
Abstract
Upon activation, naive CD4 T cells differentiate into a variety of T-helper-cell subsets characterized by different cytokine production and functions. Currently, lineage commitment is considered to depend mostly on the environmental context to which naive CD4 T cells are exposed. Here we challenge this model based on the supposed homogeneity of the naive CD4 T-cell compartment. We show that peripheral naive CD4 T cells can be subdivided into two subsets according to Ly-6C expression. Furthermore, the two newly defined subsets (Ly-6C− and Ly-6C+ naive CD4 T cells) are not equal in their intrinsic ability to commit into the induced regulatory T-cell lineage. Finally, phenotypic analysis, imaging and adoptive transfer experiments reveal that Ly-6C expression is modulated by self-recognition, allowing the dichotomization of the naive CD4 T-cell compartment into two cell subsets with distinct self-reactivity. Altogether, our results show that naive CD4 T cells with the highest avidity for self are prone to differentiate into regulatory T cells.
Similar content being viewed by others


Introduction
In normal young adult mice, the peripheral CD4 T-cell compartment is composed of almost 80% naive T cells. The remaining 20% are distributed between regulatory and effector/memory-phenotype CD4 T cells. Following activation by antigen-presenting cells (APCs) in the periphery, naive CD4 T (CD4 TN) cells can differentiate into a variety of well-documented T-helper (TH) cell subsets, such as TH1, TH2, TH17 or induced regulatory T (iTreg) cells, characterized by their cytokine-production profiles and specific effector functions. For instance, TH1 cells are characterized by their production of interferon (IFN)-γ, a potent activator of cell-mediated immunity1,2. TH2 cells are characterized by production of interleukin (IL)-4, IL-5 and IL-13, which are potent activators of B-cell immunoglobulin production1,2. In addition, TH17 cells are characterized by IL-17A, IL-17F and IL-22 secretion and are described to have an important role in anti-microbial responses3,4. Finally, iTreg cells produce transforming growth factor-β (TGF-β) and share phenotypic and functional characteristics with natural regulatory T cells that have a crucial role in maintaining peripheral self-tolerance5. Identification of T-bet, GATA-3, RORγt/RORα and Foxp3 as lineage-defining transcription factors solidified the categorization into TH1, TH2, TH17 and iTreg effector cell subsets6,7,8,9,10. Additional TH cell subsets have also been described, such as IL-10 producing type 1 regulatory T cells11, IL-9 producing TH9 cells12 or TFH cells13.
Lineage commitment leads to a broad diversity of effector cell functions and is considered to depend mostly on the immunological context in which CD4 TN cells are immersed at the time of their activation. Indeed, the differentiation fate appears predominantly governed by extrinsic factors such as cytokines or environmental cues. Specifically, in vitro culture assays and in vivo models have been used to establish that IL-12, IL-4, TGF-β alone or TGF-β in combination with pro-inflammatory cytokines, such as IL-6, are crucially required for TH1, TH2, iTreg or TH17-cell differentiation, respectively14,15,16,17,18. This commonly accepted model implicitly posits that the CD4 TN cells on which the various cytokines act constitute an homogeneous population of cells. However, intrinsic heterogeneity within the CD4 TN-cell pool such as TCR affinity for antigen or sensitivity to co-stimulatory signals and cytokines should be taken into account when considering, in vivo, CD4 TN-cell differentiation19.
In light of these considerations, we decided to study whether intrinsic properties of CD4 TN cells, in association with the cytokine environment in which they are immersed, could contribute to helper CD4 T-cell commitment. Here we show that peripheral CD4 TN cells can be subdivided into two subsets according to Ly-6C expression. These two newly defined subsets are not equal in their intrinsic ability to commit into the iTreg-cell lineage with Ly-6C− CD4 TN cells differentiating more efficiently into iTreg cells than their Ly-6C+-cell counterparts both in vivo and in vitro.
Results
Heterogeneity of Ly-6C expression in CD4 TN cells
Throughout this study, CD4 TN lymphocytes were defined as Foxp3− CD25− CD44−/lo CD4+ CD8α− TCRβ+ cells (Supplementary Fig. S1a). Although the CD4 TN-cell compartment is commonly considered as homogenous, we observed, as previously described20,21, that CD4 TN cells from the secondary lymphoid organs of C57BL/6 mice (peripheral lymph node (pLN) and mesenteric lymph node (mLN) and spleen) can be subdivided into two subsets according to Ly-6C expression (Fig. 1a,b). This applied also to CD4 TN cells from bone marrow (BM), Peyer’s patches, blood and peritoneal cavity. Although the proportion of CD4 TN cells varied greatly between these organs, in all of them, about two-thirds of CD4 TN cells expressed Ly-6C (Supplementary Fig. S1b). In contrast, Ly-6C+ CD4 TN cells were almost completely absent in the thymus.

