
Abstract
Eukaryotic ribosomal proteins, unlike their bacterial homologues, possess nuclear localization signals (NLSs) to enter the cell nucleus during ribosome assembly. Here we provide a comprehensive comparison of bacterial and eukaryotic ribosomes to show that NLSs appear in conserved ribosomal proteins via remodelling of their RNA-binding domains. This finding enabled us to identify previously unknown NLSs in ribosomal proteins from humans, and suggests that, apart from promoting protein transport, NLSs may facilitate folding of ribosomal RNA.
Similar content being viewed by others
Introduction
Ribosomes share similar architecture in all branches of life, but their assembly pathways are dramatically divergent across species, particularly between bacteria and eukaryotes. In eukaryotes, ribosome assembly requires an intricate trafficking of ribosomal proteins; produced in the cytoplasm, they first enter the cell nucleus and accumulate in the nucleolus before they associate into nascent ribosomes1. Therefore, eukaryotic ribosomal proteins are thought to harbour nuclear/nucleolar localization signals (NLSs) – short, predominantly basic stretches of amino acids that trigger active transport of proteins to the nucleus2,3,4. Given the ancient origin of ribosomes, the question arises – how did NLSs emerge in conserved ribosomal proteins? Were similar motifs present in prokaryotic proteins, and if not, what structural changes were required to evolve the NLSs? Subsequently, how might these changes influence the overall ribosome structure?
To address these questions, we provide here a comprehensive comparison of homologous proteins from bacterial and eukaryotic ribosomes and revise available data about the NLSs in ribosomal proteins. Firstly, we show that NLSs emerged in conserved ribosomal proteins via remodelling of their RNA-binding domains. Surprisingly, this remodelling occurred mainly in the highly conserved interior of the ribosome. In the interior, NLSs form extensive and selective interactions with ribosomal RNA (rRNA), binding predominantly to single-stranded helical junctions, which point to a possible role of the NLSs in rRNA folding. Finally, we use these structural observations to identify novel NLSs in human ribosomal proteins uS12 and uL24.
Results
NLSs altered RNA-binding domains of the conserved proteins
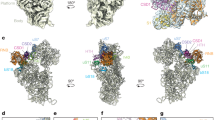
To gain an insight into the evolutionary origin of the NLSs in ribosomal proteins, we first mapped previously identified NLSs in the crystal structure of the eukaryotic ribosome from budding yeast Saccharomyces cerevisiae (S. cerevisiae), analysed their structure, and compared them to the corresponding segments in homologous proteins from the Escherichia coli (E. coli) ribosome (Methods). In total, we analysed twelve NLSs from ten conserved ribosomal proteins (Supplementary Table 1)2,5,6,7,8,9,10,11,12,13. We found both that all the NLSs of ribosomal proteins reside within non-globular extensions of rRNA-binding domains and that these NLS-carrying extensions have different structures in eukaryotes and in bacteria. For instance, NLSs of eukaryotic proteins uS3, uS4, uL13, uL15 and uL18 reside within the extensions that overlap with those of bacterial proteins, but adopt different secondary and tertiary structures (Fig. 1a, Supplementary Fig. 1). This finding was surprising, both because these extensions have similar size and charge in bacteria and eukaryotes and were previously assigned as conserved, according to sequence alignments14,15,16. Other NLSs reside within rRNA-binding extensions that are absent in bacterial proteins – as sequence alignments had shown for proteins uS8, uL3 (ref. 2), uL18 (ref. 6), uL23 (ref. 13) and uL29 (ref. 7; (Fig. 1a, Supplementary Fig. 1). Taken together, this comparison illustrated that, despite high content of basic residues in ribosomal proteins, particularly at their rRNA-binding interface, the NLSs or similar motifs are absent in bacteria and apparently emerged via remodelling of the rRNA-binding domains of conserved ribosomal proteins.

