
Abstract
Cadmium is a transition metal ion that is highly toxic in biological systems. Although relatively rare in the Earth’s crust, anthropogenic release of cadmium since industrialization has increased biogeochemical cycling and the abundance of the ion in the biosphere. Despite this, the molecular basis of its toxicity remains unclear. Here we combine metal-accumulation assays, high-resolution structural data and biochemical analyses to show that cadmium toxicity, in Streptococcus pneumoniae, occurs via perturbation of first row transition metal ion homeostasis. We show that cadmium uptake reduces the millimolar cellular accumulation of manganese and zinc, and thereby increases sensitivity to oxidative stress. Despite this, high cellular concentrations of cadmium (~17 mM) are tolerated, with negligible impact on growth or sensitivity to oxidative stress, when manganese and glutathione are abundant. Collectively, this work provides insight into the molecular basis of cadmium toxicity in prokaryotes, and the connection between cadmium accumulation and oxidative stress.
Similar content being viewed by others

Introduction
Global cadmium (Cd) production has risen by >1,000-fold since the beginning of the twentieth century to ~20,000 tons per year1. Correspondingly, anthropogenic release of Cd into the atmosphere now significantly outstrips natural fluxes and predominantly occurs via non-ferrous ore processing, combustion of fossil fuels, and manufacturing, use and disposal of Cd-containing products1. As Cd cannot be degraded, its flux into marine and terrestrial ecosystems has increased the risk of exposure as it accumulates in the food chain, and it is estimated that humans ingest 30 μg of Cd every day1,2. Cd, which occurs as the divalent cation in the natural environment, is acutely toxic to all forms of life, although there are exceptions to this rule for organisms that have evolved in environments with extremely low zinc abundance3. Cd toxicity in humans and microbes is, at a cellular level, closely associated with oxidative stress, despite the inability of Cd2+ to directly produce reactive oxygen species4. As a consequence, understanding the molecular basis for how Cd2+ causes toxicity is a crucial issue.
Cd2+ accumulation in microbes and humans primarily occurs by uptake via divalent metal transporters, such as manganese (Mn2+) transporters, although the molecular basis for this process remains unknown5,6. We investigated Cd2+ toxicity in the Gram-positive bacterium Streptococcus pneumoniae (pneumococcus) as it has only a single Mn2+-specific uptake pathway. The pneumococcus acquires Mn2+ via the PsaBCA permease, which comprises the ATP-binding cassette (ABC) transporter, PsaBC, and a cell-surface solute-binding protein (SBP), PsaA. The Psa permease is essential for both Mn2+ uptake and in vivo virulence7,8,9. Similar to other ABC permeases, the SBP PsaA defines the specificity of the uptake pathway. However, our recent studies showed that the functional specificity of the Psa permease in Mn2+ acquisition arose not from the specificity of Mn2+ binding by PsaA, but instead resulted from the concerted action of protein metal ion coordination chemistry and structural rearrangements that prevented release of other divalent transition metal ions, such as Zn2+, into the permease, despite their ability to bind to PsaA8. Thus, although it is conceivable that Cd2+ may interact with the Psa permease, whether it causes injury to the cell by non-productively competing for metal ion uptake or by accumulation in the cytosol remains unclear.
Here we use a combination of in vitro, in vivo and structural analyses to investigate how Cd2+ impacts transition metal homeostasis in S. pneumoniae. We show that S. pneumoniae accumulates millimolar concentrations of transition metal ions, and that Cd2+ dysregulates metal ion homeostasis by perturbing the Mn2+ and Zn2+ uptake and efflux pathways, although via strikingly different mechanisms. Manganese accumulation is most affected, and this arises from Cd2+ competing with Mn2+ ions for uptake via the Psa permease and the concomitant upregulation of the Mn2+-efflux pathway. High-resolution structural analyses reveal that although the Mn2+-specific Psa permease uses coordination chemistry and structural features to prevent the mistranslocation of other ions, such as Zn2+, the larger ionic radius of Cd2+ enables this metal to circumvent these regulatory mechanisms. The increased sensitivity to oxidative stress associated with Cd2+ toxicity is due to impaired Mn2+ acquisition and not directly due to the intracellular accumulation of Cd2+ ions, which are buffered on cellular glutathione. Collectively, these findings highlight the challenge that biological systems encounter when non-physiological elements enter the biosphere. This study provides novel insights into the intracellular concentrations of transition metal ions in a Gram-positive cell, the mechanisms associated with their homeostasis, and how Cd2+ dysregulates these processes.
Results
S. pneumoniae accumulates high concentrations of metal ions
We first sought to determine the cellular quotient of first row transition metal ions in S. pneumoniae. Inductively coupled plasma–mass spectrometry (ICP–MS) data were combined with scanning electron microscopy analyses and colony-forming unit (c.f.u.) counts to derive the intracellular metal ion accumulation data in molar units. In this context, the intracellular metal ion concentration corresponds to the total accumulation of transition metal ions in the cytoplasmic compartment of S. pneumoniae and represents all cytoplasmic metal ions, from those bound in tight metal–protein complexes to those occurring as labile pools of exchangeable ions. Although the total number of metal ions per cell was similar to that previously reported for Escherichia coli10, the pneumococcus has a smaller mean (±s.d.) cell volume, 0.113±0.04 fl (n=44; Table 1), and only a single cellular compartment, which results in the total cellular metal ion concentrations being in the millimolar range (Fig. 1a). The Irving–Williams stability series is the near-universal order of metal–protein-binding affinity, which arises from the imperfect steric selection of metal ions that occurs within protein–metal-binding sites. In order of increasing atomic number this can be represented as Mn2+<Fe2+<Co2+<Ni2+<Cu2+>Zn2+ (ref. 11). Our data also revealed that the total cellular quotient of Mn2+ (8.6±0.5 mM) was higher than the accumulation of other more competitive ions, such as Zn2+ (6.8±0.4 mM). Hence, in S. pneumoniae, the accumulation of Mn2+ to higher intracellular concentrations may be one factor that aids in increasing its bioavailability, relative to more competitive ions, such as Zn2+, and thereby aid in its acquisition by proteins that require Mn2+. We then proceeded to ascertain the impact of Cd2+ on the cellular accumulation of transition metal ions.

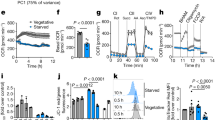
(a) Total cellular metal ion accumulation of S. pneumoniae represented as the mean (±s.e.m.) concentration of ions per cell (determined as c.f.u.) of known cell volume (determined by scanning electron microscopy (SEM)) grown in the specified conditions. The values represent the total cellular metal ion quota, that is, bound, buffered and unbound metal ions, and are from at least three independent biological experiments. Metal ions assessed were Mn2+, Fe2+/3+, Co2+, Ni2+, Cu1+/2+, Zn2+ and Cd2+. The statistical significance of the differences in the mean metal concentrations was determined by a one-way ANOVA (ND, not detected; NS, not significant and ****P<0.0001). (b,c) S. pneumoniae (D39) grown in CDM supplemented with metal ions as indicated. The data correspond to mean (±s.e.m.) absorbance at 600 nm measurements from three independent biological experiments. Errors bars, where not visible, are overlapped by the representative symbols. (d) Relative transcription, corrected to 16S rRNA, of psaA by S. pneumoniae when grown in CDM supplemented with 1 μM Mn2+ (black), 1 μM Mn2+: 30 μM Cd2+ (light grey) and 30 μM Mn2+: 30 μM Cd2+ (dark grey). (e) Quantitative immunoblot data of PsaA (Supplementary Fig. 2), from equivalent total protein loadings of whole-cell extracts, from S. pneumoniae when grown in CDM supplemented with 1 μM Mn2+ (black), 1 μM Mn2+: 30 μM Cd2+ (light grey) and 30 μM Mn2+: 30 μM Cd2+ (dark grey). (f) Relative transcription, corrected to 16S rRNA, of czcD by S. pneumoniae when grown in CDM supplemented with 1 μM Mn2+ (black), 1 μM Mn2+: 30 μM Cd2+ (light grey) and 30 μM Mn2+: 30 μM Cd2+ (dark grey). The data correspond to the mean (±s.e.m.) of three independent biological experiments. The statistical significance of the differences in the mean data was determined by two-tailed unpaired t-tests (*P<0.05, **P<0.01 and ***P<0.001).