(a) Ly-6C fluorescence histograms of CD4 TN cells (CD4+ CD8α− TCRβ+ Foxp3− CD25− CD44−/lo T cells) recovered from pLNs, mLNs, spleen (Spl) and thymus (Thy) are shown for a representative C57BL/6 Foxp3-GFP mouse. Numbers in each histogram represent the percentage of Ly-6C+ cells among CD4 TN cells. (b) The proportions of Ly-6C− and Ly-6C+ cells among CD4 TN cells recovered from the thymus, BM, peritoneal lavage fluid (PLF), blood, Peyer’s patches (PP), spleen, mLNs and pLNs of C57BL/6 mice were calculated. Results are shown as means±s.e.m. for six mice from two independent experiments. (c) CD69, CD45RB, CD62L and CD127 fluorescence histograms of pLN Ly-6C− CD4 TN cells, Ly-6C+ CD4 TN cells, regulatory CD4 T cells (CD4 Treg defined as Foxp3+ CD4+ CD8α− TCRβ+) and memory CD4 T cells (CD4 Tmem defined as Foxp3− CD44hi CD4+ CD8α− TCRβ+ cells) are shown for a representative C57BL/6 Foxp3-GFP mouse. The histograms are representative of at least six mice from three individual experiments. (d) The proportions of IL-2, IL-4, IL-10, IL-17, IFN-γ or TNF-α producing cells among memory, regulatory, Ly-6C− and naive Ly-6C+ CD4 TN cells recovered from pLNs of C57BL/6 Foxp3-GFP mice were estimated after a pulse stimulation with phorbol myristate acetate and ionomycin. Results are shown as means±s.e.m. for six mice from two independent experiments.
Both Ly-6C− and Ly-6C+ CD4 TN cells from pLNs exhibited a true naive phenotype illustrated by their high expression of CD45RB, CD62L and CD127 and their lack of expression of CD69 (Fig. 1c). Consistent with their naive phenotype, both TN-cell subsets were not able to produce IL-2, IL-4, IL-10, IL-17 or IFN-γ in response to stimulation (Fig. 1d). By contrast, memory-phenotype CD4 T cells (CD4 Tmem cells) produced all of these cytokines and, as expected, regulatory CD4 T cells (CD4 Treg cells) synthesized IL-10. Finally, we observed that a similar proportion of Ly-6C− and Ly-6C+ CD4 TN cells produced TNF-α. Therefore, Ly-6C expression divides the peripheral CD4 TN-cell compartment into two subsets.
Pathogenicity of Ly-6C− and Ly-6C+ CD4 TN cells
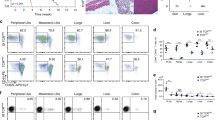
To test whether the phenotypic heterogeneity of the CD4 TN-cell compartment might reveal differential expansion/differentiation potentials, highly purified Ly-6C− or Ly-6C+ CD4 TN cells were transferred into T-cell-deficient recipients (Fig. 2a, Supplementary Fig. S2). T-cell compromised animals injected with CD4 TN cells are known to rapidly develop inflammatory bowel disease (IBD)22,23. Interestingly, mice injected with Ly-6C− CD4 TN cells developed with time a more severe wasting disease than those injected with Ly-6C+ CD4 TN cells (Fig. 2b). This correlated with higher histological colitis scores (determined 7 weeks after transfer by examination of colon sections) when mice were injected with Ly-6C− CD4 TN cells (Fig. 2c). Surprisingly, contrasting with clinical observations, Ly-6C− CD4 TN cells expanded to a lesser extent than their Ly-6C+-cell counterparts when transferred into T-cell-deficient recipients (Fig. 2d). Differences were not only quantitative as, in all studied secondary lymphoid organs, Ly-6C− CD4 TN cells gave rise to greater proportions and absolute numbers of iTreg cells as compared with Ly-6C+ CD4 TN cells (Fig. 2e, Supplementary Fig. S3). In addition, although Ly-6C+ CD4 TN-cell transfer led to higher proportions and absolute numbers of IFNγ-producing CD4 T cells in secondary lymphoid organs, the differentiation of Ly-6C− CD4 TN cells resulted in greater proportions and absolute numbers of newly generated IL-17+ CD4 T cells (Fig. 2f, Supplementary Fig. S3). Interestingly, the proportion of IL-17+ CD4 T cells was tightly correlated with the proportion of newly generated iTreg cells (Fig. 2g). These latter results suggest that, in the IBD experimental model, iTreg-cell generation promotes TH17 CD4 T-cell development as recently suggested in other settings24,25.

0.5 × 106 flow-cytometry-sorted Ly-6C− or Ly-6C+ CD4 TN cells from C57BL/6 Foxp3-GFP mice were injected intravenously into C57BL/6 CD3ε−/− mice. Seven weeks after transfer, pLNs, mLNs and spleen were recovered separately. (a) Diagram illustrating the experimental model. (b) Body weight of recipient mice was monitored up to 7 weeks post transfer, and percentages of initial body weight were calculated and plotted. (c) Colitis scores of recipient mice were assessed 7 weeks after transfer. (d) Absolute numbers of CD4 T cells recovered from pLNs, mLNs and spleen of recipient mice. (e) Foxp3-GFP/CD4 dot-plots for gated CD4+ CD8α− CD3+ T cells recovered from pLNs of representative recipient mice and proportion of Foxp3+ cells among CD4+ CD8α− CD3+ T cells recovered from pLNs, mLNs and spleen of recipient mice. (f) IL-17/IFNγ dot-plots for gated CD4+ CD8α− CD3+ T cells recovered from pLNs of representative recipient mice and proportion of IL-17- and IFNγ-producing CD4+ CD8α− CD3+ T cells recovered from pLNs, mLNs and spleen of recipient mice. (g) Correlation between the proportion of Foxp3-expressing and IL-17-producing cells among CD4+ CD8α− CD3+ T cells recovered from pLNs, mLNs and spleen of recipient mice. (b–c) Results are shown as means±s.e.m. for at least six mice per group per experiment, from four independent experiments. (d–g) Each dot represents an individual mouse. (b–f) Significance of differences between two series of results was assessed using a two-tailed unpaired Student’s t-test. (g) Correlation analyses were performed using Pearson’s correlation test. Values of P<0.05 were considered as statistically significant (*P<0.05; **P<0.01; and ***P<0.001).
iTreg cells promote TH17 differentiation in an IBD model
We then performed co-transfer experiments in which Ly-6C− CD4 TN cells from CD45.2 mice were co-injected with Ly-6C+ CD4 TN cells from CD45.1 mice into T-cell-deficient recipients (Fig. 3a). T-cell-deficient mice injected with Ly-6C− or Ly-6C+ CD4 TN cells alone were used in parallel as control mice (Supplementary Fig. S4a).

