
Abstract
Hepatic natural killer (NK) cells mediate antigen-specific contact hypersensitivity (CHS) in mice deficient in T cells and B cells. We report here that hepatic NK cells, but not splenic or naive NK cells, also developed specific memory of vaccines containing antigens from influenza, vesicular stomatitis virus (VSV) or human immunodeficiency virus type 1 (HIV-1). Adoptive transfer of virus-sensitized NK cells into naive recipient mice enhanced the survival of the mice after lethal challenge with the sensitizing virus but not after lethal challenge with a different virus. NK cell memory of haptens and viruses depended on CXCR6, a chemokine receptor on hepatic NK cells that was required for the persistence of memory NK cells but not for antigen recognition. Thus, hepatic NK cells can develop adaptive immunity to structurally diverse antigens, an activity that requires NK cell–expressed CXCR6.
Similar content being viewed by others
Main
The textbook view of adaptive immunity is that it is mediated exclusively by T cells and B cells, which rely on nonhomologous recombination of variable, diversity and joining gene segments mediated by the RAG-1–RAG-2 recombinase to generate a diverse repertoire of T cell and B cell antigen receptors1. The activation of such receptors by cognate antigen triggers clonal selection and differentiation into short-lived effector cells and long-lived memory cells that, after antigen rechallenge, mount accelerated and enhanced recall responses. Classic examples of such antigen-specific memory, the hallmark of adaptive immunity, include delayed-type hypersensitivity (DTH)2,3 and hapten-induced contact hypersensitivity (CHS)4,5.
Like T cells and B cells, natural killer (NK) cells mediate resistance to tumors and infections by a variety of mechanisms that are still incompletely understood6. However, NK cells cannot express RAG-dependent receptors and instead use germline-encoded receptors that integrate activating and dampening signals7. Notable examples of mouse NK cell receptors that detect specific pathogen–associated structures include NKp46 and Ly49H, which recognize influenza A–encoded hemagglutinin (HA)8,9 and the murine cytomegalovirus (MCMV)-encoded m157 protein10, respectively. Mouse NK cells can express just one Ly49 receptor or random combinations of several Ly49 receptors, some of which recognize host major histocompatibility complex (MHC) class I (refs. 11,12). Ly49H is a member of this family and confers NK cell–mediated resistance to MCMV infection in C57BL/6 mice. Notably, Ly49H+ NK cells develop features of adaptive immunity after infection with this virus13. However, as Ly49H is found only in the C57BL/6 strain14, it has been unclear whether NK cells can develop virus-specific memory in other genetic backgrounds or in response to viruses other than MCMV. This question may also be clinically relevant, as certain NK cell subsets have been linked to the ability of rare people infected with human immunodeficiency virus type 1 (HIV-1) to exert long-term control over HIV-1 replication15,16.
The idea that NK cells can mediate adaptive immunity was initially suggested by observations that mouse strains that lack T cells and B cells develop vigorous CHS responses to various distinct haptens17,18. These responses have the hallmarks of adaptive immunity: they are sensitization dependent, persist for at least 4 weeks and are elicited only by haptens to which mice have previously been sensitized17. Several lines of evidence have established that NK cells are necessary and sufficient for this activity: CHS responses in mice deficient in recombination-activating gene 2 (Rag2−/− mice) are abolished after depletion of NK cells and are absent from Rag2−/− mice also deficient in the gene encoding the γ-chain of the interleukin 2 (IL-2) receptor (Il2rg−/− mice), which lack all lymphocytes, including NK cells19. Moreover, after sensitization of donor mice with 2,4-dinitro-1-fluorobenzene (DNFB), adoptive transfer of NK cells from donor livers confers vigorous CHS to naive Rag2−/−Il2rg−/− recipient mice but adoptive transfer of NK cells from spleens does not. Among hepatic NK cells, DNFB-specific memory is concentrated in a subset expressing Thy-1 and Ly49C/I17. However, those and all other NK receptors described so far are similarly represented among hapten-reactive (hepatic) and nonreactive (nonhepatic) NK cells17,20. Thus, although classic NK receptors contribute to the function of NK cells, it is unclear whether and to what extent they are involved in the unique adaptive properties of hapten-specific hepatic NK cells.
It is also unknown why hapten-specific memory NK cells reside preferentially in the liver. This restricted distribution is similar to that of NKT cells, which express RAG-dependent invariable T cell antigen receptors21 and whose intrahepatic survival and homeostasis requires CXCR6 (CD186), a chemokine receptor that is prominently expressed on NKT cells, a subset of activated T cells and some NK cells22,23. CXCR6 has a single ligand, CXCL16, a chemokine that is constitutively expressed on liver sinusoidal endothelium24,25. Although CXCR6-deficient mice have been described, the function of CXCR6+ NK cells has not been investigated22,26.
We undertook the present study to address several questions. First, we set out to further characterize hapten-specific memory NK cells and the role of antigen in their maintenance and trafficking. Second, we investigated whether NK cells can develop memory of viruses other than MCMV and whether this ability depends on the host's genetic background or the pathogenicity of the virus. Third, we explored the role of CXCR6 in both hapten- and virus-specific memory NK cells. Our results obtained with two different mouse strains indicate that a discrete subset of CXCR6+ NK cells acquired and retained antigen-specific memory of at least five structurally, chemically and biologically distinct exogenous antigens without requiring antigen persistence. In each case, memory NK cells were concentrated in the liver and were critically dependent on CXCR6, which was not required for antigen recognition but seemed to regulate the effector function and survival of memory NK cells.
Results
Antigen-independent persistence of memory NK cells
Published experiments with Rag2−/− mice have shown that NK cells retain memory of DNFB for up to 4 weeks after skin sensitization17. However, haptens form covalent adducts with self proteins, which may persist in the body for weeks27, so it has remained unclear if NK cell memory is maintained by residual antigen. To address this, we adoptively transferred sensitized hepatic Thy-1+ NK cells into antigen-free recipients. We either left Rag1−/− mice naive (treated them with solvent) or sensitized them with DNFB or oxazolone (OXA) and used them as donors for adoptive transfer of flow cytometry–sorted NK cells into Rag2−/−Il2rg−/− recipients; we chose to use these mice because this strain cannot produce lymphocytes, including NK cells19,28, which allows easy recovery and analysis of donor NK cells. Then, 1 d or 4 months later, we challenged recipients by painting one ear with hapten and the other with solvent and measured ear swelling after 24 h (Fig. 1a). Whereas recipients of sensitized splenic NK cells (data not shown) or naive hepatic NK cells were unresponsive, the challenged ears of DNFB- or OXA-primed hepatic NK cell recipients became markedly inflamed at both time points. A response was elicited only in mice challenged with the sensitizing hapten, not in mice challenged with the other hapten. The difference between transferred NK cell subsets in recall capacity cannot be explained by differences in survival, as all groups had similar NK cell numbers 4 months after transfer (Fig. 1b). Indeed, consistent with published reports29, the transferred NK cells expanded their populations about five- to sevenfold in the lymphopenic Rag2−/−Il2rg−/− recipients regardless of their organ of origin or immunization history.

