
Abstract
Huntington's disease (HD) is characterized by striatal medium spiny neuron (MSN) dysfunction, but the underlying mechanisms remain unclear. We explored roles for astrocytes, in which mutant huntingtin is expressed in HD patients and mouse models. We found that symptom onset in R6/2 and Q175 HD mouse models was not associated with classical astrogliosis, but was associated with decreased Kir4.1 K+ channel functional expression, leading to elevated in vivo striatal extracellular K+, which increased MSN excitability in vitro. Viral delivery of Kir4.1 channels to striatal astrocytes restored Kir4.1 function, normalized extracellular K+, ameliorated aspects of MSN dysfunction, prolonged survival and attenuated some motor phenotypes in R6/2 mice. These findings indicate that components of altered MSN excitability in HD may be caused by heretofore unknown disturbances of astrocyte-mediated K+ homeostasis, revealing astrocytes and Kir4.1 channels as therapeutic targets.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout








Similar content being viewed by others

References
Kuffler, S.W. Neuroglial cells: physiological properties and a potassium mediated effect of neuronal activity on the glial membrane potential. Proc. R. Soc. Lond. B Biol. Sci. 168, 1–21 (1967).
Barres, B.A. The mystery and magic of glia: a perspective on their roles in health and disease. Neuron 60, 430–440 (2008).
Ransom, B.R. & Ransom, C.B. Astrocytes: multitalented stars of the central nervous system. Methods Mol. Biol. 814, 3–7 (2012).
Maragakis, N.J. & Rothstein, J.D. Mechanisms of Disease: astrocytes in neurodegenerative disease. Nat. Clin. Pract. Neurol. 2, 679–689 (2006).
Ilieva, H., Polymenidou, M. & Cleveland, D.W. Non-cell autonomous toxicity in neurodegenerative disorders: ALS and beyond. J. Cell Biol. 187, 761–772 (2009).
Clarke, L.E. & Barres, B.A. Emerging roles of astrocytes in neural circuit development. Nat. Rev. Neurosci. 14, 311–321 (2013).
Mangiarini, L. et al. Exon 1 of the HD gene with an expanded CAG repeat is sufficient to cause a progressive neurological phenotype in transgenic mice. Cell 87, 493–506 (1996).
Cepeda, C., Cummings, D.M., André, V.M., Holley, S.M. & Levine, M.S. Genetic mouse models of Huntington's disease: focus on electrophysiological mechanisms. ASN Neuro. 2, e00033 (2010).
Shin, J.Y. et al. Expression of mutant huntingtin in glial cells contributes to neuronal excitotoxicity. J. Cell Biol. 171, 1001–1012 (2005).
Faideau, M. et al. In vivo expression of polyglutamine-expanded huntingtin by mouse striatal astrocytes impairs glutamate transport: a correlation with Huntington's disease subjects. Hum. Mol. Genet. 19, 3053–3067 (2010).
Bradford, J. et al. Expression of mutant huntingtin in mouse brain astrocytes causes age-dependent neurological symptoms. Proc. Natl. Acad. Sci. USA 106, 22480–22485 (2009).
Pouladi, M.A., Morton, A.J. & Hayden, M.R. Choosing an animal model for the study of Huntington's disease. Nat. Rev. Neurosci. 14, 708–721 (2013).
Menalled, L.B. et al. Comprehensive behavioral and molecular characterization of a new knock-in mouse model of Huntington's disease: zQ175. PLoS ONE 7, e49838 (2012).
Heikkinen, T. et al. Characterization of neurophysiological and behavioral changes, MRI brain volumetry and 1H MRS in zQ175 knock-in mouse model of Huntington's disease. PLoS ONE 7, e50717 (2012).
Sofroniew, M.V. Molecular dissection of reactive astrogliosis and glial scar formation. Trends Neurosci. 32, 638–647 (2009).
Higashi, K. et al. An inwardly rectifying K(+) channel, Kir4.1, expressed in astrocytes surrounds synapses and blood vessels in brain. Am. J. Physiol. Cell Physiol. 281, C922–C931 (2001).
Cahoy, J.D. et al. A transcriptome database for astrocytes, neurons, and oligodendrocytes: a new resource for understanding brain development and function. J. Neurosci. 28, 264–278 (2008).
Wang, C.E. et al. Suppression of neuropil aggregates and neurological symptoms by an intracellular antibody implicates the cytoplasmic toxicity of mutant huntingtin. J. Cell Biol. 181, 803–816 (2008).
