
Abstract
A properly mounted immune response is indispensable for recognizing and eliminating danger arising from foreign invaders and tissue trauma. However, the 'inflammatory fire' kindled by the host response must be tightly controlled to prevent it from spreading and causing irreparable damage. Accordingly, acute inflammation is self-limiting and is normally attenuated after elimination of noxious stimuli, restoration of homeostasis and initiation of tissue repair. However, unresolved inflammation may lead to the development of chronic autoimmune and degenerative diseases and cancer. Here, we discuss the key molecular mechanisms that contribute to the self-limiting nature of inflammatory signaling, with emphasis on the negative regulation of the NF-κB pathway and the NLRP3 inflammasome. Understanding these negative regulatory mechanisms should facilitate the development of much-needed therapeutic strategies for treatment of inflammatory and autoimmune pathologies.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout



Similar content being viewed by others
References
Sugimoto, M.A., Sousa, L.P., Pinho, V., Perretti, M. & Teixeira, M.M. Resolution of inflammation: what controls its onset? Front. Immunol. 7, 160 (2016).
Medzhitov, R. Origin and physiological roles of inflammation. Nature 454, 428–435 (2008).
Dinarello, C.A., Simon, A. & van der Meer, J.W. Treating inflammation by blocking interleukin-1 in a broad spectrum of diseases. Nat. Rev. Drug Discov. 11, 633–652 (2012).
Headland, S.E. & Norling, L.V. The resolution of inflammation: principles and challenges. Semin. Immunol. 27, 149–160 (2015).
Tang, D., Kang, R., Coyne, C.B., Zeh, H.J. & Lotze, M.T. PAMPs and DAMPs: signal 0s that spur autophagy and immunity. Immunol. Rev. 249, 158–175 (2012).
Arthur, J.S. & Ley, S.C. Mitogen-activated protein kinases in innate immunity. Nat. Rev. Immunol. 13, 679–692 (2013).
O'Shea, J.J. et al. The JAK-STAT pathway: impact on human disease and therapeutic intervention. Annu. Rev. Med. 66, 311–328 (2015).
Zhang, Q., Lenardo, M.J. & Baltimore, D. 30 years of NF-κB: a blossoming of relevance to human pathobiology. Cell 168, 37–57 (2017).
Ben-Neriah, Y. & Karin, M. Inflammation meets cancer, with NF-κB as the matchmaker. Nat. Immunol. 12, 715–723 (2011).
Peng, B. et al. Defective feedback regulation of NF-κB underlies Sjogren's syndrome in mice with mutated κB enhancers of the IκBα promoter. Proc. Natl. Acad. Sci. USA 107, 15193–15198 (2010).
Alves, B.N. et al. IκBɛ is a key regulator of B cell expansion by providing negative feedback on cRel and RelA in a stimulus-specific manner. J. Immunol. 192, 3121–3132 (2014).
Kobayashi, K. et al. IRAK-M is a negative regulator of Toll-like receptor signaling. Cell 110, 191–202 (2002).
Zhou, H. et al. IRAK-M mediates Toll-like receptor/IL-1R-induced NFκB activation and cytokine production. EMBO J. 32, 583–596 (2013). This paper describes how IRAK-M mediates a MEKK3-dependent second wave of NF-κB activation, which results in the expression of negative regulators of this pathway, such as IκBα or A20.
Miyata, M. et al. Glucocorticoids suppress inflammation via the upregulation of negative regulator IRAK-M. Nat. Commun. 6, 6062 (2015).
Burns, K. et al. Inhibition of interleukin 1 receptor/Toll-like receptor signaling through the alternatively spliced, short form of MyD88 is due to its failure to recruit IRAK-4. J. Exp. Med. 197, 263–268 (2003).
Hardy, M.P. & O'Neill, L.A. The murine IRAK2 gene encodes four alternatively spliced isoforms, two of which are inhibitory. J. Biol. Chem. 279, 27699–27708 (2004).
Koop, A. et al. Novel splice variants of human IKKɛ negatively regulate IKKɛ-induced IRF3 and NF-kB activation. Eur. J. Immunol. 41, 224–234 (2011).
Taganov, K.D., Boldin, M.P., Chang, K.J. & Baltimore, D. NF-kappaB-dependent induction of microRNA miR-146, an inhibitor targeted to signaling proteins of innate immune responses. Proc. Natl. Acad. Sci. USA 103, 12481–12486 (2006).