(a) Hapten-specific CHS responses in naive Rag2−/−Il2rg−/− mice (n = 10–15 per group) that received hepatic CD45+NK1.1+Thy-1+ NK cells (1 × 105) from naive (vehicle-exposed; Acetone) or DNFB- or OXA-sensitized Rag1−/− donors (Sensitization) and were challenged 24 h or 4 months after transfer; ear swelling was calculated after 24 h by subtraction of background swelling in naive mice from that in recipients of NK cells. *P < 0.01 and **P < 0.001 (analysis of variance (ANOVA)). (b) Flow cytometry analysis of the survival and population expansion of adoptively transferred NK cells 2 weeks after the 4-month challenge in a, presented as the number of liver-resident CD45+ NK1.1+ cells. Results were similar for mice challenged with DNFB or OXA, so data were pooled according to donor sensitization. No NK1.1+ cells were detected in mock (PBS)-injected control recipient Rag2−/−Il2rg−/− mice (data not shown). (c) Hapten-specific CHS responses in naive C57BL/6 mice (n = 10–15 per group) that received sorted CD45+NK1.1+CD3−Thy-1+ NK cells (1 × 105) from donors transgenic for expression of GFP-tagged actin, then were challenged 6 weeks later with OXA (left) or DNFB (right) and analyzed as in a. *P < 0.001 and **P < 0.0001 (ANOVA). (d) Recruitment of memory NK cells to challenge sites in naive Rag2−/−Il2rg−/− mice (n = 6–7 per group) that received adoptive transfer of mixtures of hepatic CD45+NK1.1+Thy-1+ NK cells from naive CD45.1+ wild-type donors (C57BL/6) and from CD45.2+ DNFB- or OXA-sensitized wild-type or donors transgenic for expression of GFP-tagged actin (1 × 105 each) and, 1 month later, were challenged in the ears with either DNFB or OXA; livers and ears collected at 24, 48 and 72 h were analyzed by flow cytometry for NK cells whose origin was distinguished by congenic or fluorescent markers (results presented as mean of all mice at 24, 48 and 72 h). No NK cells were found in acetone-challenged control ears (data not shown). *P < 10−11 (ANOVA). Data are representative of three to five independent experiments (pooled results; error bars, s.d.).
Although the results reported above indicated that lymphopenia-induced proliferation is not unique to memory NK cells, our findings raised the possibility that NK cell proliferation was necessary to maintain memory. To address this, we sorted green fluorescent protein (GFP)-expressing NK cells from naive and sensitized RAG-sufficient, C57BL/6 donors in which GFP is expressed transgenically under control of the actin promoter and transferred the cells into wild-type mice, which we challenged 6 weeks later. Wild-type recipients of DNFB- or OXA-primed hepatic NK cells also mounted vigorous CHS responses when challenged with the sensitizing hapten but not when challenged with the nonsensitizing hapten, whereas recipients of sensitized splenic or naive NK cells remained unresponsive (Fig. 1c). All recipients had similar numbers of GFP+ NK cells 8 weeks after transfer; the numbers recovered at this time were 24–28% of the original input (Supplementary Fig. 1a), which suggested that NK cell populations did not expand. Thus, NK cells persist for several months in both lymphocyte-sufficient and lymphopenic hosts. Although the survival and population expansion of adoptively transferred NK cells were independent of prior sensitization or the source organ, only sensitized hepatic NK cells acquired transferable antigen-specific memory, and they did so regardless of the presence of other lymphocytes.
Accumulation of memory NK cells at effector sites
Although NK cells are known to require endothelial selectins and β2 integrins to access the challenged ears of DNFB-sensitized mice17, it has remained unclear whether this recruitment is antigen specific. Thus, we sorted NK cells from naive CD45.1+ donor mice and CD45.2+ donor mice, either wild-type or transgenic for actin promoter–driven GFP expression, that had been sensitized with DNFB or OXA. We made mixtures of equal numbers of each population and injected these into naive Rag2−/−Il2rg−/− recipient mice. Then, 1 month later, we challenged the ears of recipient mice with hapten and collected ears and livers at various time points to determine the frequency and genotype of tissue-resident NK cells. Although the livers of recipient mice contained equal numbers of all three donor cell subsets at every time point, infiltrating NK cells in hapten-challenged ears were derived almost exclusively from donors that had been sensitized with the hapten used for challenge (Fig. 1d and Supplementary Fig. 1b). Thus, only DNFB-sensitized NK cells were recruited to DNFB-challenged ears, whereas OXA-sensitized NK cells 'preferentially' accumulated in OXA-challenged ears. We conclude not only that memory NK cells discriminate between hapten-based antigens to mediate specific CHS responses but also that their recruitment and/or retention at sites of challenge is hapten specific.
NK cell memory of viruses
Having confirmed that hepatic memory NK cells are long lived and selectively migrate to sites of hapten challenge to mediate antigen-specific recall responses even after a prolonged antigen-free interval, we sought to determine whether NK cells also develop memory of other exogenous antigens. It has been reported that NK cells show features of adaptive immunity in MCMV-infected C57BL/6 mice13, in which ∼50% of NK cells express Ly49H, which recognizes the MCMV-encoded m157 protein. After a primary challenge with MCMV, NK cells proliferate and give rise to Ly49H-dependent protective memory. However, unlike hapten-specific memory NK cells, MCMV-restricted NK cells are not restricted to the liver13 and have not been found in other mouse strains, most of which do not express Ly49H14.
To assess more broadly whether NK cells can develop antiviral memory, we immunized Rag1−/− mice subcutaneously with either noninfectious virus-like particles (VLPs) containing both HA and matrix protein M1 from influenza strain A/PR/8/34 (PR8-VLPs)30,31 (Supplementary Fig. 2) or vesicular stomatitis virus inactivated by ultraviolet irradiation (UV-VSV)32. We then sorted splenic and hepatic naive and immunized NK cells and adoptively transferred them into naive Rag2−/−Il2rg−/− mice. We challenged the recipient mice 4 weeks later by injecting PR8-VLPs or UV-VSV into one ear and PBS into the contralateral ear. Hepatic NK cells mediated vigorous and sustained DTH responses to both influenza VLPs and VSV in an immunization-dependent and virus-specific manner, whereas recipients of sensitized splenic or naive NK cells were unresponsive (Fig. 2a).

(a) Virus-specific DTH in naive Rag2−/−Il2rg−/− mice (n = 8–10 per group) that received adoptive transfer of splenic or hepatic CD45+ NK1.1+ NK cells (8 × 104) 1 month after immunization of Rag1−/− donor mice, followed by challenge of ears of recipient mice by subcutaneous injection 2 months later and analysis after 24 h (as in Fig. 1a). P values, Student's t-test. (b) Survival of Rag2−/−Il2rg−/− mice (n = 15–19 per group) treated by the sensitization and transfer protocol in a, then infected intranasally with influenza A strain A/PR/8/34 (500 PFU) 3 months after NK cell transfer. P values, log-rank Mantel-Cox test. (c) Survival of the recipients in a (n = 19 per group) infected with influenza A strain A/PR/8/34 (500 PFU) 2 months after assessment of DTH; results are correlated to DTH. *P = 0.0001 (Spearman correlation). (d) Survival of Rag1−/− mice (n = 8–12 per group) immunized with PR8-VLPs, M1-VLPs or UV-VSV, then challenged 1 month later with live virus (2,500 PFU influenza strain A/PR/8/34 intranasally or 500 PFU VSV intravenously). P values, log-rank Mantel-Cox test. (e) Survival of Rag2−/− mice (n = 15–22 per group) immunized with VLPs containing influenza (PR8-VLP) or HIV-1 (HIV-VLP) or with UV-VSV, then challenged 1 month later by intramuscular injection of VSV at the median lethal dose (250 PFU). *P = 0.0116 (log-rank Mantel-Cox test). (f) Virus-specific DTH in Rag1−/− mice (n = 10–15 per group) immunized with HA-containing VLPs (PR8) or HA-free VLPs (M1), then challenged 1 month later and analyzed as in a. P values, unpaired Student's t-test. Data are representative of three to five independent experiments (pooled results; error bars, s.d.).