(a–c) Ly-6C− CD4 TN cells (0.25 × 106) from C57BL/6 Foxp3-GFP CD45.2 mice were injected intravenously (i.v.) into C57BL/6 CD3ε−/− mice together with 0.25 × 106 Ly-6C+ CD4 TN cells from C57BL/6 Foxp3-GFP CD45.1 mice. Seven weeks after transfer, pLNs, mLNs and spleen were recovered separately and analysed. (a) Diagram illustrating the experimental model. (b) Proportions of CD45.1+ or CD45.2+ CD4 T cells expressing Foxp3 or producing IL-17 or IFNγ recovered from pLNs, mLNs and spleen of recipient mice. (c) Absolute numbers of CD45.1+ or CD45.2+ CD4 T cells recovered from pLNs, mLNs and spleen of recipient mice. (d–f) Ly-6C− CD4 TN cells (0.5 × 106) from DEREG mice were injected i.v. into C57BL/6 CD3ε−/− mice treated or not twice a week with DT. (d) Diagram illustrating the experimental model. (e) Body weight of recipient mice was monitored up to 8 weeks post transfer, and percentages of initial body weight were calculated and plotted. (f) Proportions of IL-17-producing CD4+ CD8α− CD3+ T cells recovered from pLNs, mLNs and spleen of recipient mice. (b,c,f) Each dot represents an individual mouse. (e,f) Results are shown as means±s.e.m. for two independent experiments. Significance of differences between two series of results was assessed using a two-tailed paired (b,c) or unpaired (e,f) Student’s t-test. Values of P<0.05 were considered as statistically significant (*P<0.05; **P<0.01; and ***P<0.001). ns, not significant.
Mice co-injected with both Ly-6C− and Ly-6C+ CD4 TN cells developed a wasting disease and colitis as severe as mice injected with Ly-6C− CD4 TN cells alone (Supplementary Fig. S4b,c). When both cell subsets were injected together, Ly-6C− CD4 TN cells still gave rise to a higher proportion of newly differentiated iTreg cells than their Ly-6C+-cell counterparts (Fig. 3b). By contrast, in all secondary lymphoid organs studied, neither expansion nor TH17/TH1 differentiation appeared significantly different between both transferred cell subsets (Fig. 3b,c). More precisely, when injected together, Ly-6C− and Ly-6C+ CD4 TN cells gave rise to proportions of IL-17+ among CD4 T cells similar to those observed when Ly-6C− CD4 TN cells were injected alone (Supplementary Fig. S5). We then analysed, in this setting, the correlations between the ability of Ly-6C− CD4 TN cells to differentiate into iTreg cells and the capacity of co-injected Ly-6C− and Ly-6C+ CD4 TN cells to give rise to IL-17-producing CD4 T cells (Supplementary Fig. S6). Interestingly, the proportion of IL-17-producing CD4 T cells arising from initially injected Ly-6C− (CD45.2) and Ly-6C+ (CD45.1) CD4 TN cells similarly correlated with the percentage of iTreg cells generated from CD45.2+ Ly-6C− CD4 TN cells. These results agreed with our hypothesis that newly differentiated iTreg cells would favour TH17-cell differentiation. Finally, we conducted iTreg-cell-depletion experiments. Ly-6C− CD4 TN cells from DEREG mice26 were injected into T-cell-deficient recipients treated or not with diphtheria toxin (DT) twice a week (Fig. 3d). As expected, DT efficiently depleted iTreg cells (Supplementary Fig. S7a,b). More interestingly, DT treatment resulted in the reduction of the severity of the wasting disease (Fig. 3e) correlated with a significant decrease of the histological colitis score (Supplementary Fig. S7c). Depletion of iTreg cells also led to a strong inhibition of TH17-cell development (Fig. 3f). Taken together, our data strongly suggest that, in our experimental model, iTreg-cell differentiation promotes TH17-cell generation.
Efficient Ly-6C- CD4 TN-cell polarization into iTreg cells
Ly-6C− CD4 TN cells were more efficient in differentiating into iTreg cells than Ly-6C+ CD4 TN cells in vivo when transferred into a lymphopenic environment. To further assess whether this characteristic feature was an intrinsic property of Ly-6C− CD4 TN cells, we performed iTreg-cell polarization assays in vitro. Purified Ly-6C− or Ly-6C+ CD4 TN cells were stimulated with anti-CD3- and anti-CD28-coated antibodies (Abs) in the presence of graded doses of TGFβ. Although they expressed similar amounts of the TGFβ receptors I and II (Supplementary Fig. S8), Ly-6C− CD4 TN cells gave rise to a twofold higher proportion of iTreg cells than Ly-6C+ CD4 TN cells did for suboptimal doses of exogenous TGFβ (Fig. 4a). Differences were quantitative rather than qualitative, as iTregs cells generated from both Ly-6C− and Ly-6C+ CD4 TN cells displayed similar suppressive capacities (Fig. 4b). Two groups have recently described that suboptimal activation of CD4 TN cells in the absence of exogenous TGFβ led to their conversion into iTreg cells27,28. In this setting, a more efficient conversion was also observed in Ly-6C− CD4 TN-cell cultures (Supplementary Fig. S9).