Cd dysregulates transition metal homeostasis
Titrations of Cd2+ into a cation-defined medium (CDM), with a constant concentration of Mn2+ (1 μM), showed that pneumococcal growth was perturbed (Fig. 1b and Supplementary Fig. 1a), but that this growth defect could essentially be abrogated by the addition of a stoichiometric concentration of Mn2+ to the Cd2+-supplemented CDM (Fig. 1c and Supplementary Fig. 1b). S. pneumoniae growth in the presence of 30 μM Cd2+ reduced the cellular concentrations of Mn2+ and Zn2+. Manganese accumulation was most affected, with a reduction of ~90% (Fig. 1a; 1 μM Mn2+: 30 μM Cd2+, P<0.0001 (one-way analysis of variance (ANOVA))), despite no significant effect on cell volume (Table 1; 1 μM Mn2+: 30 μM Cd2+, P=0.376 (one-way ANOVA)). Supplementation with Mn2+ restored pneumococcal Mn2+ accumulation to ~60% of the unchallenged levels (Fig. 1a and Supplementary Table 1; 30 μM Mn2+: 30 μM Cd2+, P<0.0001 (one-way ANOVA)). Zinc also showed a significant decrease in accumulation in the presence of Cd2+ (Fig. 1a; 1 μM Mn2+: 30 μM Cd2+, P<0.0001; 30 μM Mn2+: 30 μM Cd2+, P<0.0001 (one-way ANOVA)). Cd2+ accumulation increased in the presence of 30 μM Cd2+, with or without Mn2+ (Fig. 1a). The degree to which Cd2+ was accumulated was more than that of any other transition metal ion in the pneumococcus, despite having no physiological role. At 1 μM Mn2+: 30 μM Cd2+ in the growth medium, intracellular Cd2+ concentration was 46.4±2.9 mM. Cd2+ accumulation was reduced by nearly 70% on the addition of 30 μM Mn2+ to the growth medium (16.5±0.7 mM Cd2+), but cellular accumulation of the ion still remained more than that of the other first row transition metal ions.
We then analysed the impact of Cd2+ on transition metal ion homeostatic pathways. Transcriptional analyses revealed that psaA transcription increased by 8.4-fold (P=0.0007 (two-tailed unpaired t-test)) and PsaA expression by 2.8±0.3-fold (P<0.0001 (two-tailed unpaired t-test)) when grown in the presence of 30 μM Cd2+ (Fig. 1d,e), consistent with the Mn2+-responsive transcriptional regulator, PsaR, sensing the Mn2+ depletion7,12. Transcription of psaA remained elevated during growth supplemented with 30 μM Mn2+ albeit to a lesser, but still significant, extent (3.5-fold, P=0.0135 (two-tailed unpaired t-test)), although PsaA expression did not appear to be significantly increased (Fig. 1d,e). Zinc accumulation in S. pneumoniae also occurs via an ABC permease, the Adc permease9,13,14. Transcriptional analysis of the Zn2+-responsive regulator, adcR15, and the Zn2+-recruiting SBPs associated with the Adc permease, adcA and adcAII (refs 13, 14, 16), did not show significant changes in the presence of Cd2+ (Supplementary Fig. 3). Despite this, the Zn2+ accumulation levels (2.6±0.4 mM) were comparable to strains without a functional Adc permease, ΔadcCBA (2.7±0.3 mM) and ΔadcA/ΔadcAII (2.5±0.3 mM). The perturbation of Zn2+ acquisition was not due to competition for the Adc permease as the dominant Zn2+-recruiting SBP, AdcA, showed very poor interaction with Cd2+ by comparison with Zn2+ (Supplementary Fig. 3). Taken together, these findings implicate Cd2+ as being able to exert an effect on the Zn2+-responsive transcriptional regulators. S. pneumoniae uses two Zn2+-responsive transcriptional regulators: AdcR, which regulates the Zn2+-uptake machinery described above, and SczA, which regulates transcription of the cation diffusion facilitator efflux protein CzcD15,17. Consequently, we examined whether CzcD was also affected by Cd2+ abundance. Growth of S. pneumoniae in CDM with 1 μM Mn2+: 30 μM Cd2+ increased czcD transcription by ~70-fold (P=0.0011 (two-tailed unpaired t-test); Fig. 1f), while supplementation with Mn2+ (30 μM Mn2+: 30 μM Cd2+) reduced this to only ~12.7-fold more than cells grown in unsupplemented CDM (Fig. 1f). Although CzcD from Cupriavidus metallidurans is protective against Co2+, Zn2+ and Cd2+ ions18, there is no evidence to suggest that the S. pneumoniae orthologue has similar poly-specificity. Consequently, we examined an isogenic ΔczcD mutant strain to ascertain whether pneumococcal czcD contributed to Cd2+ resistance. The S. pneumoniae ΔczcD strain was hypersensitive to extracellular Zn2+, consistent with a role in Zn2+ homeostasis (Fig. 2). Furthermore, the Cd2+-induced efflux of Zn2+ observed in wild-type S. pneumoniae was abolished in the ΔczcD strain (ΔczcD 1 μM Mn2+, 4.4±0.3 mM Zn2+; ΔczcD 1 μM Mn2+: 30 μM Cd2+ 4.2±0.3 mM Zn2+; P=0.78 (two-tailed unpaired t-test)). However, although Cd2+ accumulation in the ΔczcD strain showed a mild elevation of ~1.4-fold compared with the wild-type, the growth phenotypes of the ΔczcD strain were indistinguishable from the wild-type strain and also showed no increase in susceptibility to extracellular Cd2+ (Fig. 2 and Supplementary Fig. 4). Taken together, these data indicate that, despite the upregulation of S. pneumoniae czcD in response to Cd2+ exposure, CzcD is primarily a Zn2+ efflux transporter and appears to serve, at best, only a minor protective role against Cd2+.

Drop test analysis of S. pneumoniae wild-type (D39), ΔczcD, ΔmntE, ΔgshT, ΔgshT/ΔczcD and ΔsodA on blood agar (BA) supplemented with: 0, 100, 300 and 1,000 μM ZnSO4 or 0, 10, 30 and 100 μM CdCl2. Cells were grown and adjusted to A600 of 0.4 and were serially diluted. Drops (5 μl) were spotted onto the BA plate, starting from the 100 dilution (top) down to the 10−5 dilution (bottom).
In addition to czcD, the pneumococcal genome encodes four other putative metal ion efflux pathways. These are as follows: spd1384 (mntE), a paralogue of CzcD involved in Mn2+ efflux19; spd1438 (cadD), a membrane protein of unknown function associated with Cd2+ tolerance20; two P-type ATPases, spd0635 (copA), which is responsible for Cu1+ efflux21; and spd1927, which has homology to pmtA, a PerR-regulated gene in S. pyogenes associated with Zn2+ resistance22. The mntE gene has previously been reported to be constitutively expressed and not responsive to cellular Mn2+ abundance19. Here, we observed that extracellular Cd2+ induced a 2.1-fold increase in mntE transcription (P=0.0211 (two-tailed unpaired t-test); Supplementary Fig. 5a). On supplementation with 30 μM Mn2+, mntE transcription was reduced to non-challenged levels (P=0.3665 (two-tailed unpaired t-test) Supplementary Fig. 5a), suggesting that intracellular Cd2+ concentrations modulated mntE expression via an unknown metal-responsive regulator. We then analysed an isogenic ΔmntE strain, which lacks the primary Mn2+-efflux pathway19, for sensitivity to Cd2+. Consistent with Cd2+ also perturbing Mn2+ homeostasis via MntE, the ΔmntE strain retained a higher cellular Mn2+ concentration (2.53±0.1 mM) than the wild-type (0.78±0.07 mM) (P<0.0001 (two-tailed unpaired t-test)), when grown in 1 μM Mn2+: 30 μM Cd2+, and showed greater resistance to extracellular Cd2+ (Fig. 2). The cadD gene showed a similar trend to czcD and mntE with a ~3.6-fold increase in transcription when challenged with 30 μM Cd2+ (P<0.0001 (two-tailed unpaired t-test); Supplementary Fig. 5b), followed by abrogation when supplemented with 30 μM Mn2+. However, the P-type ATPases, copA and spd1927, were transcriptionally unresponsive to Cd2+ (Supplementary Fig. 5c,d), suggesting that they were unlikely to contribute to Cd2+ management. Collectively, these data show that Cd2+ accumulation was associated with transcriptional activation of several metal ion membrane protein transporters. However, of those pathways with known efflux function, that is, CzcD and MntE, Cd2+ ions subverted their primary functional roles and, instead, further dysregulated transition metal ion homeostasis in S. pneumoniae via amplification of Zn2+ and Mn2+ efflux, respectively. The mechanism(s) of Cd2+ efflux in S. pneumoniae, if any, remains to be identified.