(a) Crystal structures of four pairs of homologous proteins from 70S E. coli and 80S S. cerevisiae ribosomes: proteins are coloured according to the secondary structure, with red colour and red arrows pointing to NLSs of eukaryotic proteins (top panels) and to corresponding positions in bacterial homologues (bottom panels). NLSs reside within non-globular extensions of eukaryotic proteins with substantially remodelled secondary and tertiary structure compared with analogous protein segments in bacterial ribosomal proteins. (b) Fragments of the ribosome interior with a zoom on interactions between NLSs and rRNA within the eukaryotic ribosome (top panels) and corresponding segments of bacterial ribosome structure (bottom panels); nucleotides, which contact ribosomal proteins and ions/water molecules (shown as spheres), are in blue; labels correspond to 23S/25S rRNA helices. When ribosomal proteins are incorporated into the ribosome, NLSs are buried in the rRNA: compared with bacterial ribosomes, NLSs structurally replace non-globular extensions of bacterial proteins or magnesium ions/water in the ribosome interior and form similar stabilizing contacts with single-stranded helical junctions of conserved rRNA, suggesting a role of NLSs in rRNA folding.
NLSs maintain conserved folds of the rRNA
To understand how the NLSs were accommodated in the conserved core of the ribosome, we analysed their surroundings and interactions within the ribosome interior. Compared with bacterial ribosomes, the NLSs structurally replace extensions of homologous proteins (uS3, uS8, uL13, uL15 and uL18) or magnesium ions and water (uL3, uL23, uL29 and uS2) and form extensive contacts with rRNA (Fig. 1b, Supplementary Fig. 1). In total, they establish ∼260 salt bridges, hydrogen bonds and stacking interactions. Remarkably, interactions between the rRNA and NLSs have two common tendencies. Firstly, although ribosomal proteins form most of their contacts with rRNA helices, which are thought to play a role in the recruitment of ribosomal proteins to the nascent rRNA transcript17, the NLSs bind predominantly to single-stranded rRNA. Thus, ∼96% of their contacts are established with helical junctions, internal loops, or helix–loop borders (Fig. 1b, Supplementary Table 1). Since the structure of single-stranded rRNA segments governs three-dimensional packing of RNA helices, these contacts suggest that NLSs may participate in the rRNA folding. Secondly and most unexpectedly, we found that the NLSs bind predominantly to highly conserved rRNA, whose structure is nearly invariant in both eukaryotes and bacteria, as exemplified by proteins uL3, uL13, uL15 and uL18 (Fig. 1b, Supplementary Fig. 1). This observation challenges the current view that conserved components of the ribosome are mainly divergent at the cytoplasm-exposed surface14,15,16, and illustrates that substantial changes in conserved ribosomal proteins are also present between the universally conserved ribosomal cores of eukaryotes and bacteria.
Structure-guided search for novel NLSs in ribosomal proteins
Having elucidated previously unknown consensus structural features of NLSs of ribosomal proteins, we next endeavoured to use our structural observations to uncover unknown NLSs. For this purpose, we examined how many homologous proteins have different folds of their rRNA-binding domains within similar rRNA cavities of bacterial and eukaryotic ribosomes. We preformed structural comparison of all homologous proteins from eukaryotic (S. cerevisiae) and bacterial (E. coli) ribosomes and, where necessary, analysed surrounding of ribosomal proteins in the ribosome structure (Supplementary Online Methods). This approach allowed us to resolve ambiguity between previous sequence15,16 and secondary structure alignments14 and correct numerous mistakes in the correspondence between the residues of uS3, uS4, uS11, uS12, uS13, uS15, uS19, uL2, uL3, uL4, uL5, uL9, uL13, uL15 and uL18 (Supplementary Fig. 2). In total, we found that 27 out of 32 conserved proteins possess differently folded segments within their rRNA-binding domains between bacteria and eukaryotes, despite overall conserved structure of the adjacent rRNA: in fifteen proteins, seemingly conserved extensions have different fold in bacteria and eukaryotes, and, in seventeen proteins, extensions differ in size despite the high conservation of the surrounding rRNA (Supplementary Fig. 2). To verify if these