NK cell–mediated protection against lethal viral infection
As the findings reported above indicated that hepatic NK cells specifically recognize noninfectious viral antigens, we determined whether antiviral NK cell memory confers protection against lethal viral challenge. Indeed, we found that Rag2−/−Il2rg−/− mice given 8 × 104 PR8-VLP–primed hepatic NK cells and challenged 3 months later with a lethal dose of influenza strain A/PR/8/34 survived longer than naive mice did (median survival time, 36 d and 23 d, respectively). In contrast, primed splenic or naive hepatic NK cells did not confer protection (Fig. 2b).
When we first used recipients of PR8-VLP–sensitized hepatic NK cells in DTH assays with PR8-VLPs and 2 months later infected the mice with intact virus, we found the magnitude of the DTH-induced ear swelling correlated with the degree of protection against subsequent lethal infection (Fig. 2c). Analogous to our findings obtained with haptens, PR8-VLP–sensitized splenic NK cells conferred very low or absent DTH responses and failed to protect against influenza infection, which indicated that memory NK cells are excluded from the spleen regardless of their antigen specificity. Of note, recipients of NK cells purified from lungs of PR8-VLP–sensitized donors also showed a modest DTH response and protection against viral challenge, albeit to a lesser degree than that of recipients of hepatic NK cells (Fig. 2c); this indicated that some memory NK cells reside in the lungs. Notably, the protection afforded by NK cells was virus specific, as immunization of Rag1−/− mice with two different formulations of influenza VLPs (VLPs containing both HA and M1 (PR8-VLPs) or HA-free VLPs containing M1 (M1-VLPs)) protected vaccinated mice from lethal challenge with influenza but did not protect them from lethal challenge with VSV (Fig. 2d). Analogously, Rag1−/− mice immunized with UV-VSV survived longer than PBS-treated controls did after challenge with VSV but not after challenge with influenza.
Although the experiments reported above suggested that NK cells may be likely vaccine targets, all recipients of memory NK cells eventually succumbed to VSV infection (Fig. 2d). This could have reflected an inability of memory NK cells to confer sterilizing immunity or the number of memory NK cells may have been insufficient for the relatively high load and systemic dissemination of VSV in our protocol (500 plaque-forming units (PFU) administered intravenously). To distinguish between those possibilities, we monitored the survival of UV-VSV–immunized and naive Rag2−/− BALB/c mice in response to localized infection with the median lethal dose for this strain (250 PFU VSV administered intramuscularly). After challenge with this protocol, nearly all immunized mice survived, whereas about half of the naive mice and mice immunized with non-VSV antigens (such as VLPs containing influenza or HIV) died, as expected (Fig. 2e). Thus, in the absence of T cells and B cells, memory NK cells can provide effective and specific protection against certain viruses, at least when the infectious agent is administered locally and at a moderate dose.
NK cell sensitization to influenza A does not require HA
Influenza A–encoded HA is a ligand for Nkp46, an activating receptor on mouse (and human) NK cells8,9,33. However, Nkp46 is found on all NK cells regardless of anatomic localization, which seems at odds with our finding that only hepatic (and pulmonary) NK cells, but not splenic NK cells, developed influenza-specific memory. Therefore, we determined whether HA recognition was required for influenza-specific memory NK cells by immunizing Rag1−/− mice with the HA-free M1-VLPs (Supplementary Fig. 2) and comparing the ensuing DTH response with that elicited by the VLPs containing both HA and M1 (PR8-VLPs). Regardless of their HA content, both VLPs efficiently cross-sensitized mice (Fig. 2f), which indicated that NK cells do not need HA to recognize influenza. Indeed, immunization of Rag1−/− mice with either HA+ PR8-VLPs or HA− M1-VLPs significantly and equivalently prolonged the survival of the mice after lethal challenge with influenza but not after lethal challenge with VSV (Fig. 2d). Although the findings reported above do not rule out the possibility of a role for Nkp46-HA interactions, they indicate that this pathway is neither required nor sufficient for protection against influenza. However, it is possible that mice have evolved other pattern receptors for influenza and/or VSV, as mice are susceptible to these viruses14. Thus, we wondered if mouse NK cells could recognize and remember a virus that could not have imposed evolutionary pressure on mice.
Mouse NK cells develop memory of HIV-1
We addressed the question noted above by sensitizing Rag1−/− mice with VLPs containing HIV-1-derived group antigen and envelope protein34. HIV-1 is a lentivirus that causes acquired immunodeficiency syndrome (AIDS) in humans, whereas mice are resistant to HIV-1 infection35. One month after sensitizing donor mice with HIV-1-containing VLPs, we transferred splenic or hepatic NK cells into naive Rag2−/−Il2rg−/− recipients and challenged the ears of recipient mice 1 month later by injecting either HIV-1-containing VLPs or PR8-VLPs. Primed hepatic NK cells, but not splenic or naive NK cells, mounted a vigorous recall response to HIV-1 but not to influenza A (Fig. 3a). Moreover, RAG-independent memory of HIV-1 and influenza was inducible in both C57BL/6 and BALB/c mice and was always specific for the virus used during sensitization (Fig. 3b), which indicated that hepatic NK cells can develop specific memory of diverse viral antigens regardless of viral host restrictions or genetic background.

(a) Ear swelling in naive Rag2−/−Il2rg−/− mice (n = 12–15 per group) that received adoptively transferred hepatic (left) or splenic (right) CD45+NK1.1+ NK cells (8 × 104 cells per mouse) from Rag1−/− donor mice immunized with VLPs containing influenza (PR8) or HIV-1 (HIV) 1 month before transfer; recipients were challenged by subcutaneous injection of VLPs into one ear and PBS in the other ear and were assessed 2 months after transfer. NS, not significant; *P < 0.01 and **P < 0.001 (unpaired Student's t-test). (b) Ear swelling in C57BL/6 Rag1−/− mice (left) and BALB/c Rag2−/− mice (right) immunized with VLPs and challenged 1 month later (n = 10–15 mice per group). P values, unpaired Student's t-test. Background ear swelling in nonimmunized mice was subtracted from ear swelling in the experimental groups. Data are representative of three to five independent experiments (pooled results; error bars, s.d.).
CXCR6 in hepatic NK cell–mediated CHS
Next we set out to explore why memory NK cells are apparently restricted to the liver. This organ provides a specialized milieu for NKT cells, which require CXCL16, the chemokine ligand for CXCR6 that is constitutively expressed on hepatic sinusoidal endothelium22,36. CXCR6 is also expressed on hepatic NK cells22, but its importance in NK cell biology has been unclear. For these analysis we used mice in which one allele (Cxcr6+/−) or both alleles (Cxcr6−/−) of Cxcr6 were replaced with cDNA encoding GFP22. We determined that in Cxcr6+/− mice, 35–55% of hepatic NK cells, but only 3–5% of splenic NK cells, expressed CXCR6. This frequency was unchanged in Rag1−/−Cxcr6+/− mice (Fig. 4a) in which NK cells were the only GFP+ hepatic leukocytes (data not shown). To assess the role of CXCR6, we explored its contribution to hapten-induced CHS with three complementary approaches: adoptive transfer of sensitized Cxcr6+/− NK cells sorted into CXCR6-expressing (GFP+) and non–CXCR6-expressing (GFP−) subsets (Fig. 4b and Supplementary Fig. 3a–c); direct sensitization and challenge of wild-type mice and their Rag1−/−Cxcr6−/− and Rag1−/−Cxcr6+/− littermates (Fig. 4c and Supplementary Fig. 4a); and treatment of sensitized wild-type or Rag1−/− mice with antibody to CXCR6 (anti-CXCR6; Fig. 4d and Supplementary Fig. 4b).