Cd2+ competes with Mn2+ for the Psa permease
To elucidate the hitherto unexplained molecular basis for Cd2+ uptake via Mn2+ transporters, we examined the interaction of Cd2+ with the Mn2+-recruiting SBP PsaA. Although the physiological role of PsaA is in recruiting Mn2+, the metal-binding site of the protein has previously been shown to interact with other divalent transition metal ions8. However, coordination chemistry at the metal-binding site and structural dynamics prevent release of these other ions, and thereby maintain the functional role of the Psa permease in Mn2+ uptake8. Here the interaction of Cd2+ with PsaA was analysed by determining the structure of wild-type PsaA with Cd2+ at 2.0 Å resolution (Fig. 3a and Supplementary Fig. 6). The PsaA–Cd2+ complex revealed that Cd2+ was coordinated by the metal-binding site residues His67, His139, Glu205 and Asp280 in a structure that was highly similar (root-mean-square deviation (RMSD)<0.3 Å over 242 Cα atoms) to our previously reported Mn2+- and Zn2+-bound structures23,24. Nevertheless, small re-orientations of the side chains of the metal-coordinating residues allowed the bulkier Cd2+ ion (ionic radius of 95 pm for Cd2+ versus 84 pm for Mn2+) to be accommodated into the protein–metal-binding site (Fig. 3b and Supplementary Table 2). In the PsaA–Cd2+ structure, the distances between metal-coordinating atoms and the metal ion were increased by ~0.2 Å compared with those for PsaA-Zn2+ (Supplementary Table 2). Similar increases in metal–protein distances have been observed for the Cd2+–thiolate-binding sites of Cd2+-substituted metallothionein and rubredoxin proteins, compared with the Zn2+-bound equivalents25,26. Although Cd2+ and Zn2+ often display similar coordination spheres in proteins, the longer bond distances observed for Cd2+ usually result in an increase in the coordination number. Accordingly, the coordination geometry in the PsaA–Cd2+ structure is an intermediate between a square-pyramid and trigonal-bipyramid (n=5), in contrast to the 4- and 6-coordinate sites observed for the Zn2+- and Mn2+-bound PsaA structures, respectively (Fig. 3d,e and Supplementary Table 3). A comparison of the observed coordination of PsaA–Cd2+ with other metal-binding sites of metalloprotein structures in the PDB, composed of mixed N/O ligating atoms, revealed that Cd2+ is found in a variety of coordination geometries with coordination numbers ranging from 5 to 7 (Supplementary Table 4). Nevertheless, the only bona fide protein–metal-binding site for Cd2+ is in carbonic anhydrase from Thalassiosira weissflogii, which has been shown to be a cambialistic enzyme with Zn2+ in a tetrahedral geometry and Cd2+ in a trigonal-bipyramidal geometry3,27,28. Thus, the coordination geometry found in PsaA–Cd2+ conforms to what has been observed for the only known naturally occurring Cd2+-binding protein, and for other protein structures deposited in the PDB. We further examined the interaction of metal-free (apo) PsaA with Cd2+ and observed that apo-PsaA bound Cd2+ in a 1:1 molar ratio (Fig. 3f), with a KD of 5.6±1 nM (Supplementary Fig. 7). As the cognate ligand, Mn2+, has a KD of 3.3±1 nM for PsaA, the relatively small (approximately twofold) difference in the KDs is consistent not only with the competitive effect on Mn2+ acquisition but also the continued, albeit reduced, Cd2+ accumulation during growth in the presence of equimolar concentrations of both ions (30 μM Mn2+: 30 μM Cd2+). Collectively, these findings are consistent with the observed impact of Cd2+ on Mn2+ accumulation in the wild-type and mutant S. pneumoniae strains and indicate that it is a competitive effect mediated via PsaA. However, they also indicate that interaction of Cd2+ with PsaA is distinct from the previously characterized Mn2+ and Zn2+ interactions.

(a) Cartoon illustration of Cd2+-bound wild-type PsaA in closed conformation (PDB accession code 4UTP). The structure consists of N-terminal and C-terminal (β/α)4 domains with a domain-linking helix. The metal-binding site is situated between the two domains, in which a Cd2+ ion (pale-yellow sphere) is bound. (b) The metal-binding site of wild-type PsaA in the presence (light blue, PDB accession code 4UTP) and absence (green, PDB accession code 3ZK7) of Cd2+ (pale-yellow sphere), with the metal coordination shown as dashed lines. (c) Superposition of Cd2+-bound (light blue, PDB accession code 4UTP) and apo (green, PDB accession code 3ZK7) wild-type PsaA structures (Cα atoms, residues 32–163, were used for the superposition). The linking helix is shown in cartoon representation, with other helices shown as cylinders. The crossing angle of the C-terminal (β/α)4 domains between the structures is marked. The bound Cd2+ ion is shown as a pale-yellow sphere. In the apo structure, backbone atoms in the flexible region of the linking helix (residues 184–194) are shown in stick representation, with hydrogen bonds between backbone atoms as dashed lines. (d) Comparison of the metal-binding sites between Cd2+-bound (light blue, PDB accession code 4UTP) and Zn2+-bound (purple, PDB accession code 1PSZ) wild-type PsaA structures. The metal ions are shown as spheres (Cd2+, pale yellow; Zn2+, grey) and their coordination as dashed lines. (e) Comparison of the metal-binding sites between Cd2+-bound (light blue, PDB accession code 4UTP) and Mn2+-bound (yellow, PDB accession code 3ZTT) wild-type PsaA structures. The metal ions are shown as spheres (Cd2+, pale yellow; Mn2+, blue) and their coordination as dashed lines. In a–e, the metal-coordinating residues are shown in stick representation. (f) In vitro metal binding of Cd2+ to wild-type or D280N variant apo-PsaA analysed by ICP–MS. Data correspond to the mean (±s.d.) molar ratio of Cd2+ to the PsaA isoform for apo-PsaA (black), Cd2+-PsaA (light grey), EDTA-Cd2+-PsaA (dark grey). apo-D280N-PsaA (black), Cd2+-D280N-PsaA (grey) and EDTA-Cd2+-D280N-PsaA (white).
PsaA is permissive for Cd2+ binding and release
PsaA uses a ‘spring-hammer’-like mechanism for metal ion binding, in which the dynamics of the metal ion-loaded protein are dictated by a combination of metal ion geometry and the distortion of a helix linking the N- and C-terminal lobes of the protein8. We further analysed the metal-binding site coordination geometry by use of an Asp280 variant isoform (PsaA-D280N), as it would be incapable of facilitating the trigonal-bipyramidal coordination observed in wild-type PsaA8. The structure of PsaA-D280N bound to Cd2+, refined to 1.7 Å resolution, revealed a partially solvated Cd ion loosely interacting with residue Glu205 (Fig. 4a,b). The C-terminal lobe of PsaA showed minor movement (~1.0° relative to the open, metal-free conformation), but the inability of Cd2+ to ligate to residue Asp280 prevented further conformational changes. Consistent with these observations, PsaA-D280N was greatly impaired in Cd2+ binding (Fig. 3f).

(a) Superposition of Cd2+-bound PsaA-D280N (magenta, PDB accession code 4UTO) and apo wild-type (green, PDB accession code 3ZK7) PsaA structures (Cα atoms, residues 32–163, were used for the superposition). The bound Cd2+ ion is shown as a pale-yellow sphere. The linking helix is shown in cartoon representation (up to residue 183) and backbone atoms in the flexible region of the linking helix (residues 184–194) are shown in stick representation, with hydrogen bonds between backbone atoms as dashed lines. (b) Close-up view of the metal-binding sites between the superimposed Cd2+-bound PsaA-D280N (magenta, PDB accession code 4UTO) and apo wild-type (green, PDB accession code 3ZK7) PsaA structures. The bound Cd2+ ion is shown as a pale-yellow sphere and its coordination with the protein residues is shown as dashed lines. The water molecule coordinating the metal is shown as a light-blue sphere. In a,b, the metal-coordinating residues are shown in stick representation.