structural differences could indeed point to the location of the NLSs, we selected proteins uS12 and uL24, whose rRNA-binding domains are among the most divergent in the small and the large ribosomal subunits, respectively (Fig. 2a, Supplementary Fig. 3). Next, we expressed uS12 fused to enhanced green fluorescent protein (eGFP) in human cells. Unlike a control eGFP sample (Fig. 2b), human uS12–eGFP fusion accumulated in the nucleoli, indicating the presence of NLS (Fig. 2c, Supplementary Fig. 4a). By contrast, the E. coli uS12–eGFP fusion was distributed between the nucleus and the cytoplasm and largely excluded from the nucleoli, consistent with the absence of NLSs in bacteria (Fig. 2d, Supplementary Fig. 4b). Next, we replaced the N terminus of E. coli uS12 with the N terminus of human uS12, a structure that is highly divergent between the two organisms despite high conservation of rRNA (Fig. 2a). We found that the resulting chimeric protein gained nucleolar accumulation in human cell lines HEK293 (Fig. 2e, Supplementary Fig. 4c). Subsequently, when the extension of human uS12 was replaced by the N terminus of its bacterial homologue, the resulting protein was no longer able to accumulate in the nucleolus (Fig. 2f, Supplementary Fig. 4d). Finally, upon examining isolated individual domains we showed that the N terminus of human uS12 alone is sufficient for the nucleolar accumulation of eGFP (Fig. 2g), whereas the globular domain alone is insufficient (Fig. 2h), indicating that the N-terminal segment indeed carries the NLS activity. Similarly, we identified the NLSs of ribosomal protein uL24, residing within structurally diverged N- and C-termini of this protein (Supplementary Fig. 3).

Exemplified by identification of NLS in human ribosomal protein uS12. (a) Structure of protein uS12, coloured according to structural conservation: conserved fold is shown in grey and bacteria- and eukaryote-specific in blue and red, respectively; surrounding rRNA is shown schematically with labels indicating 16S/18S rRNA helices. Protein uS12 is one of the 15 ribosomal proteins in which extensions have different folds in bacteria and eukaryotes, despite being bound to nearly identical rRNA cavities of bacterial and eukaryotic ribosomes. (b–h) eGFP fluorescence (top panels) and phase-contrast (bottom panels) snapshots of human cell line HEK293, which express eGFP fusions of human or E. coli protein uS12. Arrows point to nucleoli. All scale bars represent 10 μm. For each sample, eGFP localization was examined in 200 cells. Cells, in which eGFP distribution pattern was common for >95% of the analysed population, were used for imaging. The experiments were replicated three times. (b) eGFP alone (a negative control). (c) Human uS12–eGFP fusion. (d) E. coli uS12–eGFP fusion. (e) A protein hybrid carrying the N terminus of human uS12 and the globular domain of E. coli uS12. (f) A protein hybrid carrying the N terminus of E. coli uS12 and the globular domain of human uS12. (b–h) Identifying NLSs within human uS12 shows that its N-terminal extension (N-H. sapiens) carries NLS activity – by contrast to analogous extension in bacterial uS12 (N-E. coli).
Discussion
Collectively, our analysis revealed that the NLSs of conserved ribosomal proteins reside within highly diverged rRNA-binding domains and have extensive contacts with the rRNA. These contacts suggest that having evolved NLSs at the interface with conserved rRNA allows to use the NLSs to not only promote protein trafficking, but also to facilitate rRNA folding during ribosome biogenesis, thereby coordinating delivery of ribosomal proteins to the nascent rRNA with the rRNA folding. This notion is indirectly supported by recent studies of the ribosome biogenesis in bacteria and yeast18,19,20. For instance in E. coli, deletion of the N-terminal extension of protein uS12 causes defects of the 16S rRNA architecture and provokes lethality18. Since eukaryotic uS12 forms similar contacts with the 18S rRNA, it is possible that its N-terminal and NLS-carrying extension could also promote a proper folding of the 18S rRNA. In another case study in yeast, deletion of protein uL24 was recently shown to cause abnormal folds of the 25S rRNA19. Since the contacts between uL24 and rRNA are mediated largely by the NLS-carrying N- and C-terminal extensions of uL24, it is possible that the NLSs of this protein may carry a dual function by promoting a proper folding of rRNA.
