 886–900 (2010).
Tai, Y. et al. TRPC6 channels promote dendritic growth via the CaMKIV-CREB pathway. J. Cell Sci. 121, 2301–2307 (2008).
Muller, M., Cardenas, C., Mei, L., Cheung, K.H. & Foskett, J.K. Constitutive cAMP response element binding protein (CREB) activation by Alzheimer's disease presenilin-driven inositol trisphosphate receptor (InsP3R) Ca2+ signaling. Proc. Natl. Acad. Sci. USA 108, 13293–13298 (2011).
Yu, C. et al. Pharmacologic mitogen-activated protein/extracellular signal-regulated kinase kinase/mitogen-activated protein kinase inhibitors interact synergistically with STI571 to induce apoptosis in Bcr/Abl-expressing human leukemia cells. Cancer Res. 62, 188–199 (2002).
Fei, Z., Bera, T.K., Liu, X., Xiang, L. & Pastan, I. Ankrd26 gene disruption enhances adipogenesis of mouse embryonic fibroblasts. J. Biol. Chem. 286, 27761–27768 (2011).
Wei, F., Scholer, H.R. & Atchison, M.L. Sumoylation of Oct4 enhances its stability, DNA binding, and transactivation. J. Biol. Chem. 282, 21551–21560 (2007).
Walton, P.D. et al. Ryanodine and inositol trisphosphate receptors coexist in avian cerebellar Purkinje neurons. J. Cell Biol. 113, 1145–1157 (1991).
Beliveau, F. et al. Essential role of endocytosis of the type II transmembrane serine protease TMPRSS6 in regulating its functionality. J. Biol. Chem. 286, 29035–29043 (2011).
Acknowledgements
We thank X. Tang (Nankai University) for BK-GFP plasmid, M. Nathanson (Yale University) for PV-NLS plasmid, H. Bading (University of Heidelberg) for CaMKIVK75E and CaMBP4 plasmids, and F.-X. Liang and the Microscopy Core of New York University Medical Center for immunoelectron microscopy analysis. This work was supported by the National Natural Science Foundation of China (grants 81030022, 81329003 and U1201225 to T.-M.G., 30900581 to B.L., 81001129 to L.H.), the Program for Changjiang Scholars and Innovative Research Team (grant IRT1142), Guangdong Natural Science Foundation (grants 9351051501000003 and CXZD1018 to T.-M.G., 10451051501004726 to L.H.), Guangzhou Science and Technology Project (grant 7411802013939), Medical Scientific Research Foundation of Guangdong Province (B2010170 to L.H.) and the US National Heart, Lung, and Blood Institute (grant R01-HL102758 to A.L.M.).
Author information
Authors and Affiliations
Contributions
B.L. and T.-M.G. designed the study; B.L., W.J. and L.H. conducted biochemistry, Ca2+ imaging and immunocytochemistry experiments; B.L., P.W. and Z.L. conducted electrophysiological experiments; B.L. and S.L. cultured neurons; A.K.F., A.L.M., M.-H.H. and X.-H.Z. contributed new reagents or analytic tools; B.L., W.J., P.W., Z.L. and L.H. collected and analyzed data; B.L., L.H. and T.-M.G. wrote the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Integrated supplementary information
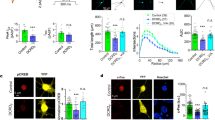
Supplementary Figure 1 BK β4 subunits are expressed in the nuclear envelope.
The distribution of BK channel β4 subunits (red) was examined by confocal immunofluorescence in cultured hippocampal neurons. Intracellular ring-like labeling around the nucleus was observed and colocalized with lamin B (green). Scale bar represents 20 μm.
Supplementary Figure 2 The effects of BK channel shRNAs on nBK channel expression and nuclear Ca2+ signal.
(a) BK channel expression after knockdown by two pairs of lentiviral shRNAs. (b) Paxilline-induced changes in Fluo-4 AM fluorescence intensity in the isolated nuclei after knockdown of BK channel (n = 25 nuclei for each group, unpaired t-test, P = 1.83 × 10−7, t48 = 6.09). (c) Paxilline-induced changes in Fluo-4 dextran fluorescence intensity in the isolated nuclei after knockdown of BK channel (n = 25 nuclei for each group, unpaired t-test, P = 2.67 × 10−7, t48 = 5.98). The bottom and top of the box represent the first and third quartiles, and the band inside the box is the median. The whisker represents the minimum and maximum of all of the data. **P < 0.01. The full-length blots for a are presented in Supplementary Figure 7.