We then assessed the distortion of the linking helix by monitoring the number of disrupted backbone hydrogen bonds in its flexible region (residues 184–194) in wild-type PsaA–Cd2+ (Figs 3c and 4a, and Supplementary Table 5)8. Both the hinge-bending angle between the N- and C-terminal lobes and the number of main-chain hydrogen bonds disrupted in the residue range 184–194 corresponded more closely to the Mn2+-bound structure than to the ‘locked’ Zn2+-structure. This indicated that Cd2+ binding was a reversible process (Supplementary Table 5), consistent with the observation that Cd2+ could be extracted (~40%) from the PsaA–Cd2+ complex by using the chelating agent EDTA (Fig. 3f). Thus, these findings provide further support for the conclusion that both Mn2+ and Cd2+ compete for transport via the Psa permease, consistent with the cellular accumulation data, which shows that Mn2+ uptake occurs at the expense of Cd2+ import and vice versa.
Reduced Mn2+ levels result in sensitivity to oxidative stress
We then sought to assess how Cd2+ affected oxidative stress management in S. pneumoniae. Manganese has a prominent role in regulating the expression of superoxide dismutase (sodA), where it also serves as a cofactor29,30,31. Transcription of sodA was decreased under Cd2+-induced Mn2+ starvation, but this was restored when supplemented with Mn2+ (Fig. 5a), and direct measurement of SodA activity from S. pneumoniae showed a similar trend (Fig. 5b). The impact of Cd2+ on oxidative stress response was ascertained using paraquat, which causes oxidative damage by promoting a futile redox cycle in the cytoplasm. We observed that although growth in the presence of Cd2+ resulted in a significant decrease in survival (Fig. 5c, P=0.0087 (two-tailed unpaired t-test)), the increased sensitivity was not due to Cd2+, but instead was a result of the decreased Mn2+ accumulation, as S. pneumoniae grown in 30 μM Mn2+: 30 μM Cd2+ exhibited wild-type levels of survival (Fig. 5c). Hence, the perturbation of Mn2+ homeostasis by Cd2+ heightens the sensitivity to oxidative stress, while Cd2+ could be accumulated to high intracellular concentrations with no apparent deleterious effect. This ability to accumulate Cd2+ without direct toxicity indicated that intracellular buffering of the ion was crucial.

(a) Relative transcription, corrected to 16S rRNA, of sodA by S. pneumoniae when grown in CDM supplemented with 1 μM Mn2+ (black), 1 μM Mn2+: 30 μM Cd2+ (light grey) and 30 μM Mn2+: 30 μM Cd2+ (dark grey). The data correspond to the mean (±s.e.m.) of three independent biological experiments. (b) SodA activity assay S. pneumoniae when grown in CDM supplemented with 1 μM Mn2+ (black), 1 μM Mn2+: 30 μM Cd2+ (light grey) and 30 μM Mn2+: 30 μM Cd2+ (dark grey). Activity was calculated as a percentage of SodA activity observed for S. pneumoniae cells grown in 1 μM Mn2+. The data correspond to the mean (±s.e.m.) of three independent biological experiments. (c) Paraquat killing of S. pneumoniae grown in CDM supplemented with 1 μM Mn2+ (black), 1 μM Mn2+: 30 μM Cd2+ (light grey) and 30 μM Mn2+: 30 μM Cd2+ (dark grey). Survival was calculated as a percentage of colonies at 30 min compared with cells not challenged with paraquat. The data correspond to the mean (±s.e.m.) of three independent biological experiments. (d) Total glutathione determination per mean cell volume for S. pneumoniae grown in CDM supplemented with 1 μM Mn2+ (black), 1 μM Mn2+: 30 μM Cd2+ (light grey) and 30 μM Mn2+: 30 μM Cd2+ (dark grey). The statistical significance of the differences in the mean data for a–d was determined by two-tailed unpaired t-tests (NS, not significant; *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001). (e,f) S. pneumoniae ΔgshT (e) and ΔsodA (f) grown in CDM supplemented with metal ions as indicated. The data correspond to mean (±s.e.m.) absorbance 600 nm measurements from three independent biological experiments. Errors bars, where not visible, are overlapped by the representative symbols.
Reduced thiol groups on small peptides, primarily glutathione in S. pneumoniae, have been implicated in having an essential role in intracellular transition metal ion management (Fe2+ and Zn2+) and in ameliorating Cd2+ toxicity32. S. pneumoniae is incapable of de novo glutathione synthesis and acquires glutathione via a high-affinity ABC permease, the Gsh permease32. Here we investigated the contribution of glutathione to management of transition metal ion stress. In S. pneumoniae, the total glutathione content, in response to Cd2+ exposure, increased by approximately twofold to a mean (±s.e.m.) pneumococcal cell concentration of 19.0±0.9 mM (Fig. 5d). Glutathione abundance is crucial for Cd2+ management, as a mutant strain incapable of glutathione acquisition, S. pneumoniae ΔgshT, was hypersensitive to Cd2+ stress (1 μM Mn2+: 30 μM Cd2+; Figs 2 and 5e) and, in contrast to the wild type, could not be rescued by addition of Mn2+ (Fig. 5e; 30 μM Mn2+: 30 μM Cd2+). By contrast, the ΔsodA strain, which we have previously shown to be hypersensitive to oxidative stress challenge30,33, demonstrated a wild-type growth phenotype in the presence of Cd2+, with no apparent increase in sensitivity to metal ion stress (Figs 2 and 5f). Taken together, these findings indicate that glutathione serves a major role in Cd2+ buffering, whereas SodA appears to be dispensable, consistent with the lack of redox activity of Cd2+ ions.
To ascertain whether this was also the case for Zn2+, which is also presumed to be buffered by glutathione, we examined the response of the ΔgshT strain to Zn2+ exposure. Intriguingly, while the ΔgshT strain was insensitive to subinhibitory concentrations of Zn2+, similar to the ΔczcD strain, the combination mutant (ΔgshT/ΔczcD) showed significant attenuation of growth (100 μM Zn2+; Fig. 2). At higher concentrations of Zn2+, the growth of the ΔgshT/ΔczcD strain corresponded with the hypersensitive phenotype of the ΔczcD strain, while the ΔgshT strain showed a very mild impact, relative to wild type (300 μM Zn2+; Fig. 2). Taken together, these data show that intracellular management of Zn2+ occurs via the concerted actions of both glutathione and Zn2+ efflux, wherein the activation and efflux via CzcD is crucial to prevent Zn2+ intoxication. By contrast, management of Cd2+, which has no major efflux pathway, is predominantly dependent on cellular glutathione to prevent mismetallation of other proteins. Concordantly, when Cd2+ accumulation surpasses the buffering capacity of the glutathione pool (1 μM Mn2+: 30 μM Cd2+), or in the absence of intracellular glutathione (ΔgshT), this leads to Cd2+ toxicity.