Beyond insights into the ribosome biogenesis, our observations suggest how the nuclear localization signals could have emerged in the course of evolution. Of note, there are only 61 universally abundant proteins, which are present in all forms of life, among which 32 are conserved ribosomal proteins21. Therefore, the future extension of our analysis to a broad range of organisms, including distant eukaryotic and archaeal species, and systematic screening of their NLSs and NLS-binding partners may bring us closer to understanding how cells developed the nucleocytoplasmic transport system and may potentially provide new tools to control the essential process of ribosome biogenesis in all forms of life.
Methods
Comparison of bacterial and eukaryotic ribosomal proteins
Structures of 32 pairs of homologous proteins were derived from E. coli 70S ribosome (pdbs 4wao-p) and S. cerevisiae 80S ribosome (pdbs 3u5b-d) structures, and were initially aligned by the automated flexible FatCat fitting22. The alignments were corrected by comparison of a conformation of a polypeptide chain relative to its surrounding within the ribosome interior. By contrast to previous attempts to comprehensively compare bacterial and eukaryotic proteins – by us14 and others15,16 – we identified numerous extensions with unique structure in bacteria and eukaryotes that were mistakenly annotated as conserved due to their similar size in different species and lack of secondary structure and conserved sequence. These extensions were found in proteins uS3, uS4, uS11, uS12, uS13, uS15, uS19, uL2, uL3, uL4, uL5, uL9, uL13, ul15 and uL18 (Supplementary Fig. 2). NLSs of ribosomal proteins were mapped on the S. cerevisiae ribosome according to the original studies summarized in (Supplementary Table 1).
Cloning and constructing of protein mutants and chimeras
Genes, coding for Homo sapiens and E. coli, uS12 and uL24, were amplified from HeLa S3 (American Type Culture Collection (ATCC)) cDNA library or from a colony of JM109 E. coli strain and were later used for cloning or constructing chimeric genes. The primers used in this study were: to clone H. sapiens uS12 (residues 1–143) – GCGCCTCGAGATGGGCAAGTGTCGTGGA and GCGCGGATCCCCTTATGATCTTGGTCTTTCCTTC ; the N terminus of H. sapiens uS12 (residues 1–41) – GCGCCTCGAGATGGGCAAGTGTCGTGGA and GCGCGGATCCCCAGGGTTGGCCTTTAGGGCTG ; the globular domain of H. sapiens uS12 (residues 42–143) – GCGCCTCGAGATGTTTGGAGGTGCTTCTCATGC and GCGCGGATCCCCTTATGATCTTGGTCTTTCCTTC ; E. coli uS12 (residues 1–124) – GCGCCTCGAGATGGCAACAGTTAACCAGCT and GCGCGGATCCCCAGCCTTAGGACGCTTCACGC ; the hybrid protein comprising the N terminus of H. sapiens uS12 (residues 1–41) and the globular domain of E. coli uS12 (residues 24–124) – GCGCCTCGAGATGGGCAAGTGTCGTGGA , ATGCTTCCAGAGGGTTGGCCTTTAGGGCTG , GGCCAACCCTCTGGAAGCATGCCCGCAAAA and GCGCGGATCCCCAGCCTTAGGACGCTTCACGC ; the hybrid protein comprising the N terminus of E. coli uS12 (1–23) and the globular domain of H. sapiens uS12 (residues 42–143) – GCGCCTCGAGATGGCAACAGTTAACCAGCT , CACCTCCAAACGCAGGCACGTTGCTTTTCG , CGTGCCTGCGTTTGGAGGTGCTTCTCATGCA and GCGCGGATCCCCTTATGATCTTGGTCTTTCCTTC ; H. sapiens uL24 (residues 1–127) – GCGCCTCGAGATGAAGTTTAATCCCTTTGT and GCGCGGATCCCCTTCCTGCATCTTCTCAATGG ; the N terminus of H. sapiens (residues 1–49) – GCGCCTCGAGATGAAGTTTAATCCCTTTGT and GCGCGGATCCCCGATGGGCATGGATCGCACGT ; the globular domain of H. sapiens uL24 globular domain (residues 50–107) – GCGCCTCGAGCGAAAGGATGATGAAGTTCA and GCGCGGATCCCCGCGAGATTTGGCTTTCCGT ; and the C terminus of H. sapiens uL24 (residues 108–127) – GCGCCTCGAGCAAGTAGGAAAGGAAAAGGG and GCGCGGATCCCCTTCCTGCATCTTCTCAATGG . All the PCR products were cloned into pEGFP-N1 vector (Promega) between KpnI and XhoI sites. Their sequence was verified via sequencing analysis (GATC Biotech).