(a) Purified LN Ly-6C− or Ly-6C+ CD4 TN cells from C57BL/6 Foxp3-GFP mice were stimulated for 4 days with coated anti-CD3 and anti-CD28 Abs in the presence of graded concentrations of TGFβ1. Representative Foxp3-GFP/CD4 dot-plots for gated CD4 T cells and the proportion of Foxp3+ cells among CD4 T cells are shown as a function of TGFβ1 concentration. (b) Purified Ly-6C− or Ly-6C+ CD4 TN cells were stimulated for 3 days with coated anti-CD3 and anti-CD28 Abs in the presence of 1 ng ml−1 of TGFβ1 and then let to rest for 3 more days in the presence of IL-2 (13 ng ml−1). GFP-expressing cells were then sorted using flow cytometry, and their suppressive abilities to inhibit the proliferation of Cell Trace Violet-labelled conventional CD4 T cells (LN GFP− CD4 T cells from C57BL/6 Foxp3-GFP mice) in response to anti-CD3 Abs in the presence of APCs were tested. (c) Ly-6C− or Ly-6C+ CD4 TN cells from C57BL/6 Foxp3-GFP mice were stimulated for 4 days with coated anti-CD3 and anti-CD28 Abs in the presence of IL-6 (20 ng ml−1) and graded concentrations of TGFβ1. Representative IL-17/Foxp3 dot-plots for gated CD4 T cells, the proportion of IL-17-producing CD4 T cells and the proportion of Foxp3+ cells among CD4 T cells are shown as a function of TGFβ1 concentration. (d) Ly-6C− or Ly-6C+ CD4 TN cells from C57BL/6 Foxp3-GFP mice were stimulated for 4 days with coated anti-CD3 and anti-CD28 Abs in the presence of graded concentrations of IL-12. Representative IL-17/IFNγ dot-plots for gated CD4 T cells and the proportion of IFNγ-producing CD4 T cells are shown as a function of IL-12 concentration. Results are representative of at least three independent experiments.
We then compared the capacity of Ly-6C− and Ly-6C+ CD4 TN cells to polarize into TH1 and TH17 cells in vitro (Fig. 4c,d). In response to low doses of TGFβ in the presence of IL-6, Ly-6C− and Ly-6C+ CD4 TN cells gave rise to similar proportions of TH17 cells. However, for high doses of TGFβ, Ly-6C− CD4 TN cells differentiated less efficiently into TH17 cells than their Ly-6C+-cell counterparts (Fig. 4c). This decreased ability of Ly-6C− CD4 TN cells to generate TH17 cells in response to high doses of TGFβ may derive from the fact that part of them differentiated into iTreg cells in these conditions despite the presence of IL-6. CD4 TN-cell polarization into TH1 cells did not differ whether or not cells were initially expressing Ly-6C (Fig. 4d). Taken together, our data strongly suggest that Ly-6C− CD4 TN cells have an intrinsic higher capacity to differentiate into iTreg cells than their Ly-6C+-cell counterparts, whereas both subsets differentiate with a similar efficiency into TH1 and TH17 cells.
Ly-6C expression is acquired in the periphery
We then decided to determine where and when Ly-6C expression was acquired by CD4 TN cells. To follow thymic output and specifically identify recent thymic emigrants in secondary lymphoid organs, we used recombination-activating gene 2 promoter (RAG2p)-green fluorescent protein (GFP) transgenic mice, in which GFP expression is driven by the RAG2p. The transgene generates a bright GFP signal during thymic differentiation that remains detectable in the periphery throughout the first few days after their migration29. Although most CD4 TN single-positive thymocytes were Ly-6C−, about 40% of CD4 TN recent thymic emigrants (GFPhi) in LNs already expressed Ly-6C molecule, suggesting that the acquisition of Ly-6C expression by CD4 TN cells occurs very rapidly after their exit from the thymus (Fig. 5a). Furthermore, the proportion of Ly-6C+ cells among peripheral CD4 TN lymphocytes increased with GFP brightness decay, indicating that acquisition of Ly-6C expression occurred in a rapid and gradual manner over time.

(a) GFP/Ly-6C contour plots and Ly-6C fluorescence histograms of gated CD4 TN cells recovered from the thymus and LNs of a representative C57BL/6 RAG2p-GFP reporter mouse. Numbers in each histogram represent the percentage of Ly-6C+ cells among the indicated CD4 TN cell subsets. (b) 1 × 106 thymic Ly-6C− CD4 TN cells from C57BL/6 Foxp3-GFP CD45.2 mice were injected intravenously (i.v.) into C57BL/6 CD45.1 mice lacking (MHC-II−) or not lacking (MHC-II+) MHC class II molecule expression. A diagram illustrating the experimental model and Ly-6C fluorescence histograms of gated CD45.2+ CD4 TN cells recovered from LNs of recipient mice 14 days after transfer are shown. (c) Ly-6C− (1 × 106) or Ly-6C+ CD4 TN cells from LNs of C57BL/6 Foxp3-GFP CD45.2 mice were injected i.v. into CD45.1 C57BL/6 mice lacking or not lacking MHC class II molecule expression. A diagram illustrating the experimental model is shown. The absolute numbers of CD45.2+ CD4 TN cells recovered from the periphery (LNs+spleen) of recipient mice 28 days after transfer are shown as means±s.e.m. Ly-6C fluorescence histograms of CD45.2+ CD4 TN cells recovered from LNs of recipient mice 14 and 28 days after transfer are shown. Results are representative of at least two independent experiments.