Supplementary Figure 3 RyR expression was downregulated by RyR shRNA.
(a) RyRs mRNA level after knockdown by shRNAs. GAPDH was used as control. One-way ANOVA (N = 3 for each group, P = 9.47 × 10−6, F3,8 = 57.27) and post hoc test. (b) RyRs protein level after knockdown by shRNAs. Fisher's least significant difference test was used for post hoc test in one-way ANOVA. Error bars represent the mean ± s.e.m.. **P < 0.01. The full-length blots for b are presented in Supplementary Figure 7.
Supplementary Figure 4 The effects of IbTx on pmBK channel activity and expression.
(a) Outward potassium channel currents were recorded in whole-cell patch clamp to show that paxilline (10 μM) had no additive effect on pmBK channel blockade by IbTx (100 μM). The recording solution contains 1mM 4-AP, 1μM TTX, and 5μM glybenclamide. (b) The effects of IbTx (100 μM) and IbTx (100 μM)+paxilline (10 μM) on action potentials. IbTx broadened the action potential by blocking BK channels. Paxilline did not show additive effects. (c) Pre-incubation (10 min) of IbTx did not affect BK channel expression on plasma membrane. Membrane proteins were isolated as described in Methods section. Na/K ATPase was used as loading control. The full-length blots for c are presented in Supplementary Figure 7.
Supplementary Figure 5 Validation for shRNAs against nBK channel-regulated genes.
mRNA level of Fos, Npas4, Atf3, Btg2, Bcl6 and Ifi202b after knockdown by shRNAs as indicated. One-way ANOVA (N = 3 independent cultures from at least 3 litters for each group; P = 1.77 × 10−5, F2,6 = 112.05 for Fos; P = 1.70 × 10−6, F2,6 = 248.60 for Npas4; P = 2.47 × 10−5, F2,6 = 99.99 for Atf3; P = 1.09 × 10−5, F2,6 = 132.38 for Btg2; P = 2.84 × 10−5, F2,6 = 95.38 for Bcl6; P = 1.09 × 10−5, F2,6 = 132.29 for Ifi202b) and post hoc test. Fisher's least significant difference test was used for post hoc test in one-way ANOVA. Error bars represent the mean ± s.e.m..
Supplementary Figure 6 The effects of nBK channel-regulated genes on dendritic arborization.
(a) Representative micrographs of hippocampal neurons transfected with GFP-tagged plasmids containing gene-specific shRNAs or non-silencing shRNAs with the treatment of DMSO. (b) Sholl analysis (left) and quantification of total dendritic length (right) in hippocampal neurons treated as indicated. Right, one-way ANOVA (P = 0.042, F6,133 = 2.26) and post hoc test. N = 20 cells from 4 independent cultures from at least 4 litters for each group. Fisher's least significant difference test was used for post hoc test in one-way ANOVA. Error bars represent the mean ± s.e.m.. Scale bar = 30 μm. **P < 0.01. N.S., non-silencing shRNAs.
Supplementary information
Supplementary Text and Figures
Supplementary Figures 1–7 (PDF 1212 kb)
Rights and permissions
About this article

Cite this article
Li, B., Jie, W., Huang, L. et al. Nuclear BK channels regulate gene expression via the control of nuclear calcium signaling. Nat Neurosci 17, 1055–1063 (2014). https://doi.org/10.1038/nn.3744
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nn.3744
This article is cited by
-
Astrocytic lactate dehydrogenase A regulates neuronal excitability and depressive-like behaviors through lactate homeostasis in mice
Nature Communications (2023)
-
CD38/ADP-ribose/TRPM2-mediated nuclear Ca2+ signaling is essential for hepatic gluconeogenesis in fasting and diabetes
Experimental & Molecular Medicine (2023)
-
Neuronal nuclear calcium signaling suppression of microglial reactivity is mediated by osteoprotegerin after traumatic brain injury
Journal of Neuroinflammation (2022)
-
Inhibition of BKCa channels protects neonatal hearts against myocardial ischemia and reperfusion injury
Cell Death Discovery (2022)
-
Astrocyte-Derived Lactate Modulates the Passive Coping Response to Behavioral Challenge in Male Mice
Neuroscience Bulletin (2021)