Boldin, M.P. et al. miR-146a is a significant brake on autoimmunity, myeloproliferation, and cancer in mice. J. Exp. Med. 208, 1189–1201 (2011). This paper demonstrates that the targeted deletion of miR-146a in mice increases susceptibility to endotoxic shock and results in the development of an autoimmune disorder that is at least partially due to the derepression of the miR-146a-target genes TRAF6 and IRAK1.
Zhao, J.L. et al. NF-kappaB dysregulation in microRNA-146a-deficient mice drives the development of myeloid malignancies. Proc. Natl. Acad. Sci. USA 108, 9184–9189 (2011).
Zhou, X. et al. MicroRNA-302b augments host defense to bacteria by regulating inflammatory responses via feedback to TLR/IRAK4 circuits. Nat. Commun. 5, 3619 (2014).
O'Connell, R.M., Rao, D.S. & Baltimore, D. microRNA regulation of inflammatory responses. Annu. Rev. Immunol. 30, 295–312 (2012).
Mehta, A. & Baltimore, D. MicroRNAs as regulatory elements in immune system logic. Nat. Rev. Immunol. 16, 279–294 (2016).
Carballo, E., Lai, W.S. & Blackshear, P.J. Feedback inhibition of macrophage tumor necrosis factor-alpha production by tristetraprolin. Science 281, 1001–1005 (1998).
Molle, C. et al. Tristetraprolin regulation of interleukin 23 mRNA stability prevents a spontaneous inflammatory disease. J. Exp. Med. 210, 1675–1684 (2013).
Taylor, G.A. et al. A pathogenetic role for TNF alpha in the syndrome of cachexia, arthritis, and autoimmunity resulting from tristetraprolin (TTP) deficiency. Immunity 4, 445–454 (1996).
Mino, T. et al. Regnase-1 and Roquin regulate a common element in inflammatory mRNAs by spatiotemporally distinct mechanisms. Cell 161, 1058–1073 (2015).
Pratama, A. et al. Roquin-2 shares functions with its paralog Roquin-1 in the repression of mRNAs controlling T follicular helper cells and systemic inflammation. Immunity 38, 669–680 (2013).
Matsushita, K. et al. Zc3h12a is an RNase essential for controlling immune responses by regulating mRNA decay. Nature 458, 1185–1190 (2009).
Miao, R. et al. Targeted disruption of MCPIP1/Zc3h12a results in fatal inflammatory disease. Immunol. Cell Biol. 91, 368–376 (2013).
Garg, A.V. et al. MCPIP1 endoribonuclease activity negatively regulates interleukin-17-mediated signaling and inflammation. Immunity 43, 475–487 (2015).
Jeltsch, K.M. et al. Cleavage of roquin and regnase-1 by the paracaspase MALT1 releases their cooperatively repressed targets to promote TH17 differentiation. Nat. Immunol. 15, 1079–1089 (2014). In this study, the authors demonstrate that Roquin and Regnase-1 repress differentiation of IL-17-producing helper T cells via targeting a specific set of mRNAs. This repression is released after T cell–receptor stimulation by MALT1-mediated cleavage of Roquin and Regnase-1.
Uehata, T. et al. Malt1-induced cleavage of regnase-1 in CD4+ helper T cells regulates immune activation. Cell 153, 1036–1049 (2013).
Afonina, I.S., Elton, L., Carpentier, I. & Beyaert, R. MALT1: a universal soldier: multiple strategies to ensure NF-κB activation and target gene expression. FEBS J. 282, 3286–3297 (2015).
Brechmann, M. et al. A PP4 holoenzyme balances physiological and oncogenic nuclear factor-kappa B signaling in T lymphocytes. Immunity 37, 697–708 (2012).
Shen, X.F., Zhao, Y., Jiang, J.P., Guan, W.X. & Du, J.F. Phosphatase Wip1 in immunity: an overview and update. Front. Immunol. 8, 8 (2017).
Través, P.G. et al. Pivotal role of protein tyrosine phosphatase 1B (PTP1B) in the macrophage response to pro-inflammatory and anti-inflammatory challenge. Cell Death Dis. 5, e1125 (2014).
Lawrence, T., Bebien, M., Liu, G.Y., Nizet, V. & Karin, M. IKKα limits macrophage NF-κB activation and contributes to the resolution of inflammation. Nature 434, 1138–1143 (2005).