Discussion
Anthropogenic-facilitated entry of non-physiological elements, such as Cd2+, into the biosphere presents unique cellular challenges for biological systems. The prior absence of these elements from the biosphere obviated any selective pressure towards evolving mechanisms for managing their impact on biological processes. As a consequence, many non-physiological elements rapidly accumulate in the food chain leading to significant toxicity in higher organisms1,2. We observed that Cd2+ was toxic to S. pneumoniae and that this arose from acute dysregulation of the transition metal ion homeostatic mechanisms. Elucidation of the total cellular quotient of metal ions in S. pneumoniae not only showed the impact of Cd2+ but also revealed novel aspects of cellular transition metal homeostasis. S. pneumoniae is a Gram-positive organism, comprising only a single cellular compartment; thus the concentration of transition metal ions accurately reflects their total intracellular accumulation and encompasses metal ions occurring in labile exchangeable pools and those bound weakly or tightly to proteins, peptides, nucleic acids and other molecules. Determination of the cellular quotient of metal ions in wild-type S. pneumoniae and mutant variants revealed the range over which cellular variation of transition metal ion concentrations is permissible. Manganese accumulation shows significant plasticity, with cells being viable at concentrations as low as ~3% of typical accumulation levels23,30. By contrast, Zn2+ accumulation is regulated in a much narrower window, with its minimal cellular quotient being between 20 and 40% of typical accumulation. Intriguingly, the higher total cellular quotient of Mn2+ ions suggests that the concentration ‘set-points’ for cellular accumulation of transition metal ions may also contribute to ensuring efficient acquisition of metal cofactors by proteins. Manganese, which has been reported to be buffered by molecules such as citrate, phosphate and histidines residues from proteins in the cytoplasm34,35, has fewer reported cellular roles. Hence, its presence at such high cellular concentrations suggests that in S. pneumoniae it could occur predominantly as a labile pool of cytoplasmic-buffered Mn2+. By contrast Zn2+, which is reported in numerous studies to be highly utilized in intracellular proteins36,37, has been predicted to have a significantly smaller labile pool10,36. Comprehensive intracellular metal ion speciation studies have been limited by a combination of technical and methodological challenges that remain to be surmounted. However, the data here allow us to speculate that, in S. pneumoniae, the potentially larger labile pool of Mn2+ ions, relative to Zn2+ ions, could be one mechanism that aids in Mn2+-dependent proteins correctly acquiring their cognate metal cofactor. The millimolar cellular quotient of Zn2+ ions in S. pneumoniae may also provide a plausible basis for the unexplained lower sensitivity of the zinc-responsive transcriptional regulator AdcR to Zn2+ ions, when compared with the homoplastic regulator, Zur, from E. coli10,15. In E. coli, Zn2+ has been shown to accumulate to an overall (cytoplasmic plus periplasmic) cellular concentration of ~0.2 mM and Zur is highly sensitive to Zn2+ abundance, sensing Zn2+ ions in the femtomolar range10. By contrast, AdcR is 2–3 orders of magnitude less sensitive, suggesting that a higher cellular ‘set-point’ for sensing of Zn2+ ions in S. pneumoniae may exist15.
In the context of exposure to Cd2+, the impact on transition metal ion homeostasis corresponds to specific dysregulation of Mn2+ and Zn2+ accumulation, but via different mechanisms. Manganese accumulation was disrupted by direct competition of Cd2+ for the Psa permease and the upregulation of the Mn2+-efflux pathway MntE. These results also suggest that PsaR, the transcriptional regulator of the Psa permease, is not functionally activated by Cd2+ and thus permits upregulation of psaBCA. In this way, the interaction of Cd2+ with PsaR may mimic the binding of Zn2+ rather than Mn2+, which occurs in a manner that does not permit repression of psaBCA transcription12. Although competition for the Psa permease was consistent with expectations, based on the inability of the SBP to be specific for solely Mn2+ ions, the mistranslocation of Cd2+ ions via the permease highlights an evolutionary limitation in managing extracellular stress by non-physiological elements. Specific acquisition of Mn2+ via the Psa permease arises from a combination of mechanisms that prevent the transport of Zn2+ ions, which are more abundant in the native niche of S. pneumoniae. However, the larger ionic radius and ‘strained’ coordination of the Cd2+ ion at the metal-binding site, similar to Mn2+ (Supplementary Table 3), resulted in an easily destabilized metal–protein complex, such that PsaA was incapable of preventing release of Cd2+ ions into the permease. This underscores the challenge encountered by metalloregulatory mechanisms when interacting with elements that have only recently entered the biosphere. In PsaA, the regulatory mechanisms that have evolved to prevent mistranslocation of Zn2+ ions are not capable of preventing the translocation of Cd2+ ions. By contrast with Mn2+, the Cd2+-induced disruption of Zn2+ homeostasis occurred via upregulation of czcD, the Zn2+-efflux pathway, and the apparent inability of the pneumococcus to transcriptionally activate additional Zn2+-recruiting proteins (that is, AdcAII (ref. 13)) to compensate for the depletion of cellular Zn2+ ions. Hence, Cd2+ accumulation activated the coordinated pneumococcal physiological response to Zn2+ intoxication or overload13,15,17,23, resulting in the futile export and depletion of cellular Zn2+. Our findings suggest that Cd2+ accumulation results in the mismetallation of the Zn2+-responsive transcriptional regulators, AdcR and SczA13,15,17. Whether this is due to Cd2+ directly interacting with the Zn2+-binding sites of intracellular metal sensors38,39,40 or via Cd2+-mediated displacement of Zn2+ from thiol buffering sites41, which results in the non-physiological activation of the Zn2+-responsive transcriptional regulators, remains to be determined. Irrespective of this fact, transcriptional activation of czcD is often construed as a mechanism to facilitate Cd2+ efflux based on studies of the archetypal protein from C. metallidurans18. However, in S. pneumoniae, similar to S. pyogenes42, CzcD is primarily a Zn2+-efflux pathway and does not provide a major protective role against Cd2+ ions. As a consequence, in S. pneumoniae, the Cd2+-induced upregulation of czcD appears to preferentially deplete intracellular Zn2+, despite the greater abundance of Cd2+ ions.
Cd2+ accumulation is associated with oxidative stress, despite the redox-inert nature of the ion4. The data here show that, in S. pneumoniae, the increased sensitivity to oxidative stress arises from the loss of sodA expression and activity due to Cd2+-induced Mn2+ depletion. However, the loss of SodA was an indirect by-product of Cd2+ accumulation and, importantly, not associated with the ability of the pneumococcus to tolerate Cd2+ ions. This was demonstrated by the ΔsodA strain, which showed wild-type growth phenotypes in the presence of extracellular Cd2+, and in wild-type S. pneumoniae, which retained the ability to accumulate high cellular concentrations of Cd2+ without an increase in sensitivity to oxidative stress, when supplemented with Mn2+. However, SodA still has a major role in the in vitro and in vivo fitness of S. pneumoniae owing to a lack of other mechanisms to detoxify superoxide23,30,33. Here, in the presence of Cd2+, the loss of SodA also occurs in conjunction with dysregulation of Mn2+ and Zn2+ homeostatic mechanisms and the accumulation of very high concentrations of Cd2+. Hence, it is the combination of these factors that compromises pneumococcal viability in the presence of Cd2+.
Intracellular accumulation of Cd2+ in S. pneumoniae was crucially dependent on glutathione, which acts as a low-affinity metal ion buffer for certain transition metal ions (Fe2+, Zn2+ and Cd2+), as phenotypic growth perturbations were seen when Cd2+ accumulation exceeded the total cellular glutathione pool (19–23 mM). Although it cannot be completely discounted that glutathione contributes to pneumococcal oxidative stress management32, the hypersensitivity of the ΔgshT strain to extracellular Cd2+, in contrast to the ΔsodA strain, strongly implicates the major role for cellular glutathione is in metal ion buffering. The observed hypersensitivity of the ΔgshT/ΔczcD strain to Zn2+ stress further underscores how intracellular metal ion homeostasis relies on the contributions of both cellular glutathione and the efflux machinery13,43. The results are also consistent with the central role of low-molecular-weight thiols for metal ion homeostasis in other microorganisms. Bacillithiol has recently been shown to have a crucial role in the buffering of Zn2+ ions in Bacillus subtilis41, while it also serves an undefined, but protective role in Cd2+ stress for B. subtilis and Staphylococcus aureus44. Although S. pneumoniae and other microorganisms lack a bacillithiol biosynthetic pathway, the role of low-molecular-weight species for metal ion homeostasis is emerging as a crucial mechanism. Overall, our findings here highlight the role of glutathione as an intracellular metal ion buffer, in preference to oxidative stress management. This finding has broad ramifications for the assumed role(s) of cellular glutathione, and related thiol derivatives, in prokaryotes and potentially eukaryotes43,41.
In conclusion, Cd2+ accumulation in the food chain and its toxicity are crucial, but largely overlooked concerns. Our findings reveal the molecular basis by which Cd2+ is mistranslocated via a Mn2+ transporter and provides new insights into how Cd2+ dysregulates intracellular transition metal homeostasis. Disruption of multiple essential metal ion homeostatic mechanisms is highly damaging to biological organisms, such as S. pneumoniae14, and in this way Cd2+ acts to severely dysregulate both Mn2+ and Zn2+ homeostasis (Supplementary Fig. 8). By revealing the remarkably high (millimolar) cellular quotients of transition metal ions and glutathione, this work also shows how Cd2+, despite its inability to directly generate reactive oxygen species, is associated with oxidative stress. This work provides a new understanding of the mechanisms by which Cd2+ enters cells and causes toxicity, and thereby, opens the way to identifying new routes towards developing specific therapeutic agents capable of preventing Cd2+ toxicity.