(a) Frequency of CXCR6-expressing CD45+ NK1.1+ NK cells from Cxcr6+/− mice on a Rag1-sufficient (C57BL/6) or Rag1−/− background in different tissues, assessed by flow cytometry. LN, lymph node; BM, bone marrow. (b) Ear swelling in naive Rag2−/−Il2rg−/− mice (n = 10–12 per group) that received 1 × 105 NK cells from DNFB-sensitized Rag1−/−Cxcr6+/− donor spleen or liver, sorted for expression of NK1.1 and GFP; recipient mice were challenged 1 month later with DNFB on one ear and solvent on the other. (c) DNFB-induced CHS in lymphocompetent C57BL/6 mice (left) and Rag1−/− C57BL/6 mice (right; n = 10–12 mice per group). (d) DNFB-induced CHS in C57BL/6 mice (left) and Rag1−/− C57BL/6 mice (right) sensitized with hapten and given mAb to CXCR6 (100 μg per mouse) or isotype-matched control antibody intravenously 24 h before DNFB challenge (n = 10–15 mice per group). *P < 0.01, **P < 0.001 and ***P < 0.0001 (unpaired Student's t-test (a,c,d) or ANOVA (b)). Data are representative of three to five independent experiments (pooled results; error bars, s.d.).
After adoptive transfer, all NK cell subsets proliferated equivalently in naive Rag2−/−Il2rg−/− hosts (Supplementary Fig. 3b), but only primed GFP+ hepatic NK cells elicited CHS responses (Fig. 4b and Supplementary Fig. 3a); this indicated that memory NK cells were concentrated in the CXCR6+ subset. However, adoptive transfer of GFP+ splenic NK cells failed to mediate CHS, which suggested that CXCR6 expression is insufficient to develop memory. After transfer, GFP+ hepatic NK cells persisted unchanged for at least 6 weeks (Supplementary Fig. 3c), whereas we noted some loss of GFP among GFP+ splenic NK cells, and ∼20% of liver-derived GFP− NK cells (but not splenic GFP− NK cells) became GFP+. It is unclear whether this conversion reflected differentiation of mature NK cells or new development from GFP− precursors.
Although CXCR6 alone was apparently insufficient to enable NK cell memory, the results of our adoptive-transfer experiments were consistent with a role for this receptor NK cell–mediated adaptive immunity. Thus, we compared the responsiveness of Cxcr6+/− and Cxcr6−/− mice to DNFB and OXA. CHS responses were much lower but not abolished in Cxcr6−/− mice that had T cells and B cells (Fig. 4c, left), whereas Rag1−/−Cxcr6−/− mice were unresponsive to either DNFB (Fig. 4c, right) or OXA (Supplementary Fig. 4a). Hence, in the absence of T cells and B cells, NK cell–expressed CXCR6 is absolutely required for NK cell–mediated CHS, whereas wild-type mice depend only partially on CXCR6. Because depletion of NK cells in wild-type mice does not result in lower CHS responses17, the compromised response in Cxcr6−/− mice probably reflected a combined effect on NK cells and NKT cells37.
The results reported above were consistent with three nonexclusive roles for CXCR6 in the development or survival, the trafficking, or the effector function of memory NK cells. To address the last possibility, we gave sensitized mice neutralizing monoclonal antibody (mAb) to CXCR6 (Supplementary Fig. 4b) or isotype-matched control mAb 12 h before hapten challenge and compared their CHS responses. Anti-CXCR6 decreased the CHS responses of wild-type mice to DNFB (Fig. 4d, left) and OXA (Supplementary Fig. 4c, left) and abolished such responses in sensitized Rag1−/− mice (Fig. 4d, right, and Supplementary Fig. 4c, right). This effect of anti-CXCR6 was not due to depletion of NK cells, and short-term treatment with mAb did not result in the relocalization of Cxcr6+/− NK cells to organs other than the liver (Supplementary Fig. 4b). After treatment of wild-type or Rag1−/− mice with anti-CXCR6 or anti-CXCL16, the number of Cxcr6+/− NK cells recovered from various organs was similar to that of mice treated with istoype-matched control mAb.
Having established that memory NK cells require CXCR6 for CHS, we sought to determine if this pathway is involved in antiviral NK cell memory. We sensitized Rag1−/− mice to UV-VSV or to VLPs containing influenza or HIV-1 and treated the mice with anti-CXCR6 or isotype-matched control antibody 12 h before challenging them by injecting viral recall antigen into one ear and PBS into the other. The ensuing DTH response was abolished after CXCR6 blockade regardless of the type of viral antigen or the MHC class I haplotype of the mice (Fig. 5a). Furthermore, anti-CXCR6 abolished the protective effect of memory NK cells after lethal challenge with influenza or VSV (Fig. 5b). Thus, for each of the five antigenic entities tested, CXCR6 was essential for NK cell–mediated adaptive immunity regardless of the genetic background.

(a) Antiviral DTH responses in Rag1−/− C57BL/6 mice (left) or Rag2−/− BALB/c mice (right) immunized and challenged with various combinations of VLPs and UV-VSV (below graphs) and given mAb to CXCR6 (100 μg per mouse) or isotype-matched control antibody 24 h before challenge. P values, unpaired Student's t-test. (b) Survival of Rag1−/− and Rag2−/− mice (n = 8–12 per group) immunized with PR8-VLP or M1-VLP, challenged 1 month later by lethal infection with influenza A strain A/PR/8/34 (2,500 PFU for Rag1−/− (left) and 10,000 PFU for Rag2−/− (right)) and injected with mAb to CXCR6 (100 μg per mouse) or isotype-matched control antibody on days 1 and 5. P values, log-rank Mantel-Cox test. Data are representative of three to five independent experiments (pooled results; error bars, s.d.).
Role of CXCR6 in the homeostasis of memory NK cells
CXCR6 may exert this critical activity, at least in part, by regulating the homeostasis of hepatic NK cells, similar to its role in the survival of hepatic NKT cells22. Steady-state livers of Cxcr6−/− mice contained normal numbers of GFP− NK cells but significantly fewer GFP+ NK cells than did those of Cxcr6+/− mice (Fig. 6a,b and Supplementary Fig. 5a), and the frequency of the small fraction of GFP+ NK cells in other organs was the same for both genotypes (Fig. 6a). Thus, CXCR6 is required for the development and/or survival of CXCR6+ NK cells exclusively in the liver, where CXCL16 is constitutively expressed. Furthermore, 4 weeks after adoptive transfer of equal numbers of GFP+ and GFP− hapten-sensitized NK cells from Cxcr6+/− donors, the GFP− NK cells were distributed equally in spleen and liver, whereas the distribution of the GFP+ subset was strongly biased toward the liver (Fig. 6c). Moreover, when we transferred equal numbers of GFP+ and GFP− hepatic NK cells sorted from Cxcr6−/− or Cxcr6+/− donors, the Cxcr6+/− NK cell populations expanded in the livers of Rag2−/−Il2rg−/− recipients regardless of their GFP expression, whereas in recipients of CXCR6-deficient NK cells, only the GFP− subset expanded, whereas GFP+ NK cells were very rare (Fig. 6d). Furthermore, adoptive transfer of DNFB-primed GFP+ hepatic NK cells to naive hosts conferred hapten responsiveness only when NK cells were from Cxcr6+/− donors, whereas the same number of Cxcr6−/− NK cells did not transfer hapten sensitivity (Fig. 6e) and survived poorly even after rechallenge with DNFB (Supplementary Fig. 5b).