Then, we injected Ly-6C− CD4 TN single-positive thymocytes from CD45.2 mice into CD45.1 mice expressing or lacking major histocompatibility complex (MHC) class II molecules (Fig. 5b). As expected, Ly-6C− CD4 TN single-positive thymocytes gave rise to both Ly-6C− and Ly-6C+ CD4 TN cells after transfer into MHC class II-competent recipients. More surprisingly, all CD45.2+ CD4 TN cells recovered from MHC class II-deficient recipients were expressing Ly-6C, suggesting that all CD4 TN cells acquired Ly-6C expression or that survival of Ly-6C− CD4 TN cells was highly compromised in this context.
To address these issues, peripheral Ly-6C− or Ly-6C+ CD4 TN cells from CD45.2 mice were transferred into CD45.1 mice expressing or lacking MHC class II molecules (Fig. 5c). The survival of both Ly-6C− and Ly-6C+ CD45.2+ CD4 TN-cell subsets was affected by MHC class II molecule deprivation, excluding the hypothesis of a preferential disappearance of Ly-6C− CD4 TN cells in MHC class II-deficient recipients. Ly-6C expression by CD45.2+ CD4 TN cells was also assessed. First, we noticed that Ly-6C− and Ly-6C+ phenotypes were rather stable over time after transfer into MHC class II-competent recipients. One can notice that some CD4 TN cells, initially Ly-6C−, expressed low levels of this marker 14 days after transfer. However, this percentage remained relatively stable thereafter (33.1±2.1 on day 14 versus 37.3±1.9 on day 28, not significant). By contrast, virtually all peripheral CD45.2+ CD4 TN cells recovered from MHC class II-deficient recipients expressed Ly-6C whether or not they were expressing it before transfer. One may argue that MHC class II-deficient recipients lack not only MHC class II molecule expression but also endogenous CD4 T cells and that this latter point introduces another element of variability between both types of recipients. To address this issue, we generated BM chimeras in which CD4 T cells were produced in the thymus but were unable to interact with MHC class II molecules in the periphery (Supplementary Fig. S10a (refs 30, 31)). Although such chimeras contained as many peripheral CD4 T cells as control chimeras 4 weeks after BM cell transfer, almost all their CD4 TN cells were expressing Ly-6C in the periphery (Supplementary Fig. S10b, c). Altogether, these results suggest that the downmodulation of Ly-6C expression by CD4 TN cells is dependent on continuous interactions with MHC class II molecules.
Ly-6C expression is modulated by self-recognition
In line with a role for TCR signalling in the modulation of Ly-6C expression in CD4 TN cells, we found that in vitro Ly-6C+ CD4 TN-cell activation in response to TCR stimulation led to Ly-6C downregulation (Supplementary Fig. S11). Injection of CD4 TN cells into lymphopenic recipients (such as in the IBD model) led to the activation, the strong proliferation and the conversion into memory-like cells of part of the injected cells in response to self and environmental antigens, a process known as spontaneous proliferation32,33. This CD4 T-cell spontaneous proliferation relied on interactions with MHC class II molecules, and 2 weeks after transfer, most CD4 T cells recovered from the periphery of recipient mice derived from this process. Interestingly, the progeny of both Ly-6C− or Ly-6C+ CD4 TN cells was lacking Ly-6C expression 2 weeks after transfer into lymphopenic recipients (Supplementary Fig. S12). Thus, the TCR signals leading to T-cell activation induced Ly-6C downregulation both in vitro and in vivo.
We hypothesized that Ly-6C surface levels on CD4 TN cells might predict their propensity to interact with peripheral self-MHC class II molecules and thus to receive subthreshold TCR signals. CD5 expression on CD4 TN cells has been shown to be adjusted to reflect TCR contact with self34,35,36,37. In fact, the more a CD4 TN cell interacts with self, the more this cell expresses CD5. We thus analysed the expression of CD5, but also of CD4 and of the TCR itself by peripheral Ly-6C− and Ly-6C+ CD4 TN cells. Consistent with our hypothesis, we observed that Ly-6C− CD4 TN cells exhibited significantly higher densities of CD5 and lower levels of the TCRβ chain and CD4 compared with their Ly-6C+-cell counterparts (Fig. 6a). Mandl et al.37 have recently shown that the more CD4 TN cells expressed CD5, the more the ζ-chain of the CD3 complex was phosphorylated. In line with a higher capacity of Ly-6C− CD4 TN cells to interact with self, we found that the extent of CD3ζ phosphorylation was far more important in Ly-6C− CD4 TN cells than in Ly-6C+ CD4 TN cells (pζ/ζ ratio, Fig. 6b and Supplementary Fig. S11).