Cell lines, transfection and microscopy
Nucleolar accumulation of ribosomal proteins was examined in human cell line HEK293T (ATCC). In brief, cells were plated on 35 mm glass-bottom dishes (MatTek) at 25–40% confluence and in 24 h were transfected with FuGene 6 (Promega), according to the manufacturers’ protocol. Confocal fluorescent images were obtained by a Zeiss LSM510NLO scanning microscope: 8–12 h after transfection cells were rapidly transferred on a microscope stage, and the eGFP-localization pattern was examined in 200 cells. Cells, which eGFP-localization pattern was common for at least 95% of the analysed population, were used for imaging. For each genetic construct, experiments were repeated at least two times.
Additional information
How to cite this article: Melnikov, S. et al. Insights into the origin of the nuclear localization signals in conserved ribosomal proteins. Nat. Commun. 6:7382 doi: 10.1038/ncomms8382 (2015).
References
Melese, T. & Xue, Z. The nucleolus: an organelle formed by the act of building a ribosome. Curr. Opin. Cell Biol. 7, 319–324 (1995).
Moreland, R. B., Nam, H. G., Hereford, L. M. & Fried, H. M. Identification of a nuclear localization signal of a yeast ribosomal protein. Proc. Natl Acad. Sci. USA 82, 6561–6565 (1985).
Wool, I. G., Chan, Y. L. & Gluck, A. Structure and evolution of mammalian ribosomal proteins. Biochem. Cell Biol. 73, 933–947 (1995).
Rout, M. P., Blobel, G. & Aitchison, J. D. A distinct nuclear import pathway used by ribosomal proteins. Cell 89, 715–725 (1997).
Underwood, M. R. & Fried, H. M. Characterization of nuclear localizing sequences derived from yeast ribosomal protein L29. EMBO J. 9, 91–99 (1990).
Rosorius, O. et al. Human ribosomal protein L5 contains defined nuclear localization and export signals. J. Biol.Chem. 275, 12061–12068 (2000).
Chen, I. J., Wang, I. A., Tai, L. R. & Lin, A. The role of expansion segment of human ribosomal protein L35 in nuclear entry, translation activity, and endoplasmic reticulum docking. Biochem. Cell Biol. 86, 271–277 (2008).
Koch, B. et al. Yar1 protects the ribosomal protein Rps3 from aggregation. J. Biol. Chem. 287, 21806–21815 (2012).
Lindstrom, M. S. Elucidation of motifs in ribosomal protein S9 that mediate its nucleolar localization and binding to NPM1/nucleophosmin. PloS ONE 7, e52476 (2012).
Timmers, A. C., Stuger, R., Schaap, P. J., van 't Riet, J. & Raue, H. A. Nuclear and nucleolar localization of Saccharomyces cerevisiae ribosomal proteins S22 and S25. FEBS Lett. 452, 335–340 (1999).