Shembade, N., Pujari, R., Harhaj, N.S., Abbott, D.W. & Harhaj, E.W. The kinase IKKα inhibits activation of the transcription factor NF-κB by phosphorylating the regulatory molecule TAX1BP1. Nat. Immunol. 12, 834–843 (2011).
Hu, H. & Sun, S.C. Ubiquitin signaling in immune responses. Cell Res. 26, 457–483 (2016).
Komander, D. & Rape, M. The ubiquitin code. Annu. Rev. Biochem. 81, 203–229 (2012).
Abdul-Sater, A.A. et al. The signaling adaptor TRAF1 negatively regulates Toll-like receptor signaling and this underlies its role in rheumatic disease. Nat. Immunol. 18, 26–35 (2017).
Seymour, R.E. et al. Spontaneous mutations in the mouse Sharpin gene result in multiorgan inflammation, immune system dysregulation and dermatitis. Genes Immun. 8, 416–421 (2007).
Gerlach, B. et al. Linear ubiquitination prevents inflammation and regulates immune signalling. Nature 471, 591–596 (2011).
Teh, C.E. et al. Linear ubiquitin chain assembly complex coordinates late thymic T-cell differentiation and regulatory T-cell homeostasis. Nat. Commun. 7, 13353 (2016).
Fiil, B.K. et al. OTULIN restricts Met1-linked ubiquitination to control innate immune signaling. Mol. Cell 50, 818–830 (2013).
Keusekotten, K. et al. OTULIN antagonizes LUBAC signaling by specifically hydrolyzing Met1-linked polyubiquitin. Cell 153, 1312–1326 (2013). This paper, along with refs. 46 and 48 , provides evidence that OTULIN is a unique deubiquitinase that hydrolyzes M1-linked ubiquitin chains and negatively regulates NF-κB signaling.
Rivkin, E. et al. The linear ubiquitin-specific deubiquitinase gumby regulates angiogenesis. Nature 498, 318–324 (2013).
Elliott, P.R. et al. Molecular basis and regulation of OTULIN-LUBAC interaction. Mol. Cell 54, 335–348 (2014).
Damgaard, R.B. et al. The deubiquitinase OTULIN is an essential negative regulator of inflammation and autoimmunity. Cell 166, 1215–1230.e1220 (2016). This article shows that OTULIN is essential in preventing TNF-dependent systemic inflammation in mice and humans.
Zhou, Q. et al. Biallelic hypomorphic mutations in a linear deubiquitinase define otulipenia, an early-onset autoinflammatory disease. Proc. Natl. Acad. Sci. USA 113, 10127–10132 (2016).
Brummelkamp, T.R., Nijman, S.M., Dirac, A.M. & Bernards, R. Loss of the cylindromatosis tumour suppressor inhibits apoptosis by activating NF-κB. Nature 424, 797–801 (2003).
Kovalenko, A. et al. The tumour suppressor CYLD negatively regulates NF-κB signalling by deubiquitination. Nature 424, 801–805 (2003).
Trompouki, E. et al. CYLD is a deubiquitinating enzyme that negatively regulates NF-κB activation by TNFR family members. Nature 424, 793–796 (2003).
Elliott, P.R. et al. SPATA2 links CYLD to LUBAC, activates CYLD, and controls LUBAC signaling. Mol. Cell 63, 990–1005 (2016).
Kupka, S. et al. SPATA2-mediated binding of CYLD to HOIP enables CYLD recruitment to signaling complexes. Cell Rep. 16, 2271–2280 (2016).
Wagner, S.A., Satpathy, S., Beli, P. & Choudhary, C. SPATA2 links CYLD to the TNF-α receptor signaling complex and modulates the receptor signaling outcomes. EMBO J. 35, 1868–1884 (2016).
Schlicher, L. et al. SPATA2 promotes CYLD activity and regulates TNF-induced NF-κB signaling and cell death. EMBO Rep. 17, 1485–1497 (2016).
Lork, M., Verhelst, K. & Beyaert, R. CYLD, A20 and OTULIN deubiquitinases in NF-κB signaling and cell death: so similar, yet so different. Cell Death Differ. http://dx.doi.org/10.1038/cdd.2017.46 (2017).
Dixit, V.M. et al. Tumor necrosis factor-α induction of novel gene products in human endothelial cells including a macrophage-specific chemotaxin. J. Biol. Chem. 265, 2973–2978 (1990).
Wertz, I.E. et al. Phosphorylation and linear ubiquitin direct A20 inhibition of inflammation. Nature 528, 370–375 (2015).