(a) CD5, TCRβ and CD4 fluorescence histograms of gated Ly-6C− and Ly-6C+ CD4 TN cells from LNs of a representative C57BL/6 Foxp3-GFP mouse. Mean fluorescence intensities (MFIs) are shown as means±s.e.m. for at least six mice from two independent experiments. (b) Flow-cytometry-sorted Ly-6C− and Ly-6C+ CD4 TN cells (3 × 105 cells/condition) were lysed, and both CD3ζ protein levels (ζ) and the extent of CD3ζ chain phosphorylation (pζ) were determined by immunoblotting. The pζ/ζ ratio was then quantified. Each dot represents an individual sample. (c,d) LN cells from C57BL/6 Foxp3-GFP mice were fixed immediately in 4% paraformaldehyde after isolation. Ly-6C− or Ly-6C+ CD4 TN cells were then purified and stained for CD4 (green) and ZAP-70 (red) expression. (c) Representative CD4 and ZAP-70 sum-projection images are shown. Scale bar, 5 μm. (d) CD4 polarization indexes were estimated for Ly-6C− (n=212) or Ly-6C+ (n=352) CD4 TN cells by automated analyses of two independent experiments (see Methods). (e) Distribution of Ly-6C− or Ly-6C+ CD4 TN cells as a function of their CD4 polarization index. (f–h) C57BL/6 Foxp3-GFP mice were injected or not intraperitoneally with anti-αL and anti-α4 Abs to block LN entry. Twenty-one hours later, LNs were recovered and CD4 T cells were analysed. (f) Diagram illustrating the experimental model. (g) Absolute numbers of CD4 T cells in mLNs. (h) Representative Ly-6C fluorescence histograms and percentages of Ly-6C− cells are shown for CD4 TN cells recovered from mLNs of treated and untreated mice. (i–j) T cells (15 × 106) from CD45.2 C57BL/6 Foxp3-GFP mice were transferred into C57BL/6 CD45.1 mice lacking (MHC-II−) or not lacking (MHC-II+) MHC class II molecule expression. Two hours later, LN entry was blocked. LNs were collected 21 h after entry blockade, and CD45.2+ CD4 T cells were analysed. (i) Diagram illustrating the experimental model. (j) Absolute numbers of CD45.2+ CD4 T cells in mLNs. Results are representative of at least two independent experiments. Significance of differences between two series of results was assessed using a two-tailed paired (b) or unpaired (d,g,h,j) Student’s t-test. Values of P<0.05 were considered as statistically significant (*P<0.05; **P<0.01; and ***P<0.001). ns, not significant.
We then decided to image Ly-6C− and Ly-6C+ CD4 TN cells. Stefanova et al.38 have shown that recognition of self-ligands by CD4 TN cells induced the clustering of the TCR complex and associated signalling molecules to the region of the lymphocyte membrane in contact with APCs. Here we found that LN CD4 TN cells directly fixed after isolation exhibited various degrees of clustering of both the CD4 co-receptor and ZAP-70 (Fig. 6c). We quantified CD4 polarization and found that the CD4 polarization index was significantly greater in Ly-6C− CD4 TN cells than in Ly-6C+ CD4 TN cells (Fig. 6d). More precisely, Ly-6C− CD4 TN cells contained a higher proportion of cells exhibiting high degree of CD4 polarization than their Ly-6C+-cell counterparts (Fig. 6e). Thus, at one given time, a higher proportion of Ly-6C− than Ly-6C+ CD4 TN cells might be interacting with APCs in LNs.
A greater ability of Ly-6C− CD4 TN cells to interact with self-MHC class II molecules should have consequences on their behaviour in secondary lymphoid organs. Mandl et al.36 have recently shown that interactions with MHC class II molecules retained CD4 TN cells in LNs. By using the same protocol (Fig. 6f), we analysed the phenotype of CD4 TN cells remaining within LNs after entry blockade. Most CD4 T cells have egressed from LNs 21 h after entry blockade (Fig. 6g). Interestingly, Ly-6C− cells were greatly enriched in the remaining CD4 TN cells (Fig. 6h). We then verified that retention of Ly-6C− CD4 TN cells in LNs after entry blockade was due to interactions with MHC class II molecules. To do so, we injected CD45.2+ T cells into CD45.1 recipients lacking (MHC-II−) or expressing (MHC-II+) MHC class II molecules. Two hours later, LN entry was blocked and LNs were collected 21 h after entry blockade (Fig. 6i). Although the absolute numbers of remaining Ly-6C+ CD4 TN cells were unaffected by MHC class II deprivation, those of Ly-6C− CD4 TN cells were significantly decreased in recipients lacking MHC class II molecules (Fig. 6j). This was not due to the acquisition of Ly-6C expression by Ly-6C− CD4 TN cells, as 24 h were not sufficient to induce upregulation of this marker (Supplementary Fig. S13). Thus, interactions with MHC class II molecules only retained Ly-6C− CD4 TN cells within LNs. These last results are in line with the greater average polarization of CD4 and ZAP-70 molecules at the cell surface of Ly-6C− CD4 TN cells and suggest that Ly-6C− CD4 TN cells are interacting more strongly than Ly-6C+ CD4 TN cells with MHC class II-expressing cells in peripheral secondary lymphoid organs.