Shigetomi, E., Tong, X., Kwan, K.Y., Corey, D.P. & Khakh, B.S. TRPA1 channels regulate astrocyte resting calcium and inhibitory synapse efficacy through GAT-3. Nat. Neuro. 15, 70–80 (2011).
Poopalasundaram, S. et al. Glial heterogeneity in expression of the inwardly rectifying K(+) channel, Kir4.1, in adult rat CNS. Glia 30, 362–372 (2000).
Barres, B.A., Koroshetz, W.J., Chun, L.L. & Corey, D.P. Ion channel expression by white matter glia: the type-1 astrocyte. Neuron 5, 527–544 (1990).
Chever, O., Djukic, B., McCarthy, K.D. & Amzica, F. Implication of Kir4.1 channel in excess potassium clearance: an in vivo study on anesthetized glial-conditional Kir4.1 knock-out mice. J. Neurosci. 30, 15769–15777 (2010).
Kofuji, P. & Newman, E.A. Potassium buffering in the central nervous system. Neuroscience 129, 1045–1056 (2004).
Furutani, K., Ohno, Y., Inanobe, A., Hibino, H. & Kurachi, Y. Mutational and in silico analyses for antidepressant block of astroglial inward-rectifier Kir4.1 channel. Mol. Pharmacol. 75, 1287–1295 (2009).
Su, S. et al. Inhibition of astroglial inwardly rectifying Kir4.1 channels by a tricyclic antidepressant, nortriptyline. J. Pharmacol. Exp. Ther. 320, 573–580 (2007).
Estrada-Sánchez, A.M. & Rebec, G.V. Corticostriatal dysfunction and glutamate transporter 1 (GLT1) in Huntington's disease: interactions between neurons and astrocytes. Basal Ganglia 2, 57–66 (2012).
Benn, C.L., Fox, H. & Bates, G.P. Optimisation of region-specific reference gene selection and relative gene expression analysis methods for pre-clinical trials of Huntington's disease. Mol. Neurodegener. 3, 17 (2008).
Strand, A.D. et al. Expression profiling of Huntington's disease models suggests that brain-derived neurotrophic factor depletion plays a major role in striatal degeneration. J. Neurosci. 27, 11758–11768 (2007).
Tang, T.S. et al. Huntingtin and huntingtin-associated protein 1 influence neuronal calcium signaling mediated by inositol-(1,4,5) triphosphate receptor type 1. Neuron 39, 227–239 (2003).
Rubinsztein, D.C. & Carmichael, J. Huntington's disease: molecular basis of neurodegeneration. Expert Rev. Mol. Med. 5, 1–21 (2003).
Adermark, L. & Lovinger, D.M. Electrophysiological properties and gap junction coupling of striatal astrocytes. Neurochem. Int. 52, 1365–1372 (2008).
Shigetomi, E. et al. Imaging calcium microdomains within entire astrocyte territories and endfeet with GCaMPs expressed using adeno-associated viruses. J. Gen. Physiol. 141, 633–647 (2013).
Ortinski, P.I. et al. Selective induction of astrocytic gliosis generates deficits in neuronal inhibition. Nat. Neurosci. 13, 584–591 (2010).
Xie, Y., Wang, T., Sun, G.Y. & Ding, S. Specific disruption of astrocytic Ca2+ signaling pathway in vivo by adeno-associated viral transduction. Neuroscience 170, 992–1003 (2010).
Hall, D.G. Ion-selective membrane electrodes: a general limiting treatment of interference effects. J. Phys. Chem. 100, 7230–7236 (1996).
Hille, B. Ion Channels of Excitable Membranes 3rd edn. (Sinauer Associates, 2001).
Klapstein, G.J. et al. Electrophysiological and morphological changes in striatal spiny neurons in R6/2 Huntington's disease transgenic mice. J. Neurophysiol. 86, 2667–2677 (2001).
Barres, B.A. Five electrophysiological properties of glial cells. Ann. NY Acad. Sci. 633, 248–254 (1991).
Herrmann, J.E. et al. STAT3 is a critical regulator of astrogliosis and scar formation after spinal cord injury. J. Neurosci. 28, 7231–7243 (2008).
Stack, E.C. et al. Chronology of behavioral symptoms and neuropathological sequela in R6/2 Huntington's disease transgenic mice. J. Comp. Neurol. 490, 354–370 (2005).