Methods
Growth experiments and whole-cell assays
The S. pneumoniae D39 ΔsodA, ΔczcD, ΔgshT, ΔmntE, ΔadcCBA and ΔadcA/ΔadcAII strains have been generated previously13,32. The ΔmntE and ΔgshT/ΔczcD strains were generated as described previously13, using primer sequences listed in Supplementary Table 6. S. pneumoniae D39, ΔsodA, ΔczcD, ΔmntE, ΔgshT, ΔgshT/ΔczcD, ΔadcCBA and ΔadcA/ΔadcAII were grown in CDM, which corresponded to the C+Y media without transition metal ion supplementation23. The base ion content of the CDM was ascertained by ICP–MS on an Agilent 7500cx ICP–MS (Adelaide Microscopy, University of Adelaide)23. Growth experiments were conducted in CDM supplemented with 1 μM MnSO4 and concentrations of 10, 30, 50 or 80 μM of CdCl2 or MnSO4, as specified. All chemicals used in this study were purchased from Sigma-Aldrich, unless otherwise specified. For bacterial growth experiments, an inoculum was prepared from overnight grown culture on a blood-agar (BA) plate and resuspended in CDM to an absorbance at 600 nm (A600) of 1.0. The inoculum was then diluted in 200 μl of CDM in a 96-well flat-bottom plate (Greiner Bio One) to a final A600 of 0.05 and sealed with gas-permeable seal (Breathe-Easy, Diversified Biotech). The plate was then incubated in a FLUOStar Omega spectrophotometer with an ACU gas controller (BMG Labtech) at 310 K in 5% CO2. A600 readings were recorded using the well-scan function every 30 min. Data from at least six independent growth experiments was averaged to ascertain the effect of CdCl2 on bacterial growth. Bacterial density at A600 was measured as the c.f.u. per ml determined by serial dilutions on BA plates.
Bacterial growth for ICP–MS, paraquat killing and glutathione assays used identical growth parameters to the microplate experiments, with MnSO4 and CdCl2 supplementation as specified. For ICP–MS, 50 ml of culture was grown to A600=0.3–0.4, harvested and prepared for analysis by ICP–MS, as described previously23. For the paraquat killing assays, 1 ml of A600=0.3 culture was incubated for 30 min with 10 mM paraquat and then serially diluted and plated on blood agar. Total glutathione assays were performed using 1 ml of A600=0.3 culture with the Promega GSH-Glo assay kit according to the manufacturer’s instructions (Promega, USA). To examine the effect of Cd2+ stress on cellular SodA activity, S. pneumoniae D39 was grown to an A600 of 0.3 as for the ICP–MS experiments in CDM supplemented with 1 μM MnSO4, 30 μM CdCl2:1 μM MnSO4 or 30 μM CdCl2:30 μM MnSO4. The cells were washed three times in PBS before lysis by sonication and intact cells and other insoluble material was removed by centrifugation at 277 K for 30 min at 14,000 g. The assay was performed using a SOD activity kit (Calbiochem) and the data were normalized to the total protein concentration of the cell-free extract, as determined using the Dc Bio-Rad protein determination assay, followed by normalization (in percentage) to the average SOD activity measured for the untreated cells (1 μM MnSO4).
Metal sensitivity assays were also performed using a S. pneumoniae drop test. Bacteria were grown in CDM supplemented with 1 μM MnSO4 and grown until the A600 reached 0.3–0.4. Cells were then serially diluted 10-fold up to a 10−5 dilution, and 5 μl of each dilution were spotted on BA plates supplemented with varying concentrations of ZnSO4 (0, 100, 300 and 1,000 μM) or CdCl2 (0, 10, 30 and 100 μM). Plates were photographically documented following overnight incubation at 37 °C, 5% CO2.
RNA extraction and RT–PCR analysis
Pneumococci were grown as for the ICP–MS analyses, then 500 μl of the culture was mixed with 1 ml of RNA Protect (Qiagen). RNA was extracted and purified using an RNeasy Protect Bacteria Mini Kit (Qiagen) after enzymatic lysis using lysozyme and mutanolysin, all according to the manufacturer’s instructions. DNase I treatment (Roche) was performed followed by quantitative reverse transcription PCR using SuperScript III (Invitrogen) with a Roche LC480 Real-Time Cycler. The transcription levels of genes analysed were normalized to those obtained for 16S rRNA. Primer sequences are in Supplementary Table 6.
Cell volume and concentration determination
Scanning electron microscopy was used to determine the dimensions of the pneumococcal cell. Bacteria were grown, as described above, harvested and then prepared for, and analysed by a Philips XL30 FEG scanning electron microscope as described in ref. 45. Cell measurements were obtained using instrument software. The cell dimensions were used to calculate volume assuming an ellipsoid:

where r1, r2, and r3 are the radii of the ellipsoidal cell in three dimensions determined by scanning electron microscopy (SEM). The total quotient of transition metal ions was then derived using cell volume (V, litres), where the total cell density is known (c.f.u.), and concentration of metal ions in a sample of known volume and known number of cells (M, moles).

The derived concentration represents mean molarity of metal ion per cell of mean dimensions.
Expression and purification of apo-PsaA
Recombinant PsaA was expressed in E. coli LEMO21(DE3) from the pCAMcLIC01-PsaA construct8. The dodecahistidine tag was removed from affinity-purified PsaA by enzymatic digestion by the human rhinovirus 3C protease and the protein purified further on a HisTrap HP column. Apo-PsaA was prepared by dialyzing the dodecahistidine tag-cleaved protein in a 20-kDa molecular-weight-cutoff membrane (Slide-A-Lyzer, Pierce) against 4 l of sodium acetate buffer, pH 4.0, with 20 mM EDTA at 300 K. The sample was then dialysed against 4 l of 20 mM Tris-HCl, pH 7.2, and 100 mM NaCl, at 277 K and centrifuged at 18,000 g for 10 min to remove any insoluble material. Protein samples were analysed for metal content by heating 5 μM protein at 370 K for 15 min in 3.5% HNO3 and the metal ion content was measured by ICP–MS.
Protein assays
Protein concentration determination was performed using the Dc Bio-Rad protein determination assay. Metal-loading assays were performed on purified apo-PsaA (30 μM) by mixing with 300 μM CdCl2 in a total volume of 2 ml in the assay buffer (20 mM MOPS, pH 7.2, and 100 mM NaCl) for 60 min at 277 K. The sample was desalted on a PD10 column (GE Healthcare) into the assay buffer and the protein concentration was quantified. Protein was then either kept for ICP–MS analysis or mixed with 3 mM EDTA in a total volume of 2 ml for 60 min at 277 K. Samples were then desalted on a PD10 column as before. Solutions (5–10 μM) of control, metal-loaded and EDTA-treated protein were prepared in 3.5% HNO3 and boiled for 15 min at 370 K. Samples were then cooled and centrifuged for 20 min at 14,000 g. The supernatant was then analysed by ICP–MS and the protein-to-metal ratio was determined.
Immunoblot analyses of PsaA expression levels
Wild-type and mutant S. pneumoniae were grown under the same conditions as for ICP–MS. After reaching an A600 of 0.4, cells were incubated with 0.1% sodium deoxycholate at 310 K for 60 min to induce lysis. Protein concentrations were determined and 10 μg of total protein was loaded into each lane. After electrophoretic separation by SDS–PAGE, proteins were transferred to a nitrocellulose membrane using the iBlot (Life Technologies) system. The blots were incubated with murine anti-PsaA serum (1:2,000; ref. 23), followed by anti-mouse IRDye 800 (LI-COR; 1:50,000), and were scanned using an Odyssey infrared imaging system (LI-COR). Band intensities were measured using the manufacturer’s application software and the results correspond to the mean (±s.e.m.) of two independent biological experiments.