Ma, A. & Malynn, B.A. A20: linking a complex regulator of ubiquitylation to immunity and human disease. Nat. Rev. Immunol. 12, 774–785 (2012).
Zhou, Q. et al. Loss-of-function mutations in TNFAIP3 leading to A20 haploinsufficiency cause an early-onset autoinflammatory disease. Nat. Genet. 48, 67–73 (2016). This study shows that A20 haploinsufficiency leads to the development of early-onset autoinflammatory disease in humans, which correlates with increased NF-κB activation and sustained levels of ubiquitinated A20 substrates.
Lee, E.G. et al. Failure to regulate TNF-induced NF-κB and cell death responses in A20-deficient mice. Science 289, 2350–2354 (2000).
Catrysse, L., Vereecke, L., Beyaert, R. & van Loo, G. A20 in inflammation and autoimmunity. Trends Immunol. 35, 22–31 (2014).
Schuijs, M.J. et al. Farm dust and endotoxin protect against allergy through A20 induction in lung epithelial cells. Science 349, 1106–1110 (2015). This article identifies single-nucleotide polymorphisms in the A20-encoding gene as a risk factor for allergy and asthma development in children growing up on farms. The authors demonstrate that A20 mediates the protective effects of chronic low-dose endotoxin or farm-dust exposure in a mouse model of house-dust-mite-induced asthma.
Newton, K. et al. RIPK3 deficiency or catalytically inactive RIPK1 provides greater benefit than MLKL deficiency in mouse models of inflammation and tissue injury. Cell Death Differ. 23, 1565–1576 (2016).
Onizawa, M. et al. The ubiquitin-modifying enzyme A20 restricts ubiquitination of the kinase RIPK3 and protects cells from necroptosis. Nat. Immunol. 16, 618–627 (2015).
Maelfait, J. et al. A20 deficiency in lung epithelial cells protects against influenza A virus infection. PLoS Pathog. 12, e1005410 (2016).
Matmati, M. et al. A20 (TNFAIP3) deficiency in myeloid cells triggers erosive polyarthritis resembling rheumatoid arthritis. Nat. Genet. 43, 908–912 (2011).
Vande Walle, L. et al. Negative regulation of the NLRP3 inflammasome by A20 protects against arthritis. Nature 512, 69–73 (2014).
Vereecke, L. et al. A20 controls intestinal homeostasis through cell-specific activities. Nat. Commun. 5, 5103 (2014).
Lin, A. & Karin, M. NF-kκB in cancer: a marked target. Semin. Cancer Biol. 13, 107–114 (2003).
Murphy, S.F. et al. Intestinal epithelial expression of TNFAIP3 results in microbial invasion of the inner mucus layer and induces colitis in IL-10-deficient mice. Am. J. Physiol. Gastrointest. Liver Physiol. 307, G871–G882 (2014).
Wertz, I.E. et al. De-ubiquitination and ubiquitin ligase domains of A20 downregulate NF-kappaB signalling. Nature 430, 694–699 (2004).
Lu, T.T. et al. Dimerization and ubiquitin mediated recruitment of A20, a complex deubiquitinating enzyme. Immunity 38, 896–905 (2013).
De, A., Dainichi, T., Rathinam, C.V. & Ghosh, S. The deubiquitinase activity of A20 is dispensable for NF-κB signaling. EMBO Rep. 15, 775–783 (2014). The authors show that knock-in mice expressing mutant A20 that lacks deubiquitinase activity do not show an inflammatory phenotype and respond normally to LPS and TNF, thus challenging the prevailing view that the deubiquitinase activity of A20 is essential for its anti-inflammatory function.
Verhelst, K. et al. A20 inhibits LUBAC-mediated NF-κB activation by binding linear polyubiquitin chains via its zinc finger 7. EMBO J. 31, 3845–3855 (2012).
Zilberman-Rudenko, J. et al. Recruitment of A20 by the C-terminal domain of NEMO suppresses NF-κB activation and autoinflammatory disease. Proc. Natl. Acad. Sci. USA 113, 1612–1617 (2016).
Song, L. et al. miR-486 sustains NF-κB activity by disrupting multiple NF-κB-negative feedback loops. Cell Res. 23, 274–289 (2013).
Gantier, M.P. et al. A miR-19 regulon that controls NF-κB signaling. Nucleic Acids Res. 40, 8048–8058 (2012).