Antoine, M. et al. Identification of an unconventional nuclear localization signal in human ribosomal protein S2. Biochem. Biophys. Res. Commun. 335, 146–153 (2005).
Das, P. et al. Insights into the mechanism of ribosomal incorporation of mammalian L13a protein during ribosome biogenesis. Mol. Cell. Biol. 33, 2829–2842 (2013).
Schaap, P. J., van't Riet, J., Woldringh, C. L. & Raue, H. A. Identification and functional analysis of the nuclear localization signals of ribosomal protein L25 from Saccharomyces cerevisiae. J. Mol. Biol. 221, 225–237 (1991).
Ben-Shem, A. et al. The structure of the eukaryotic ribosome at 3.0 A resolution. Science 334, 1524–1529 (2011).
Klinge, S., Voigts-Hoffmann, F., Leibundgut, M., Arpagaus, S. & Ban, N. Crystal structure of the eukaryotic 60S ribosomal subunit in complex with initiation factor 6. Science 334, 941–948 (2011).
Rabl, J., Leibundgut, M., Ataide, S. F., Haag, A. & Ban, N. Crystal structure of the eukaryotic 40S ribosomal subunit in complex with initiation factor 1. Science 331, 730–736 (2011).
Klein, D. J., Moore, P. B. & Steitz, T. A. The roles of ribosomal proteins in the structure assembly, and evolution of the large ribosomal subunit. J. Mol. Biol. 340, 141–177 (2004).
Calidas, D., Lyon, H. & Culver, G. M. The N-terminal extension of S12 influences small ribosomal subunit assembly in Escherichia coli. RNA. 20, 321–330 (2014).
Talkish, J., May, G., Lin, Y., Woolford, J. L. Jr. & McManus, C. J. Mod-seq: high-throughput sequencing for chemical probing of RNA structure. RNA. 20, 713–720 (2014).
Gamalinda, M. & Woolford, J. L. Jr. Deletion of L4 domains reveals insights into the importance of ribosomal protein extensions in eukaryotic ribosome assembly. RNA. 20, 1725–1731 (2014).
Koonin, E. V. Comparative genomics, minimal gene-sets and the last universal common ancestor. Nat. Rev. Microbiol. 1, 127–136 (2003).
Ye, Y. & Godzik, A. Flexible structure alignment by chaining aligned fragment pairs allowing twists. Bioinformatics. 19, ii246–ii255 (2003).
Acknowledgements
We are grateful to Mike Rout (The Rockefeller University, USA) and Peter Moore (Yale University, USA) for critical and inspirational feedback, Marcel Boeglin and David Bruchlen (IGBMC, France) for help with cell cultures and microscopy, Corwin Miller and Tonnie van den Elzen (Yale University, USA) for discussions and critical proofreading. This study was supported by SPF20111223404 FRM fellowship and ASTF 434-2014 EMBO fellowship (both to S.M.), the French National Research Agency grant ANR-11-BSV8-006 01 (to G.Y.), the ERC Advanced grant 294312, the HFSP grant RGP0062/2012 and the Russian Government Program of Competitive Growth of Kazan Federal University (all to M.Y.).
Author information
Authors and Affiliations
Contributions
S.M. and A.B-S. devised experiments, S.M. performed cloning and microscopy studies, S.M. and A.B-S. analysed the data and S.M., A.B-S., G.Y. and M.Y. wrote the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Supplementary Information
Supplementary Figures 1-4, Supplementary Table 1 and Supplementary References. (PDF 1735 kb)
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Melnikov, S., Ben-Shem, A., Yusupova, G. et al. Insights into the origin of the nuclear localization signals in conserved ribosomal proteins. Nat Commun 6, 7382 (2015). https://doi.org/10.1038/ncomms8382
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/ncomms8382
This article is cited by
-
Nuclear import of dimerized ribosomal protein Rps3 in complex with its chaperone Yar1
Scientific Reports (2016)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