(a) Frequency of GFP+ and GFP− NK cell subsets (identified as CD45+NK1.1+ cells) in various organs of Cxcr6+/− and Cxcr6−/− mice. *P < 0.01 and **P < 0.001 (unpaired Student's t-test). (b) Ratio of NK cell subsets (CXCR6+/CXCR6−) in liver and spleen of wild-type (Cxcr6+/+), Cxcr6+/− and Cxcr6−/− mice (all C57BL/6; n = 12–15 per group). *P < 0.00001 (unpaired Student's t-test). (c) Distribution of NK cells recovered from liver or spleen 1 month after adoptive transfer of sorted subsets (1 × 105 cells) into Rag2−/−Il2rg−/− recipients (n = 10–15 per group). *P < 0.001 and **P < 0.0001 (unpaired Student's t-test). (d) Flow cytometry of NK cells in livers and spleens of Rag2−/−Il2rg−/− recipients (n = 8–10 per group) 2 weeks after transfer of mixtures of 1 × 105 GFP+ or GFP− CD45+NK1.1+ NK cells sorted from Cxcr6+/− or Cxcr6−/− donors. *P < 0.001 and **P < 0.0001 (unpaired Student's t-test). (e) Ear swelling in Rag2−/−Il2rg−/− mice (n = 8 per group) that received 8 × 104 DNFB-primed CD45+ NK1.1+ GFP+ NK cells from Cxcr6+/− or Cxcr6−/− donors and were challenged 24 h or 1 month later. *P < 0.01 and **P < 0.001 (unpaired Student's t-test). Data are representative of three to five independent experiments (pooled results; error bars, s.d.).
NK cell–mediated hapten-specific cytotoxicity
Given that a relatively short (12-hour) mAb-induced inhibition of CXCR6 efficiently blocked NK cell–mediated responses to haptens and viruses in vivo, we determined whether CXCR6 regulates acute NK cell effector functions such as degranulation and killing. We cultured naive or primed hepatic or splenic NK cells together with a mixture of equal numbers of target B cells whose surfaces had been modified with DNBS (a PBS-soluble analog of DNFB) and mock-treated control B cells. We then determined the ratio of control B cells to haptenated B cells in the presence or absence of NK cells 12 h later. Only DNFB-sensitized hepatic NK cells killed DNBS-haptenated targets; no other subset showed hapten-specific killing activity (Fig. 7a). Furthermore, only the CXCR6+ hepatic NK cell subset was able to kill DNBS-modified targets (Fig. 7b), in agreement with our CHS-DTH experiments. However, all NK cell samples killed MHC class I–deficient target cells with similar efficiency, which indicated that no subset was compromised in its ability to kill appropriate targets per se.

(a) Hapten-specific killing of target B cells by naive and hapten-sensitized CD45+ NK1.1+ NK cells cultured for 12 h at various target cell/effector cell ratios (horizontal axis) with a mixture of two populations of B cells labeled with a large (CFSEhi) or small (CFSElo) amount of the cytosolic dye CFSE (n = 10–20 donor mice per group); alternatively, target and control B cells were distinguished by use of the congenic markers CD45.1 and CD45.2. CFSElo or CD45.1+ B cells served as a control; CFSEhi or CD45.2+ B cells were from wild-type donors and were haptenated with DNBS (left and middle) or were from MHC class I–deficient (MHC-KO) donors (right). Hapten-specific killing was assessed as the ratio of CFSElo to CFSEhi cells, corrected for input. *P < 10−8 and **P < 10−12, DNFB- or OXA-sensitized versus acetone (left and middle) or MHC-KO versus DNBS-labeled (right; unpaired Student's t-test). (b) Killing capacity of DNFB-primed hepatic CD45+ NK1.1+ NK cells from Cxcr6+/− or Cxcr6−/− donor mice (n = 12 donor mice per group), assessed as in a in the presence of mAb to CXCR6 or isotype-matched control mAb. *P < 0.01, **P < 0.001 and ***P < 0.00001, compared with Cxcr6−/− (unpaired Student's t-test). (c) Killing capacity of acetone- or DNFB-primed hepatic CD45+ NK1.1+ NK cells from Rag1−/− donors (n = 15 per group) at a target cell/effector cell ratio of 1:25, assessed in the presence of mAb to CXCR6 (10 μg/ml), mAb to CXCL16 (10 μg/ml) or CXCL16 (500 ng/ml); results are presented relative to those of cultures treated with isotype-matched control antibody (10 μg/ml). *P < 0.01 and **P < 0.001, compared with isotype-matched control antibody (unpaired Students t-test). (d) Flow cytometry analysis of the incorporation of anti-LAMP-1 by NK1.1+ NK cells sorted from the livers or spleens of Rag1−/− donor mice sensitized with acetone, DNFB or OXA on days 0 and 1 and injected with 100 μg mAb to CXCR6 or isotype-matched control mAb 12 h before NK cell isolation; NK cells were cultured together with DNBS-labeled B cells in the presence of fluorescein isothiocyanate–conjugated anti-LAMP-1 (10 μg/ml) with or without mAb to CXCR6 or isotype-matched control mAb (10 μg/ml) and assessed after 3 h (n = 10–18 donor mice total with 12–20 wells per group). (e) Frequency of LAMP-1+ NK cells among the cells in d. *P < 10−9 (unpaired Student's t-test). Data are representative of three to five independent experiments (pooled results; error bars, s.d.).
The addition of mAb to CXCR6 to cytotoxicity assays boosted the killing of haptenated B cells by Cxcr6+/− NK cells, especially at suboptimal ratios of target cells to effector cells (Fig. 7b,c). We therefore compared NK cell–mediated hapten-specific killing in the presence of blocking mAbs to CXCR6 or CXCL16 or in the presence of recombinant CXCL16 at a target cell/effector cell ratio of 1:25. Blockade of the CXCR6-CXCL16 pathway significantly enhanced the cytotoxicity of sensitized hepatic NK cells, whereas the addition of CXCL16 significantly dampened the response (Fig. 7c). We then measured the frequency of actively degranulating NK cells by assessing transient surface expression of lysosomal marker LAMP-1 (CD107a)38; about 10% of DNFB-sensitized hepatic NK cells degranulated after exposure to DNBS-modified target cells, whereas naive DNFB-sensitized splenic NK cells and OXA-sensitized splenic and hepatic NK cells did not respond (Fig. 7d,e and Supplementary Fig. 6). Notably, the addition of anti-CXCR6 during the 3 h of in vitro coculture did not affect degranulation, whereas DNFB-sensitized NK cells obtained from mice 12 h after in vivo administration of anti-CXCR6 failed to upregulate LAMP-1. In conclusion, beyond the widespread ability of NK cells to respond to 'missing self'39, the hepatic memory NK cell subset has the unique ability to also exert cytotoxic activity after encountering MHC class I–sufficient target cells decorated with an antigen to which NK cells were previously sensitized. This adaptive ability seemed to be rapidly lost in vivo when CXCR6 signaling was disrupted, even though CXCR6 was apparently not required for hapten recognition but instead attenuated the cytotoxic effector activity of memory NK cells during the in vitro assay.