Behrens, P.F., Franz, P., Woodman, B., Lindenberg, K.S. & Landwehrmeyer, G.B. Impaired glutamate transport and glutamate-glutamine cycling: downstream effects of the Huntington mutation. Brain 125, 1908–1922 (2002).
Miller, B.R. et al. Up-regulation of GLT1 expression increases glutamate uptake and attenuates the Huntington's disease phenotype in the R6/2 mouse. Neuroscience 153, 329–337 (2008).
Bradford, J. et al. Mutant huntingtin in glial cells exacerbates neurological symptoms of Huntington disease mice. J. Biol. Chem. 285, 10653–10661 (2010).
Hamby, M.E. et al. Inflammatory mediators alter the astrocyte transcriptome and calcium signaling elicited by multiple G-protein–coupled receptors. J. Neurosci. 32, 14489–14510 (2012).
Zamanian, J.L. et al. Genomic analysis of reactive astrogliosis. J. Neurosci. 32, 6391–6410 (2012).
Ariano, M.A. et al. Striatal potassium channel dysfunction in Huntington's disease transgenic mice. J. Neurophysiol. 93, 2565–2574 (2005).
Ariano, M.A., Wagle, N. & Grissell, A.E. Neuronal vulnerability in mouse models of Huntington's disease: membrane channel protein changes. J. Neurosci. Res. 80, 634–645 (2005).
Djukic, B., Casper, K.B., Philpot, B.D., Chin, L.S. & McCarthy, K.D. Conditional knock-out of Kir4.1 leads to glial membrane depolarization, inhibition of potassium and glutamate uptake, and enhanced short-term synaptic potentiation. J. Neurosci. 27, 11354–11365 (2007).
Amzica, F. & Steriade, M. Neuronal and glial membrane potentials during sleep and paroxysmal oscillations in the neocortex. J. Neurosci. 20, 6648–6665 (2000).
Amzica, F. & Steriade, M. The functional significance of K-complexes. Sleep Med. Rev. 6, 139–149 (2002).
Acknowledgements
The authors thank P. Kofuji (University of Minnesota) for sharing Kir4.1-GFP plasmids, R. Korsak for coordinating the genotyping, and V. Beaumont, I. Munoz-Sanjuan, M.S. Levine, C. Cepeda, E. Shigetomi, and all current and past members of the Khakh and Sofroniew laboratories for discussions. This work was supported by the CHDI Foundation (B.S.K., M.V.S., X.T., Y.A.) and the US National Institutes of Health (NS060677, MH104069 to B.S.K.).
Author information
Authors and Affiliations
Contributions
X.T., J.X., M.D.H. and B.S.K. carried out the molecular biology, imaging and electrophysiology experiments. Y.A., M.A.A. and M.V.S. performed immunohistochemistry, western blot and behavior experiments. G.C.F. and I.M. carried out the potassium concentration measurements. S.E.N. and M.L.O. performed the qPCR experiments. B.S.K. and M.V.S. directed the work. B.S.K. wrote the first draft of the paper. All of the authors contributed to the final version of the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Integrated supplementary information
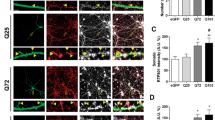
Supplementary Figure 1 Basic characterization of R6/2 mice.
a-b. Nissl (cresyl violet) stained brain sections from WT and R6/2 mice at P30 (a) and P60 (b). Note, at P60, when R6/2 mice display HD-like symptoms, the volume of the caudate putamen (CP) is reduced leading to enlarged ventricles (V). c. Quantification of the reduced CP area in R6/2 mice relative to WT and P60. d. R6/2 mice weighed less than their WT littermates at P60. Significance was assessed with an unpaired Student's t test; p values are shown.
Supplementary Figure 2 Ki67 IHC in WT and R6/2 mice.
Ki67 IHC in WT and R6/2 mice at P110 shows that many proliferating cells were seen in the subventricular zone (arrow heads), but few were seen in the striatum (arrows). In this regard, there were no differences between WT and R6/2 mice at P60-70 or P100. Some, but not all, Ki67 positive cells in the striatum colocalized with S100β, indicating that we were able to detect proliferating reactive astrocytes but that almost none were present.
Supplementary Figure 3 Western blot analysis of protein expression in striatal tissue at P104-110 for WT and R6/2 mice.
Statistical significance was assessed with an unpaired Student's t test. n indicates the number of animals from each group. P values are shown.
Supplementary Figure 4 Labeling of striatal astrocytes with four commonly used astrocyte markers.