Self-recognition reinforces iTreg-cell differentiation
We then determined whether self-recognition might have a role in the differential ability of Ly-6C− and Ly-6C+ CD4 TN cells to differentiate into iTreg cells. First, we studied the expression of CD5 and Ly-6C by peripheral monoclonal CD4 TN cells from AND and Marylin TCR-transgenic mice in a RAG2−/− background. It has recently been shown that AND CD4 T cells have a higher avidity for self than Marylin CD4 T cells37,39,40. Indeed, AND CD4 T cells expressed far higher density of CD5 than Marylin CD4 T cells (Fig. 7a (ref. 37)). In agreement with our assumption that Ly-6C non-expression might reflect the magnitude of CD4 TN-cell self-reactivity, all Marylin CD4 T cells were expressing high surface amounts of Ly-6C, whereas the majority of AND CD4 T cells did not (Fig. 7a). Finally, in line with our experiments with polyclonal CD4 TN cells, we observed that AND CD4 T cells differentiated far more efficiently than Marilyn CD4 T cells into iTreg cells in vitro (Fig. 7b).

(a) CD5 and Ly-6C fluorescence histograms and graph representing Ly-6C MFI of gated CD4 TN cells from LNs of AND (solid line/white) and Marylin (filled histogram/grey) TCR-transgenic mice as a function of their CD5 MFI. Each dot represents an individual mouse. (b) CD4 TN cells from LNs of AND and Marylin TCR-transgenic mice were stimulated for 4 days with coated anti-CD3 and anti-CD28 Abs in the presence of graded concentrations of TGFβ1. Representative Foxp3/CD4 dot-plots for gated CD4 T cells and the proportion of Foxp3+ cells among CD4 T cells are shown as a function of TGFβ1 concentration. (c–e) Ly-6C− (5 × 106) or Ly-6C+ CD4 TN cells from LNs of C57BL/6 Foxp3-GFP CD45.2 mice were injected i.v. into CD45.1 C57BL/6 mice lacking or not lacking MHC class II molecule expression (MHC-II− and MHC-II+, respectively). CD45.2+ CD4 TN cells were then FACS-sorted 21 days after transfer from the periphery of recipient mice and stimulated for 4 days with coated anti-CD3 and anti-CD28 Abs in the presence of 0.25 ng ml−1 of TGFβ1. (c) Diagram illustrating the experimental model. (d) Representative Foxp3-GFP/CD4 dot-plots for gated CD4 T cells are shown as a function of recipient mice and CD4 TN-cell origin. (e) Proportion of Foxp3+ cells among CD4 T cells as a function of recipient mice and CD4 TN cells origin are shown as means±s.e.m. for at least six mice from two independent experiments. Significance of differences between two series of results was assessed using a two-tailed unpaired Student’s t-test. Values of P<0.05 were considered as statistically significant (**P<0.01). ns, not significant.
Second, we adoptively transferred Ly-6C− and Ly-6C+ CD4 TN cells from CD45.2 mice into CD45.1 recipients expressing or lacking MHC class II molecules (Fig. 7c). Three weeks after transfer, CD45.2+ CD4 TN cells were purified from the periphery of recipient mice and assessed for their ability to differentiate into iTreg cells in vitro. For suboptimal doses of TGFβ, cells purified from MHC-II-competent recipients initially injected with Ly-6C− CD4 TN cells gave rise to a twofold higher proportion of iTreg cells than cells derived from MHC class II-deficient recipients initially injected with the same cells or from MHC-II-competent recipients initially injected with Ly-6C+ CD4 TN cells (Fig. 7d,e). Altogether, our data strongly suggest that the ability of a naive T cell to commit into the iTreg-cell lineage pathway upon in vitro stimulation is shaped by its capacity to interact with self in its original environment.
Discussion
Over the last decades, the CD4 TN cells have been commonly considered as a homogenous T-cell compartment. In the present paper, we confirm that Ly-6C expression splits this compartment into positive and negative cells with one-third of peripheral CD4 T cells lacking this marker20,21. Expression of Ly-6C is acquired in the periphery as almost all thymocytes are Ly-6C−. More precisely, analysis of RAG2p-GFP mice and adoptive transfer experiments indicate that thymocytes rapidly gain Ly-6C expression upon migration to the periphery. Then, the expression of Ly-6C by peripheral CD4 TN cells is stable over time. Moreover, several lines of evidence strongly suggest that the expression of Ly-6C by peripheral CD4 TN cells inversely correlates with their ability to interact with self. First, all thymic and peripheral CD4 TN cells gained Ly-6C expression when transferred into MHC class II-deficient recipients. Second, Ly-6C− CD4 TN cells expressed more CD5 and exhibited a higher degree of CD3ζ phosphorylation than Ly-6C+ CD4 TN cells. Third, Ly-6C− CD4 TN cells exhibit a higher degree of co-clustering of the CD4 co-receptor and the TCR-associated signalling molecule, ZAP-70, than their Ly-6C+-cell counterparts. Finally, interactions with MHC class II molecules retained preferentially Ly-6C− CD4 TN cells within LNs, therefore increasing their transit time. Thus, altogether, our data indicate that Ly-6C− CD4 TN cells are more ‘autoreactive’ than their Ly-6C+-cell counterparts.