Discussion
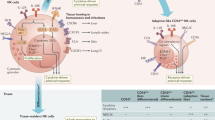
Our study has identified a subset of hepatic NK cells that acquired antigen-specific memory of at least five structurally, chemically and biologically distinct antigens. Adaptive immune responses mediated by this population depended on CXCR6, which was expressed on about half of the hepatic NK cells; this suggests that the liver has two different NK cell populations. CXCR6− NK cells shared features with splenic NK cells; their survival and homeostasis was CXCR6 independent and they did not develop memory of haptens or viruses (except perhaps MCMV). In contrast, the second hepatic NK cell subset constitutively transcribed the Cxcr6 locus and contained cells that acquired long-lived specific memory of diverse antigens, including haptens and complex assemblies of viral glycoproteins. The paucity of GFP+ NK cells in livers of Cxcr6−/− mice indicated that the CXCR6+ subset, unlike CXCR6− NK cells, required sustenance from sinusoidal endothelium, which constitutively presents CXCL16, a chemokine that exists as both a secreted polypeptide and a luminal surface transmembrane protein.
The restricted steady-state distribution of CXCL16, the only known ligand of CXCR6, may explain why memory NK cells are undetectable in blood40 or spleen17. However, the few CXCR6+ splenic NK cells were unable to mediate recall responses, which indicates that CXCR6 is required but not sufficient to induce and/or maintain memory. Indeed, the role of CXCR6 seems complex. Injection of anti-CXCR6 24 h before antigen challenge inhibited NK cell–mediated recall responses in vivo without resulting in depletion of CXCR6+ NK cells. As antigen recognition by NK cells remained intact after exposure to anti-CXCR6 in vitro, the in vivo effects most probably reflect a role for CXCR6-CXCL16 in maintaining the differentiation and/or function of memory NK cells. Moreover, anti-CXCR6 enhanced the antigen-specific cytotoxicity of memory NK cells in vitro, whereas CXCL16 attenuated cytotoxicity. In contrast, killing of MHC class I–deficient targets remained unaffected, which suggests that CXCR6 modulates a proximal signaling step downstream of antigen recognition but upstream of the terminal pathway that enables cytotoxicity triggered by activating NK cell receptors. Of note, anti-CXCR6 did not alter the frequency of the ∼10% hepatic NK cells that degranulated after antigen exposure, which suggests that anti-CXCR6 boosted killing by a finite pool of hapten-specific NK cells without increasing the size of that pool.
The hepatic milieu might have two effects on memory NK cells: continuous CXCR6 signaling, presumably triggered by transmembrane and/or secreted CXCL16 in sinusoids, promotes long-term survival and effector potential; however, tonic engagement of CXCR6 might safeguard against NK cell–mediated hepatotoxicity by preventing full-fledged killing activity. Once memory NK cells are released from the liver and access peripheral sites of antigen challenge, the absence of CXCR6 ligands may unleash their full effector potential but might also limit their persistence. However, it should be noted that CXCL16 can be upregulated on activated dendritic cells25 and in inflamed lung23 and skin41. This might provide a 'home away from home' to sustain memory NK cells at extrahepatic effector sites.
Although CXCR6 is apparently not directly involved in antigen recognition by NK cells, it will be useful to assess whether and how it regulates the underlying molecular mechanism of antigen recognition, which remains elusive. None of the known markers associated with memory NK cells17,20 are sufficient to confer hapten specificity, as they are also found on splenic NK cells. Furthermore, hapten sensitization does not change the expression of Ly49 receptors or NKG2D or mRNA for any other known activating receptor expressed by NK cells (unpublished data). In this way, hapten-specific NK cells differ from MCMV-specific memory NK cells, which upregulate the receptors KLRG1, CD43, Ly6C and Ly49H and are found in both liver and spleen13.
Although engagement of inhibitory receptors by self MHC class I usually blocks the activation of NK cells, we found that DNFB-sensitized hepatic NK cells killed DNBS-haptenated B cells that expressed MHC class I. Conceivably, covalent DNBS modification of target B cell proteins could have included MHC class I molecules. However, this effect could not have masked the recognition of MHC class I by NK cells because DNBS-modified B cells were not lysed by splenic or OXA-sensitized hepatic NK cells, which killed MHC class I–deficient targets efficiently. Thus, the pathways triggered in memory NK cells by cognate antigen over-ride inhibitory signals from MHC class I. Moreover, NK cell memory of haptens and viruses developed in both BALB/c and C57BL/6 mice, which express distinct MHC and Ly49 haplotypes; this suggests that antigen recognition by hepatic memory NK cells differs from the way conventional NK cells detect pathological cells with altered or absent MHC class I.
Whatever the mechanism, the finding that mice deficient in T cells and B cells can develop memory that is protective against lethal infection with influenza A or VSV has implications for vaccine development and antiviral immunity. These results are consistent with investigations of NK cell memory of MCMV13, although there are differences between the tissue distribution of the ensuing memory NK cells, genetic background restriction and possibly also the virus-recognition mechanism. NK cells in C57BL/6 mice express Ly49H, a germline-encoded activating receptor that is necessary for recognition and memory of MCMV13. Similarly, mouse and human NK cells use NKp46 to detect influenza-derived HA8,9. However, we found that hepatic NK cells developed influenza-specific memory without exposure to HA, whereas splenic NKp46+ NK cells did not develop memory. Thus, NKp46 is neither required nor sufficient to generate influenza-specific memory NK cells.
We cannot rule out the possibility that NK cells express other as-yet-unidentified germline-encoded receptors that recognize influenza or VSV antigens. Such receptors could conceivably have evolved, as mice are susceptible to these viruses, although neither is endemic in wild mice14. However, mouse NK cells also recognized and remembered HIV-1, which does not infect mice and therefore is unlikely to have driven the evolution of specific recognition mechanisms.
Of note, specific NK cell haplotypes are over-represented in HIV-infected people who exert long-term control over HIV-1 replication and progression to AIDS15,42. These patients have a subset of NK cells that express the receptor KIR3DS1, which inhibits HIV-1 replication in vitro16. Notably, a multiple-cohort genome-wide association study has identified the rs2234358 polymorphism in CXCR6 as another marker strongly associated with long-term nonprogression to AIDS that is independent of other genetic polymorphisms43. CXCR6 is a minor coreceptor in HIV-1 infection, but it is unclear if this function is responsible for its role in HIV progression. Although CXCR6 is expressed on some human NK cells44, it is unknown whether CXCR6+ NK cell populations or other human NK cell populations have distinct functional properties, such as the ability to carry antigen-specific memory. If such functions exist, they could potentially be exploited to develop new vaccines or to treat primary or acquired immunodeficiencies.
Methods
Mice.
Rag1−/− (C57BL/6), Rag2−/− (BALB/c), Rag2−/−Il2rg−/− (C57BL/6 × C57BL/10 F1)45, Cxcr6−/− (C57BL/6) and C57BL/6 mice were used at 6–12 weeks of age (Taconic, Jackson Laboratories and Charles River Laboratories) according to the institutional animal committees at Harvard Medical School. Mice transgenically expressing cDNA for enhanced GFP under control of the actin promoter and Cxcr6+/− and Cxcr6−/− mice on the C57BL/6 Rag1−/− background were generated and bred in-house.
Cell isolation and sorting.
Spleens, lungs, lymph nodes, bone marrow and livers were cut into small pieces with a sterile scalpel and passed through 40-μm mesh filters. For analysis of skin-infiltrating lymphocytes, mouse ears were collected and then ear sheets were mechanically pulled apart, cut into small pieces with a sterile scalpel, digested with collagenase D (5 mg/ml in 2% (vol/vol) FBS in PBS) and passed through 40-μm mesh filters. Samples were enriched for lymphocytes by density-gradient centrifugation with Nycodenz according to the manufacturer's protocol (Cederlane Labs). For flow cytometry analysis or cell sorting, cells were stained with anti-NK1.1 (PK136; Biolegend), anti-TCRβ (H57597; Pharmingen) and anti-TCRδ (GL-3; Pharmingen) or with anti-CD3 (145-2C11; Biolegend), anti-Thy-1 (30H12; Biolegend), anti-CD45.1 (A20; Pharmingen), anti-CD45.2 (104; Pharmingen) anti-LAMP-1 (1D4B; Pharmingen), anti-CXCR6 (221002; R&D Systems), anti-CXCL16 (142417; R&D Systems), rat IgG2a (54447; R&D Systems) and rat IgG2b (141945; R&D Systems), and samples were acquired on a FACSCanto (BD) and analyzed with FlowJo software. NK cells were identified as CD45+, NK1.1+, CD3− or TCR− and sometimes also Thy-1+ and/or GFP+. A FACSAria (BD) was used for cell sorting with Diva software, and purity was >98% for all experiments.