Determination of KD for PsaA with Cd2+
Excitation–emission spectra were determined on a FLUOStar Omega (BMG Labtech) at 301 K using black half-volume 384-well microtitre plates (Greiner Bio One). All experiments were performed in 20 mM MOPS (pH 6.7) and 50 mM NaCl with FluoZin-3 (Life Technologies) at a concentration of 50 nM. Deionized water and buffers were prepared and treated with Chelex-100 (Sigma-Aldrich) to avoid metal contamination. Filters used for FluoZin-3 were excitation (485±10 nm) and emission (520±5 nm). To determine the dissociation constant between a metal (X) and FluoZin-3 (F), we considered the following equilibria:

where, for a metal that increases the fluorescence of the probe by more than ~10%, the following equation, which is an exact analytical relationship derived from the mass action equation for the formation of a 1:1 complex between probe and metal, was used to estimate the dissociation constant, KD.X

where f is the measured fluorescence intensity in the presence of metal, fmax is the fluorescence in the presence of saturating metal and fmin is the fluorescence in the absence of metal. In all cases, a low concentration (50 nM) of probe was used and we assumed that the free-metal concentration was equal to the added metal concentration. The mean±s.e.m. (n=6) KD determined for FluoZin-3 with CdCl2 in the buffer system used in this study was determined to be 250±35 nM. Competition by PsaA for Cd2+ binding was assessed by monitoring the decrease in the fluorescence of 50 nM FluoZin-3-Cd2+ in response to increasing apo-PsaA concentrations and analysed using log10[inhibitor] versus response model, with the experimentally derived KD for FluoZin-3 with Cd2+, in GraphPad prism to determine the KD value for Cd2+ binding by PsaA.
Protein crystallization and structure determination
Cd-bound PsaA-D280N protein crystals were obtained in 12.5% (w/v) polyethylene glycol (PEG) 1000, 12.5% (w/v) PEG 3350, 12.5% (v/v) MPD, 0.1 M Trizma-Bicine, pH 8.7, and 0.01 M CdCl2 using the hanging drop vapour diffusion method8. Cd2+-bound wild-type PsaA crystals were grown in 26–36% PEG 400, 0.1 M NaCl, 0.1 M Trizma-HCl, pH 8.0, and 0.1 M CdCl2 also using vapour diffusion. Before data collection, the crystals were flash-cooled by rapid immersion in liquid nitrogen. The diffraction data were collected on a single crystal at the Australian Synchrotron (MX2 beamline). Cd2+-bound D280N and wild-type PsaA structures were determined by molecular replacement with Phaser46 using the Mn2+-bound D280N (PDB accession code 3ZKA) and the Zn2+-bound (PDB accession code 1PSZ) PsaA crystal structures, respectively, as phasing models followed by automatic model building in Phenix.AutoBuild47. The structures were refined iteratively using Phenix.Refine and Refmac5 manual model fitting in Coot48. Both structures had two molecules (chains A and B) in each asymmetric unit and chain A molecules were used in subsequent structural analyses. Structural analyses (superpositions, metal ion coordination and N-/C-terminal domain-crossing angles) were performed in Chimera49. Secondary structure elements were assigned by DSSP (version 2.1.0; ref. 50). The WHAT-IF51 and PISA52 web services were used to identify hydrogen bonds and salt bridges. Data processing and structure refinement statistics can be found in Supplementary Table 7.
Additional information
Accession codes: Coordinates and structure factors for Cd2+-bound PsaA-D280N, partially occluded state, and for Cd2+-bound wild-type PsaA, closed state, have been deposited in the RCSB Protein Data Bank under accession codes 4uto and 4utp, respectively.
How to cite this article: Begg, S. L. et al. Dysregulation of transition metal ion homeostasis is the molecular basis for cadmium toxicity in Streptococcus pneumoniae. Nat. Commun. 6:6418 doi: 10.1038/ncomms7418 (2015).
References
Nriagu, J. O. & Pacyna, J. M. Quantitative assessment of worldwide contamination of air, water and soils by trace metals. Nature 333, 134–139 (1988).
Joseph, P. Mechanisms of cadmium carcinogenesis. Toxicol. Appl. Pharmacol. 238, 272–279 (2009).
Lane, T. W. et al. A cadmium enzyme from a marine diatom. Nature 435, 42 (2005).
Cuypers, A. et al. Cadmium stress: an oxidative challenge. Biometals 23, 927–940 (2010).
Archibald, F. S. & Duong, M. N. Manganese acquisition by Lactobacillus plantarum. J. Bacteriol. 158, 1–8 (1984).
Hubert, N. & Hentze, M. W. Previously uncharacterized isoforms of divalent metal transporter (DMT)-1: implications for regulation and cellular function. Proc. Natl Acad. Sci. USA 99, 12345–12350 (2002).
McAllister, L. J. et al. Molecular analysis of the psa permease complex of Streptococcus pneumoniae. Mol. Microbiol. 53, 889–901 (2004).
Couñago, R. M. et al. Imperfect coordination chemistry facilitates metal ion release in the Psa permease. Nat. Chem. Biol. 10, 35–41 (2014).
Dintilhac, A. & Claverys, J. P. The adc locus, which affects competence for genetic transformation in Streptococcus pneumoniae, encodes an ABC transporter with a putative lipoprotein homologous to a family of streptococcal adhesins. Res. Microbiol. 148, 119–131 (1997).
Outten, C. E. & O'Halloran, T. V. Femtomolar sensitivity of metalloregulatory proteins controlling zinc homeostasis. Science 292, 2488–2492 (2001).
Irving, H. & Williams, R. J. P. The stability of transition-metal complexes. J. Chem. Soc. 3192–3210 (1953).
Lisher, J. P., Higgins, K. A., Maroney, M. J. & Giedroc, D. P. Physical characterization of the manganese-sensing pneumococcal surface antigen repressor from Streptococcus pneumoniae. Biochemistry 52, 7689–7701 (2013).
Plumptre, C. D. et al. AdcA and AdcAII employ distinct zinc acquisition mechanisms and contribute additively to zinc homeostasis in Streptococcus pneumoniae. Mol. Microbiol. 91, 834–851 (2014).
Dintilhac, A., Alloing, G., Granadel, C. & Claverys, J. P. Competence and virulence of Streptococcus pneumoniae: Adc and PsaA mutants exhibit a requirement for Zn and Mn resulting from inactivation of putative ABC metal permeases. Mol. Microbiol. 25, 727–739 (1997).
Reyes-Caballero, H. et al. The metalloregulatory zinc site in Streptococcus pneumoniae AdcR, a zinc-activated MarR family repressor. J. Mol. Biol. 403, 197–216 (2010).
Loisel, E. et al. AdcAII, a new pneumococcal Zn-binding protein homologous with ABC transporters: biochemical and structural analysis. J. Mol. Biol. 381, 594–606 (2008).
Kloosterman, T. G., van der Kooi-Pol, M. M., Bijlsma, J. J. & Kuipers, O. P. The novel transcriptional regulator SczA mediates protection against Zn2+ stress by activation of the Zn2+-resistance gene czcD in Streptococcus pneumoniae. Mol. Microbiol. 65, 1049–1063 (2007).
Anton, A., Grosse, C., Reissmann, J., Pribyl, T. & Nies, D. H. CzcD is a heavy metal ion transporter involved in regulation of heavy metal resistance in Ralstonia sp. strain CH34. J. Bacteriol. 181, 6876–6881 (1999).
Rosch, J. W., Gao, G., Ridout, G., Wang, Y. D. & Tuomanen, E. I. Role of the manganese efflux system mntE for signalling and pathogenesis in Streptococcus pneumoniae. Mol. Microbiol. 72, 12–25 (2009).
Crupper, S. S., Worrell, V., Stewart, G. C. & Iandolo, J. J. Cloning and expression of cadD, a new cadmium resistance gene of Staphylococcus aureus. J. Bacteriol. 181, 4071–4075 (1999).
Fu, Y. et al. A new structural paradigm in copper resistance in Streptococcus pneumoniae. Nat. Chem. Biol. 9, 177–183 (2013).
Brenot, A., Weston, B. F. & Caparon, M. G. A PerR-regulated metal transporter (PmtA) is an interface between oxidative stress and metal homeostasis in Streptococcus pyogenes. Mol. Microbiol. 63, 1185–1196 (2007).
McDevitt, C. A. et al. A molecular mechanism for bacterial susceptibility to zinc. PLoS Pathog. 7, e1002357 (2011).