Coornaert, B. et al. T cell antigen receptor stimulation induces MALT1 paracaspase-mediated cleavage of the NF-κB inhibitor A20. Nat. Immunol. 9, 263–271 (2008). This is the first report identifying MALT1 as a protease and describing A20 as a substrate.
Staal, J. et al. T-cell receptor-induced JNK activation requires proteolytic inactivation of CYLD by MALT1. EMBO J. 30, 1742–1752 (2011).
Afonina, I.S. et al. The paracaspase MALT1 mediates CARD14-induced signaling in keratinocytes. EMBO Rep. 17, 914–927 (2016).
Klei, L.R. et al. MALT1 protease activation triggers acute disruption of endothelial barrier integrity via CYLD cleavage. Cell Rep. 17, 221–232 (2016).
Demeyer, A., Staal, J. & Beyaert, R. Targeting MALT1 proteolytic activity in immunity, inflammation and disease: good or bad? Trends Mol. Med. 22, 135–150 (2016).
Kathania, M. et al. Itch inhibits IL-17-mediated colon inflammation and tumorigenesis by ROR-γt ubiquitination. Nat. Immunol. 17, 997–1004 (2016).
Jin, J. et al. Epigenetic regulation of the expression of Il12 and Il23 and autoimmune inflammation by the deubiquitinase Trabid. Nat. Immunol. 17, 259–268 (2016).
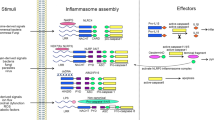
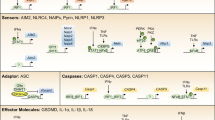
Gross, O., Thomas, C.J., Guarda, G. & Tschopp, J. The inflammasome: an integrated view. Immunol. Rev. 243, 136–151 (2011).
Zhong, Z., Sanchez-Lopez, E. & Karin, M. Autophagy, NLRP3 inflammasome and auto-inflammatory/immune diseases. Clin. Exp. Rheumatol. 34 (Suppl. 98), 12–16 (2016).
Martinon, F., Mayor, A. & Tschopp, J. The inflammasomes: guardians of the body. Annu. Rev. Immunol. 27, 229–265 (2009).
Karin, M. & Clevers, H. Reparative inflammation takes charge of tissue regeneration. Nature 529, 307–315 (2016).
Zhong, Z., Sanchez-Lopez, E. & Karin, M. Autophagy, inflammation, and immunity: a troika governing cancer and its treatment. Cell 166, 288–298 (2016).
Brydges, S.D. et al. Divergence of IL-1, IL-18, and cell death in NLRP3 inflammasomopathies. J. Clin. Invest. 123, 4695–4705 (2013).
Guo, H., Callaway, J.B. & Ting, J.P. Inflammasomes: mechanism of action, role in disease, and therapeutics. Nat. Med. 21, 677–687 (2015).
Elliott, E.I. & Sutterwala, F.S. Initiation and perpetuation of NLRP3 inflammasome activation and assembly. Immunol. Rev. 265, 35–52 (2015).
Zhou, R., Yazdi, A.S., Menu, P. & Tschopp, J. A role for mitochondria in NLRP3 inflammasome activation. Nature 469, 221–225 (2011). This study provides the first evidence of the critical role of mitochondria in NLRP3-inflammasome activation.
Zhong, Z. et al. TRPM2 links oxidative stress to NLRP3 inflammasome activation. Nat. Commun. 4, 1611 (2013).
Shimada, K. et al. Oxidized mitochondrial DNA activates the NLRP3 inflammasome during apoptosis. Immunity 36, 401–414 (2012). This is the first report showing that oxidized mitochondrial DNA activates the NLRP3 inflammasome.
Nakahira, K. et al. Autophagy proteins regulate innate immune responses by inhibiting the release of mitochondrial DNA mediated by the NALP3 inflammasome. Nat. Immunol. 12, 222–230 (2011). This study shows that autophagy inhibits the NLRP3 inflammasome through regulating the abundance of mitochondrial DNA.
Iyer, S.S. et al. Mitochondrial cardiolipin is required for Nlrp3 inflammasome activation. Immunity 39, 311–323 (2013). This study shows that the mitochondrial lipid cardiolipin is required for optimal NLRP3-inflammasome activation.
Liu, T. et al. Single-cell imaging of caspase-1 dynamics reveals an all-or-none inflammasome signaling response. Cell Rep. 8, 974–982 (2014).
Saitoh, T. et al. Loss of the autophagy protein Atg16L1 enhances endotoxin-induced IL-1β production. Nature 456, 264–268 (2008).