CHS and DTH.
Mice were sensitized days 0 and 1 by painting of skin on the shaved abdomen with 50 μl 0.5% (wt/vol) DNFB (dinitrofluorobenzene) in acetone or 50 μl 5% (wt/vol) OXA in a solution of acetone and methanol (1:1). Control mice received solvent only (Sigma-Aldrich). On day 4, mice were either used as donors for cell sorting or their right ear was challenged with 20 μl 0.2% (wt/vol) DNFB in acetone or 20 μl 1% (wt/vol) OXA in acetone and methanol and their left ear was painted with vehicle. Viral DTH was induced by subcutaneous immunization of Rag1−/− or Rag2−/− mice days 0 and 7 with PBS (control) or 5 μg viral antigen. One month later, mice were either used as donors for adoptive-transfer experiments or challenged subcutaneously with 25 μl PBS containing 2 μg viral antigen into one ear and PBS into the control ear. Ear thickness was measured every 24 h with a micrometer (193-101l Mitutoyo). To account for acute hapten- or virus-induced irritation, background swelling was measured in naive mice and sensitization-dependent, antigen-specific ear swelling was calculated as follows: (treated ear thickness − control ear thickness) − background swelling.
Generation of VLPs and influenza virus for live virus challenge.
VLPs were generated as described30,34. Spodoptera frugiperda (Sf9) cells were coinfected with recombinant baculovirus expressing the influenza proteins HA and/or M1 or the HIV group antigen and envelope proteins. Culture supernatants were collected 3 d after infection and cleared by low-speed centrifugation (2,000g for 20 min at 4 °C), then VLPs were concentrated by cross-flow filtration and purified through a 20–30–60% discontinuous sucrose gradient at 100,000g for 1 h at 4 °C. VLP bands were collected, dialyzed against PBS and analyzed by immunoblot. Mouse-adapted influenza strain A/PR/8/34 was grown once in 10-day-old embryonated hen's eggs and was used for live virus challenge. Anesthetized mice were infected intranasally by instillation of 50 μl containing influenza stain A/PR/8/34 in PBS (at various PFU values). Alternatively, 500 PFU of VSV was injected intravenously or 250 PFU of VSV was injected intramuscularly. Control mice received PBS only.
In vitro killing and LAMP-1 upregulation.
NK cells were sorted from hapten-sensitized donors at day 4 and were cultured at various ratios together with a 1:1 mixture of autologous DNBS (dinitrobenzene sulfonic acid)-labeled or MHC class I–deficient B cells and unmodified wild-type control B cells. Target and control cells were distinguished by CFSE (carboxyfluorescein diacetate succinimidyl ester) labeling or by expression of the congenic markers CD45.1 and CD45.2. Then, 12 h after coincubation, the ratio of target cells to control cells was determined by flow cytometry. Specific lysis was calculated as follows: (1 − (control / target)input / (control / target)NK-exposed]) × 100%, where (control / target)input is the ratio of control cells to target cells in the absence of NK, and (conrol / target)NK-exposed is the ratio of control cells to target cells after exposure to NK cells. For analysis of LAMP-1 upregulation on NK cells, NK cells were cultured together with DNBS-labeled B cells and fluorescein isothiocyanate–conjugated mAb specific for LAMP-1 (10 μg/ml) in the presence or absence of mAb to CXCR6 (10 μg/ml), isotype-matched control antibody, or CXCL16 protein, and analyzed by flow cytometry for incorporation of anti-LAMP-1 after 3 h.
DNBS labeling of B cell targets.
Naive B cells were isolated by negative selection with CD43 magnetic beads (Miltenyi Biotech) and were suspended at a density of 1 × 108 cells per ml in PBS. DNBS in PBS was added to a final concentration of 20 mg/ml, then cells were incubated for 10 min at 25 °C and washed twice with 10% (vol/vol) FBS in PBS.
Statistical analysis.
Statistical significance was determined by Student's t-test, ANOVA, Spearman correlation or a log-rank (Mantel-Cox) test. Significance was set at a P value of less than 0.05.
References
Bassing, C.H., Swat, W. & Alt, F.W. The mechanism and regulation of chromosomal V(D)J recombination. Cell 109 Suppl, S45–S55 (2002).
Crowle, A.J. Delayed hypersensitivity in mice; its detection by skin tests and its passive transfer. Science 130, 159–160 (1959).
Orme, I.M. & Cooper, A.M. Cytokine/chemokine cascades in immunity to tuberculosis. Immunol. Today 20, 307–312 (1999).
Dhabhar, F.S., Satoskar, A.R., Bluethmann, H., David, J.R. & McEwen, B.S. Stress-induced enhancement of skin immune function: A role for γ interferon. Proc. Natl. Acad. Sci. USA 97, 2846–2851 (2000).
Phanuphak, P., Moorhead, J.W. & Claman, H.N. Tolerance and contact sensitivity to DNFB in mice. II. Specific in vitro stimulation with a hapten, 2,4-dinitrobenzene sulfonic acid (DNB-SO3Na). J. Immunol. 112, 849–851 (1974).
Lanier, L.L. Up on the tightrope: natural killer cell activation and inhibition. Nat. Immunol. 9, 495–502 (2008).
Lanier, L.L. NK cell recognition. Annu. Rev. Immunol. 23, 225–274 (2005).
Mandelboim, O. et al. Recognition of haemagglutinins on virus-infected cells by NKp46 activates lysis by human NK cells. Nature 409, 1055–1060 (2001).
Gazit, R. et al. Lethal influenza infection in the absence of the natural killer cell receptor gene Ncr1. Nat. Immunol. 7, 517–523 (2006).
Smith, H.R. et al. Recognition of a virus-encoded ligand by a natural killer cell activation receptor. Proc. Natl. Acad. Sci. USA 99, 8826–8831 (2002).
Kim, S. et al. Licensing of natural killer cells by host major histocompatibility complex class I molecules. Nature 436, 709–713 (2005).
Jonsson, A.H. & Yokoyama, W.M. Natural killer cell tolerance licensing and other mechanisms. Adv. Immunol. 101, 27–79 (2009).
Sun, J.C., Beilke, J.N. & Lanier, L.L. Adaptive immune features of natural killer cells. Nature 457, 557–561 (2009).
Scalzo, A.A., Manzur, M., Forbes, C.A., Brown, M.G. & Shellam, G.R. NK gene complex haplotype variability and host resistance alleles to murine cytomegalovirus in wild mouse populations. Immunol. Cell Biol. 83, 144–149 (2005).
Martin, M.P. et al. Epistatic interaction between KIR3DS1 and HLA-B delays the progression to AIDS. Nat. Genet. 31, 429–434 (2002).
Alter, G. et al. Differential natural killer cell-mediated inhibition of HIV-1 replication based on distinct KIR/HLA subtypes. J. Exp. Med. 204, 3027–3036 (2007).