Lawrence, M. C. et al. The crystal structure of pneumococcal surface antigen PsaA reveals a metal-binding site and a novel structure for a putative ABC-type binding protein. Structure 6, 1553–1561 (1998).
Braun, W. et al. Comparison of the NMR solution structure and the x-ray crystal structure of rat metallothionein-2. Proc. Natl Acad. Sci. USA 89, 10124–10128 (1992).
Graham, S. C., Maher, M. J., Simmons, W. H., Freeman, H. C. & Guss, J. M. Structure of Escherichia coli aminopeptidase P in complex with the inhibitor apstatin. Acta Crystallogr. D Biol. Crystallogr. 60, 1770–1779 (2004).
Price, N. M. & Morel, F. M. M. Cadmium and cobalt substitution for zinc in a marine diatom. Nature 344, 658–660 (1990).
Xu, Y., Feng, L., Jeffrey, P. D., Shi, Y. & Morel, F. M. Structure and metal exchange in the cadmium carbonic anhydrase of marine diatoms. Nature 452, 56–61 (2008).
Ogunniyi, A. D. et al. Central role of manganese in regulation of stress responses, physiology, and metabolism in Streptococcus pneumoniae. J. Bacteriol. 192, 4489–4497 (2010).
Eijkelkamp, B. A. et al. Extracellular zinc competitively inhibits manganese uptake and compromises oxidative stress management in Streptococcus pneumoniae. PLoS ONE 9, e89427 (2014).
Yesilkaya, H. et al. Role of manganese-containing superoxide dismutase in oxidative stress and virulence of Streptococcus pneumoniae. Infect. Immun. 68, 2819–2826 (2000).
Potter, A. J., Trappetti, C. & Paton, J. C. Streptococcus pneumoniae uses glutathione to defend against oxidative stress and metal ion toxicity. J. Bacteriol. 194, 6248–6254 (2012).
Tseng, H. J., McEwan, A. G., Paton, J. C. & Jennings, M. P. Virulence of Streptococcus pneumoniae: PsaA mutants are hypersensitive to oxidative stress. Infect. Immun. 70, 1635–1639 (2002).
Hider, R. C. & Kong, X. L. Glutathione: a key component of the cytoplasmic labile iron pool. Biometals 24, 1179–1187 (2011).
Tabares, L. C. & Un, S. In situ determination of manganese(II) speciation in Deinococcus radiodurans by high magnetic field EPR: detection of high levels of Mn(II) bound to proteins. J. Biol. Chem. 288, 5050–5055 (2013).
Sun, X. et al. Putative copper- and zinc-binding motifs in Streptococcus pneumoniae identified by immobilized metal affinity chromatography and mass spectrometry. Proteomics 11, 3288–3298 (2011).
Andreini, C., Bertini, I., Cavallaro, G., Holliday, G. L. & Thornton, J. M. Metal ions in biological catalysis: from enzyme databases to general principles. J. Biol. Inorg. Chem. 13, 1205–1218 (2008).
Ralston, D. M. & O'Halloran, T. V. Ultrasensitivity and heavy-metal selectivity of the allosterically modulated MerR transcription complex. Proc. Natl Acad. Sci. USA 87, 3846–3850 (1990).
Guerra, A. J. & Giedroc, D. P. Metal site occupancy and allosteric switching in bacterial metal sensor proteins. Arch. Biochem. Biophys. 519, 210–222 (2012).
Sun, Y., Wong, M. D. & Rosen, B. P. Role of cysteinyl residues in sensing Pb(II), Cd(II), and Zn(II) by the plasmid pI258 CadC repressor. J. Biol. Chem. 276, 14955–14960 (2001).
Ma, Z. et al. Bacillithiol is a major buffer of the labile zinc pool in Bacillus subtilis. Mol. Microbiol. 94, 756–770 (2014).
Ong, C. L., Gillen, C. M., Barnett, T. C., Walker, M. J. & McEwan, A. G. An antimicrobial role for zinc in innate immune defense against group A Streptococcus. J. Infect. Dis. 209, 1500–1508 (2014).
Helbig, K., Bleuel, C., Krauss, G. J. & Nies, D. H. Glutathione and transition-metal homeostasis in Escherichia coli. J. Bacteriol. 190, 5431–5438 (2008).
Rajkarnikar, A. et al. Analysis of mutants disrupted in bacillithiol metabolism in Staphylococcus aureus. Biochem. Biophys. Res. Commun. 436, 128–133 (2013).
Fairbrother, L. et al. Effect of the cyanide-producing bacterium Chromobacterium violaceum on ultraflat Au surfaces. Chem. Geol. 265, 313–320 (2009).
McCoy, A. J. Solving structures of protein complexes by molecular replacement with Phaser. Acta Crystallogr. D Biol. Crystallogr. 63, 32–41 (2007).
Terwilliger, T. C. et al. Iterative model building, structure refinement and density modification with the PHENIX AutoBuild wizard. Acta Crystallogr. D Biol. Crystallogr. 64, 61–69 (2008).
Emsley, P., Lohkamp, B., Scott, W. G. & Cowtan, K. Features and development of Coot. Acta Crystallogr. D Biol. Crystallogr. 66, 486–501 (2010).
Pettersen, E. F. et al. UCSF Chimera--a visualization system for exploratory research and analysis. J. Comput. Chem. 25, 1605–1612 (2004).
Kabsch, W. & Sander, C. Dictionary of protein secondary structure: pattern recognition of hydrogen-bonded and geometrical features. Biopolymers 22, 2577–2637 (1983).
Hekkelman, M. L. et al. WIWS: a protein structure bioinformatics Web service collection. Nucleic Acids Res. 38, W719–W723 (2010).
Krissinel, E. & Henrick, K. Inference of macromolecular assemblies from crystalline state. J. Mol. Biol. 372, 774–797 (2007).
Acknowledgements
We acknowledge the use of the Australian Synchrotron (Victoria, Australia) and the University of Queensland Remote-Operation X-Ray (UQ-ROCX) Diffraction Facility. This work was supported by the Australian Research Council (ARC) grant DP120103957 to C.A.M. and J.C.P., and the National Health and Medical Research Council (NHMRC) Project grants 1022240, to C.A.M., and 1080784, to C.A.M. and M.J.M., and Program grants 565526, to A.G.M., B.K. and J.C.P., and 1071659, to B.K. and J.C.P. M.L.O. holds an ARC DECRA (DE1211550), B.K. is a NHMRC Senior Research Fellow (1003325) and J.C.P. is a NHMRC Senior Principal Research Fellow (1043070). We thank the beamline staff at the Australian Synchrotron for their support and Dr C. Adolphe for discussions.
Author information
Authors and Affiliations
Contributions
S.L.B., B.A.E. and C.A.M. designed and executed all the biochemical studies. C.-l.Y.O. generated the mutant strains. Z.L., R.M.C. and M.J.M. designed and executed the crystallographic experiments. S.L.B., R.M.C., M.J.M., M.L.O. and C.A.M. drafted the manuscript. All authors contributed to the design, analysis, discussion of the research and writing of the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Supplementary Information
Supplementary Figures 1-8 and Supplementary Tables 1-7 (PDF 1387 kb)
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Begg, S., Eijkelkamp, B., Luo, Z. et al. Dysregulation of transition metal ion homeostasis is the molecular basis for cadmium toxicity in Streptococcus pneumoniae. Nat Commun 6, 6418 (2015). https://doi.org/10.1038/ncomms7418
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/ncomms7418
This article is cited by
-
A Matter of Metals: Copper but Not Cadmium Affects the Microbial Alpha-Diversity of Soils and Sediments — a Meta-analysis
Microbial Ecology (2023)
-
Cadmium stress dictates central carbon flux and alters membrane composition in Streptococcus pneumoniae
Communications Biology (2020)
-
Photosynthetic semiconductor biohybrids for solar-driven biocatalysis
Nature Catalysis (2020)
-
Using Chemical Modifiers and Increasing the Pyrolysis Temperature for High-Sensitivity Spectrometric Determination of Cadmium in Dairy Products
Journal of Applied Spectroscopy (2020)
-
Zinc-binding to the cytoplasmic PAS domain regulates the essential WalK histidine kinase of Staphylococcus aureus
Nature Communications (2019)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