Zhong, Z. et al. NF-κB restricts inflammasome activation via elimination of damaged mitochondria. Cell 164, 896–910 (2016). This study identifies the NF-κB–p62–mitophagy pathway as the key macrophage-intrinsic negative regulatory mechanism that keeps NLRP3 inflammasome activity in check.
Greten, F.R. et al. NF-k(B is a negative regulator of IL-1 (secretion as revealed by genetic and pharmacological inhibition of IKKβ. Cell 130, 918–931 (2007). This study provides the first evidence that inhibition of NF-κB exacerbates NLRP3-inflammasome-dependent inflammation.
Umemura, A. et al. p62, upregulated during preneoplasia, induces hepatocellular carcinogenesis by maintaining survival of stressed HCC-initiating cells. Cancer Cell 29, 935–948 (2016).
Moscat, J., Karin, M. & Diaz-Meco, M.T. p62 in cancer: signaling adaptor beyond autophagy. Cell 167, 606–609 (2016).
Guarda, G. et al. T cells dampen innate immune responses through inhibition of NLRP1 and NLRP3 inflammasomes. Nature 460, 269–273 (2009).
Guarda, G. et al. Type I interferon inhibits interleukin-1 production and inflammasome activation. Immunity 34, 213–223 (2011).
Reboldi, A. et al. 25-Hydroxycholesterol suppresses interleukin-1-driven inflammation downstream of type I interferon. Science 345, 679–684 (2014).
Mishra, B.B. et al. Nitric oxide controls the immunopathology of tuberculosis by inhibiting NLRP3 inflammasome-dependent processing of IL-1β. Nat. Immunol. 14, 52–60 (2013).
Bauernfeind, F. et al. NLRP3 inflammasome activity is negatively controlled by miR-223. J. Immunol. 189, 4175–4181 (2012).
Netea, M.G., van de Veerdonk, F.L., van der Meer, J.W., Dinarello, C.A. & Joosten, L.A. Inflammasome-independent regulation of IL-1-family cytokines. Annu. Rev. Immunol. 33, 49–77 (2015).
Yan, Y. et al. Dopamine controls systemic inflammation through inhibition of NLRP3 inflammasome. Cell 160, 62–73 (2015).
Malcomson, B. et al. Connectivity mapping (ssCMap) to predict A20-inducing drugs and their antiinflammatory action in cystic fibrosis. Proc. Natl. Acad. Sci. USA 113, E3725–E3734 (2016).
Acknowledgements
Work in the laboratory of R.B. is supported by grants from the VIB, the Fund for Scientific Research Flanders (FWO), the Foundation Against Cancer and Ghent University (Concerted Research Actions, GOA). I.S.A. is supported by an FWO postdoctoral fellowship and an FWO research grant. Z.Z. was supported by a Cancer Research Irvington Postdoctoral Fellowship, a Prevent Cancer Foundation Board of Directors Award and an American Association for the Study of Liver Diseases (AASLD) Pinnacle Research Award. Research was supported by grants from the NIH (AI043477 and CA163798) to M.K., the Leukemia and Lymphoma Society SCOR (20132569) to M.K. and the Alliance for Lupus Research (257214) to M.K., who is supported as an American Cancer Research Professor and as the Ben and Wanda Hildyard Chair for Mitochondrial and Metabolic Diseases.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
About this article

Cite this article
Afonina, I., Zhong, Z., Karin, M. et al. Limiting inflammation—the negative regulation of NF-κB and the NLRP3 inflammasome. Nat Immunol 18, 861–869 (2017). https://doi.org/10.1038/ni.3772
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ni.3772
This article is cited by
-
Microglia TREM1-mediated neuroinflammation contributes to central sensitization via the NF-κB pathway in a chronic migraine model
The Journal of Headache and Pain (2024)
-
NF-κB in biology and targeted therapy: new insights and translational implications
Signal Transduction and Targeted Therapy (2024)
-
Administration of USP7 inhibitor p22077 alleviates Angiotensin II (Ang II)-induced atrial fibrillation in Mice
Hypertension Research (2024)
-
The Role and Potential Mechanisms of Rehabilitation Exercise Improving Cardiac Remodeling
Journal of Cardiovascular Translational Research (2024)
-
Dietary Zinc Ameliorates TNBS-Induced Colitis in Mice Associated with Regulation of Th1/Th2/Th17 Balance and NF-κB/NLRP3 Signaling Pathway
Biological Trace Element Research (2024)