GFAP (a), Aldh1L1(b), Glutamine synthetase (GS) (c) and S100β (d) in WT mouse at P60. As shown in a, only a few astrocytes were labeled with GFAP antibodies compared with the other three astrocyte markers. The images in the lower panels show higher magnification views from the boxes marked in the upper panels in each group.
Supplementary Figure 5 Colocalization of astrocyte markers in striatum of R6/2 mice at P60.
(a) Shows glutamine synthetase (GS; green) colocalized with Aldh1 (red) in an astrocyte shown with an arrow. (b) As in (a), but for Aldh1L1 (green) colocalized with S100β (red).
Supplementary Figure 6 Basic properties of medium spiny neurons and astrocytes.
Medium spiny neurons (MSN) and astrocytes in the striatum were easily distinguished during whole-cell recordings with Alexa 488 fluorescent dye in the pipette solution. (a) Shows a typical dendritic morphology of a MSN dialyzed with Alexa 488 (50 μM) for 30 min and the electrophysiological traces below show a negative resting membrane potential (-81 mV) and action potential firing during small depolarizing current injections. (b) Shows an astrocyte identically dialyzed with Alexa 488: a few short processes can be seen with a generally bushy appearance. The electrophysiological traces below show a negative resting membrane potential (-80 mV), but no action potential firing even for large depolarizing current injections.
Supplementary Figure 7 Barium sensitive currents.
Representative traces showing total macroscopic currents (i), Ba2+- insensitive currents (in 100 μM Ba2+ in the bath; ii) and Ba2+-sensitive currents (i-ii) recorded by whole-cell voltage clamp (from -120 mV to 40 mV) for striatal astrocytes from WT (a) and R6/2 mice (b) at P60. c. Note on average that the Ba2+-sensitive current is smaller in R6/2 astrocytes than that in WT astrocytes at P60-80.
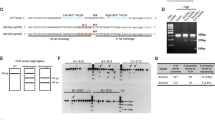
Supplementary Figure 8 qPCR analyses.
a. Kir4.1 (KCNJ10) qPCR data for R6/2 and WT mice at P60-80 relative to three control mRNAs (GAPDH, ATP5B, and UBC). There were no significant differences when the data were assessed using un paired Student's t tests. P values are shown. b. qPCR data for Q175 mice aged 10 months for Kir4.1, GFAP and GLT-1. Only mRNA levels for GLT-1 were significantly different to WT controls. Statistical significance was assessed using an unpaired Student's t test; P values are shown. The experiments were done on 4-5 mice. c. Kir4.1 (KCNJ10) qPCR data for Q175 and WT mice at 10 months age relative to three control mRNAs (GAPDH, ATP5B, and UBC). There were no significant differences when the data were assessed using un paired Student's t tests. P values are shown.
Supplementary Figure 9 Characterization of Q175 mice.
Representative images show there no CP volume change in wild type mice and heterozygous Q175 mice by Nissl staining at 6-7 months. Bar graph in right summaries the striatal area between two groups. (B) Representative images show decrease of the striatal volume in heterozygous Q175 mice at 12 months compared with comparative wild type mice. Bar graph in right shows the significant decrease of striatal areas in heterozygous Q175 mice. (C) Summary of western blots of Kir4.1, glutamine synthetase (GS), GLT-1 and GFAP protein expression levels in striatums of wild type, heterozygous and homozygous Q175 mice at 6-7 months and 9-12 months. Statistical tests were done using un paired Student's t tests; p values are shown. n indicates the number of animals in each group.
Supplementary Figure 10 Representative images for IHC using two antibodies against mHTT (mEM48 and MW8).
Both labeled nuclear inclusions in S100β labeled astrocytes to an equivalent degree. Neither is able to label cytoplasmic mHTT.
Supplementary Figure 11 Representative images show S100β immunostaining in striatal tissue from R6/2 mice injected with AAV2/5 gfaABC1D Kir4.1-GFP virus (a) or AAV2/5 gfaABC1D tdTomato virus (c) at P56.
Slice sections were collected for immunohistochemistry two weeks after viral injections at P56. Panels (a) and (c) show colocalization of S100β labeled astrocytes with Kir4.1-GFP and tdTomato positive cells, respectively. Controls from mice that received no virus injections (b and d) showed no colocalisation, but still showed S100b staining.