O'Leary, J.G., Goodarzi, M., Drayton, D.L. & von Andrian, U.H. T cell– and B cell–independent adaptive immunity mediated by natural killer cells. Nat. Immunol. 7, 507–516 (2006).
Boehncke, W.H. et al. Leukocyte extravasation as a target for anti-inflammatory therapy—Which molecule to choose? Exp. Dermatol. 14, 70–80 (2005).
DiSanto, J.P., Muller, W., Guy-Grand, D., Fischer, A. & Rajewsky, K. Lymphoid development in mice with a targeted deletion of the interleukin 2 receptor γ chain. Proc. Natl. Acad. Sci. USA 92, 377–381 (1995).
Paust, S., Senman, B. & von Andrian, U.H. Adaptive immune responses mediated by natural killer cells. Immunol. Rev. 235, 286–296 (2010).
Matangkasombut, P. et al. Lack of iNKT cells in patients with combined immune deficiency due to hypomorphic RAG mutations. Blood 111, 271–274 (2008).
Geissmann, F. et al. Intravascular immune surveillance by CXCR6+ NKT cells patrolling liver sinusoids. PLoS Biol. 3, e113 (2005).
Day, C.E. et al. A novel method for isolation of human lung T cells from lung resection tissue reveals increased expression of GAPDH and CXCR6. J. Immunol. Methods 342, 91–97 (2009).
Matloubian, M., David, A., Engel, S., Ryan, J.E. & Cyster, J.G. A transmembrane CXC chemokine is a ligand for HIV-coreceptor Bonzo. Nat. Immunol. 1, 298–304 (2000).
van der Voort, R. et al. An alternatively spliced CXCL16 isoform expressed by dendritic cells is a secreted chemoattractant for CXCR6+ cells. J. Leukoc. Biol. 87, 1029–1039 (2010).
Sato, T. et al. Role for CXCR6 in recruitment of activated CD8+ lymphocytes to inflamed liver. J. Immunol. 174, 277–283 (2005).
Park, B.K., Tingle, M.D., Grabowski, P.S., Coleman, J.W. & Kitteringham, N.R. Drug-protein conjugates–XI. Disposition and immunogenicity of dinitrofluorobenzene, a model compound for the investigation of drugs as haptens. Biochem. Pharmacol. 36, 591–599 (1987).
Di Santo, J.P., Colucci, F. & Guy-Grand, D. Natural killer and T cells of innate and adaptive immunity: lymphoid compartments with different requirements for common gamma chain-dependent cytokines. Immunol. Rev. 165, 29–38 (1998).
Jamieson, A.M., Isnard, P., Dorfman, J.R., Coles, M.C. & Raulet, D.H. Turnover and proliferation of NK cells in steady state and lymphopenic conditions. J. Immunol. 172, 864–870 (2004).
Quan, F.S., Huang, C., Compans, R.W. & Kang, S.M. Virus-like particle vaccine induces protective immunity against homologous and heterologous strains of influenza virus. J. Virol. 81, 3514–3524 (2007).
Kang, S.M., Song, J.M., Quan, F.S. & Compans, R.W. Influenza vaccines based on virus-like particles. Virus Res. 143, 140–146 (2009).
Hangartner, L., Zinkernagel, R.M. & Hengartner, H. Antiviral antibody responses: the two extremes of a wide spectrum. Nat. Rev. Immunol. 6, 231–243 (2006).
Draghi, M. et al. NKp46 and NKG2D recognition of infected dendritic cells is necessary for NK cell activation in the human response to influenza infection. J. Immunol. 178, 2688–2698 (2007).
Wang, B.Z. et al. Incorporation of high levels of chimeric human immunodeficiency virus envelope glycoproteins into virus-like particles. J. Virol. 81, 10869–10878 (2007).
Shultz, L.D. et al. NOD/LtSz-Rag1null mice: an immunodeficient and radioresistant model for engraftment of human hematolymphoid cells, HIV infection, and adoptive transfer of NOD mouse diabetogenic T cells. J. Immunol. 164, 2496–2507 (2000).
Wilbanks, A. et al. Expression cloning of the STRL33/BONZO/TYMSTR ligand reveals elements of CC, CXC, and CX3C chemokines. J. Immunol. 166, 5145–5154 (2001).
Szczepanik, M. et al. B-1 B cells mediate required early T cell recruitment to elicit protein-induced delayed-type hypersensitivity. J. Immunol. 171, 6225–6235 (2003).
Aktas, E., Kucuksezer, U.C., Bilgic, S., Erten, G. & Deniz, G. Relationship between CD107a expression and cytotoxic activity. Cell. Immunol. 254, 149–154 (2009).
Raulet, D.H. A sense of something missing. Nature 358, 21–22 (1992).
Carbone, T. et al. CD56highCD16−CD62L− NK cells accumulate in allergic contact dermatitis and contribute to the expression of allergic responses. J. Immunol. 184, 1102–1110 (2010).
Scholz, F. et al. Constitutive expression and regulated release of the transmembrane chemokine CXCL16 in human and murine skin. J. Invest. Dermatol. 127, 1444–1455 (2007).
Martin, M.P. et al. Innate partnership of HLA-B and KIR3DL1 subtypes against HIV-1. Nat. Genet. 39, 733–740 (2007).
Limou, S. et al. Multiple-cohort genetic association study reveals CXCR6 as a new chemokine receptor involved in long-term nonprogression to AIDS. J. Infect. Dis. 202, 908–915 (2010).
Berahovich, R.D., Lai, N.L., Wei, Z., Lanier, L.L. & Schall, T.J. Evidence for NK cell subsets based on chemokine receptor expression. J. Immunol. 177, 7833–7840 (2006).
Cao, X. et al. Defective lymphoid development in mice lacking expression of the common cytokine receptor γ chain. Immunity 2, 223–238 (1995).
Acknowledgements
We thank G. Cheng and J.D. Sullivan for technical support and A. Wagers (Harvard Medical School) for Act(EGFP) mice. Supported by the US National Institutes of Health (AI069259, AI072252, AI078897, HL56949 and AR42689), the Ragon Institute (U.H.v.A.), the Cancer Research Institute (S.P.) and the Ragon Institute of MIT, Harvard and MGH (S.P.).
Author information
Authors and Affiliations
Contributions
S.P. and U.H.v.A. designed the study; S.P., H.S.G., B.Z.W. and M.F. did experiments; S.P., A.T. and B.S. collected and analyzed data; E.A.M., H.S.G., B.Z.W. and R.H.C. provided reagents; E.A.M., M.S. and P.W.A. provided technical support and conceptual advice; and S.P. and U.H.v.A. wrote the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Supplementary Text and Figures
Supplementary Figures 1–6 (PDF 1189 kb)
Rights and permissions
About this article
Cite this article
Paust, S., Gill, H., Wang, BZ. et al. Critical role for the chemokine receptor CXCR6 in NK cell–mediated antigen-specific memory of haptens and viruses. Nat Immunol 11, 1127–1135 (2010). https://doi.org/10.1038/ni.1953
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ni.1953
This article is cited by
-
Roles of natural killer cells in immunity to cancer, and applications to immunotherapy
Nature Reviews Immunology (2023)
-
Autologous dendritic cell vaccination against HIV-1 induces changes in natural killer cell phenotype and functionality
npj Vaccines (2023)
-
Transcriptomic perspectives of memory-like NK cells and aging
Genome Medicine (2022)
-
Immunoglobulin-like receptors and the generation of innate immune memory
Immunogenetics (2022)
-
IL-21 and IFNα therapy rescues terminally differentiated NK cells and limits SIV reservoir in ART-treated macaques
Nature Communications (2021)