Supplementary Figure 12 Controls for AAV2/5 Kir4.1-GFP.
a. Astrocyte current-voltage (IV) relations. AAV2/5 Kir4.1-GFP expression in R6/2 mice increases the slope of the astrocyte IV relations and restores them to WT levels. However, Kir4.1-GFP expression in WT astrocytes did not boost the IV relations higher than WT levels. b. Ba2+ sensitive currents under the conditions shown. c-e. Western blot analysis from R6/2 mice injected with AAV2/5 for Kir4.1-GFP or AAV2/5 tdTomato. The Western blot data are for GFP (c) total Kir4.1 (d) and Glt-1 (e). Statistical differences were assessed using un paired Student's t tests. P values are shown.
Supplementary Figure 13 Assessment of inflammation.
a. Representative images for Iba1 and CD68 IHC under the conditions shown. b-c. Summary data for Iba1 and CD68 IHC for experiments such as those shown in a. No qualitative or quantitative differences are detectable in staining intensity for inflammatory markers when comparing mice that received AAV2/5Kir4.1-GFP or AAV2/5-tdTomato.
Supplementary Figure 14 Representative trace and average data for the astrocyte membrane potential in different concentrations of bath K+.
The slope of the line shows a 55 mV depolarization for a 10-fold change in K+ concentration. Additional Note: A salient feature across mouse models of HD is that MSNs display depolarized membrane potentials by as much as ∼12 mV. A vaguely formulated idea suggests that reducing astrocyte Kir4.1 function with Ba2+ in WT slices may reveal secondary consequences for MSNs, i.e., this approach will elevate K+ near neurons, depolarize MSNs and thus reveal astrocyte Kir4.1 contributions to altered MSN properties frequently observed in mouse models of HD. However, this view has problems. First, although Ba2+ is a useful tool to isolate astrocyte Kir4.1 currents, it also blocks MSN K+ channels, meaning that it will be impossible to tell if any measured effects are due to Ba2+ actions on astrocytes or MSNs directly. Second, the suggested experiment can only work if brain slices can buffer K+ locally, i.e. that local K+ is not set by the bath K+ concentration. We tested for this by measuring the astrocyte membrane potential in bath solutions of different K+ concentration, exploiting the fact that astrocytes function as K+ microelectrodes because of their high K+ conductance. The Nernst equation predicts that the membrane potential of an astrocyte with a high K+ conductance should change by ∼56 mV for a ten-fold change in bath K+ concentration. If brain slices can in fact buffer K+ in the vicinity of a cell, then we would predict a deviation from this, because the local K+ concentration would be set by buffering and not by the bath K+ concentration. We found that striatal astrocytes showed a near perfect Nernst-like ∼55 mV depolarization for a 10-fold change in bath K+ concentration. These data show that isolated preparations such as brain slices cannot buffer K+. Thus, the idea of blocking astrocyte Kir4.1 channels with Ba2+ and looking for downstream effects on MSNs because of disrupted local K+ buffering is flawed.
Supplementary Figure 15 Average data for behavioral analysis.
R6/2 mice injected with AAV2/5 for Kir4.1-GFP were not different to R6/2 mice or to R6/2 mice injected with AAV2/5 for tdTomato in assays that measured motor function (forelimb clasp time and the time to fall off a rotarod). The clasp time experiments were conducted over 1 min. Since the WT mice did not clasp in this time period, their data values are all at 60 s.
Supplementary information
Supplementary Text and Figures
Supplementary Figures 1–15 and Supplementary Tables 1–4 (PDF 18166 kb)
Rights and permissions
About this article
Cite this article
Tong, X., Ao, Y., Faas, G. et al. Astrocyte Kir4.1 ion channel deficits contribute to neuronal dysfunction in Huntington's disease model mice. Nat Neurosci 17, 694–703 (2014). https://doi.org/10.1038/nn.3691
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nn.3691
This article is cited by
-
In vivo identification of astrocyte and neuron subproteomes by proximity-dependent biotinylation
Nature Protocols (2024)
-
Microglia and complement mediate early corticostriatal synapse loss and cognitive dysfunction in Huntington’s disease
Nature Medicine (2023)
-
The translational potential of cholesterol-based therapies for neurological disease
Nature Reviews Neurology (2023)
-
Astrocytes in human central nervous system diseases: a frontier for new therapies
Signal Transduction and Targeted Therapy (2023)
-
Modeling disrupted synapse formation in wolfram syndrome using hESCs-derived neural cells and cerebral organoids identifies Riluzole as a therapeutic molecule
Molecular Psychiatry (2023)