Repeated subthreshold TCR stimulations are triggered within secondary lymphoid organs where T cells interact with self-peptides while scanning the surface of dendritic cells. Stefanova et al.38 have first shown that interruption of CD4 TN-cell contact with self leads to a rapid decline on signalling and response sensitivity to foreign stimuli. These data were further confirmed by several elegant studies and shown to apply also to CD8 TN cells41,42,43. Recently, using CD5 expression to discriminate between TN cells interacting strongly (CD5hi) or not (CD5lo) with self, several groups have shown that TCR contact with self in the periphery causes CD8 TN cells to be hypersensitive to IL-2, IL-7 and IL-15 (refs 44, 45, 46). Altogether, these results strongly suggest that self-recognition not only allows the survival of TN cells but also increase quantitatively their responsiveness towards their cognate antigens for both CD4 and CD8 TN cells, and towards ILs in the case of CD8 TN cells47. However, it has not been yet addressed whether self-recognition qualitatively affects the response of TN cells to stimulation. Here we propose that the ability of a CD4 TN cell to differentiate into an iTreg cell upon apropriate stimulation would be shaped by self-reactivity. In line with this assumption, Ly-6C− CD4 TN cells that are likely to interact strongly with MHC class II molecule-expressing cells in LNs are the cells that differentiate more efficiently into iTreg cells both in vitro and in vivo. Moreover, disruption of interactions with MHC class II molecules decreases the commitment efficacy of Ly-6C− CD4 TN cells towards the iTreg-cell lineage. Indeed, the iTreg-cell polarization potential of Ly-6C− CD4 TN cells parked for 3 weeks in MHC class II-deficient recipients dropped to the level observed in Ly-6C+ CD4 TN cells, the potential of which is not affected by MHC deprivation. Thus, self-recognition may affect not only quantitatively but also qualitatively the response of CD4 TN cells to their cognate antigens.
Upon activation, a CD4 TN cell can differentiate into a great variety of effector cells including TH1, TH2, TH17 or iTreg cells. The present paradigm assumes that the choice made by CD4 TN cells to commit into one of these lineages would be mainly governed by extrinsic factors with an important role of the cytokines released in the environment at the time of their activation. Several data, including those presented here, extend this theory by showing that intrinsic factors also have a role in CD4 TN-cell fate determination. Fifteen years ago, several studies have proposed that the strength of the TCR signal at the time of antigen recognition would have a role in the TH1/TH2 lineage choice48,49. More recently, Gottschalk et al.50 have shown that a low dose of a strong agonist resulted in maximal induction of iTreg cells. Thus, differentiation of CD4 TN cells into iTreg cells upon stimulation relies not only on extrinsic factors such as the presence of TGFβ in their environment but also on intrinsic factors such as the avidity of the TCR, which they express for both self and foreign ligands.
Altogether, our results strongly suggest that CD4 TN cells with the highest avidity for self (Ly-6C− CD4 TN cells) have a biased commitment towards the iTreg-cell lineage and that this intrinsic ability is linked to their enhanced pathogenicity when injected into lymphopenic mice. Indeed, colitis in mice transferred with Ly-6C− CD4 TN cells is prevented if Tregs are depleted. This last result may appear paradoxical, given the vast literature showing that Tregs prevent colitis in lymphopenic mice when co-transferred together with CD4 TN cells. The nature of Tregs by itself (natural versus induced Tregs), their number (iTregs represent from 4.7 to 11.4% of the CD4 T cells recovered from the secondary lymphoid organs of recipient mice at the end point of our experiments, whereas when co-injected at a physiological rate together with CD4 TN cells (one Treg for eight CD4 TN cells), they represent >25% of expanding peripheral CD4 T cells as soon as 1 week after transfer51) or the timing of the presence of Tregs in the periphery (from the beginning when co-transferred together with CD4 TN cells versus the time required for their generation and expansion in our setting) may explain these opposite effects of Tregs in the IBD experimental model.
In other settings, in particular in lymphoreplete animals, an enhanced ability of the most autoreactive CD4 TN cells to commit to the iTreg-cell lineage pathway might represent a previously undescribed mechanism of self-tolerance. This process would diminish the risk of the most self-reactive CD4 T cells to become deleterious effector cells through responding to bacteria, viruses or parasites. Indeed, such a mechanism would allow the organism to augment its own protection during each infectious episode, especially by avoiding molecular mimics to induce novel, potent and self-peptide-reactive CD4 T-cell effectors when T-cell responses raised against the pathogen cross-react with self-peptides.
Methods
Mice
C57BL/6 mice (CD45.2) were obtained from Charles River Laboratories. C57BL/6 CD45.1, C57BL/6 CD3ε−/−, C57BL/6 MHC-IIΔ/Δ CD45.1 and C57BL/6 AND TCR-transgenic RAG2−/− mice were maintained in our own animal facilities, under specific pathogen-free conditions. C57BL/6 Marilyn TCR-transgenic RAG2−/− mice were provided by Dr Emmanuel Donnadieu, Institut Cochin, Paris, France. C57BL/6 Foxp3-GFP CD45.252 and C57BL/6 DEREG26 mice were initially obtained from Dr Bernard Malissen, Centre d'Immunologie de Marseille-Luminy, France, and Dr Tim Sparwasser, Institute of Infection Immunology, TWINCORE, Hannover, Germany, respectively. C57BL/6 Foxp3-GFP CD45.2 mice were then crossed with C57BL/6 CD45.1 mice to generate C57BL/6 Foxp3-GFP CD45.1 mice. C57BL/6 RAG2p-GFP reporter mice53 were provided by Dr Antonio Bandeira, Unité du Développement des Lymphocytes, Institut Pasteur, Paris, France. Four- to 12-week-old mice were used for all experiments. Donor and recipient mice were sex-matched. Experiments were carried out in accordance with the guidelines of the French Veterinary Department. All procedures performed were approved by the Paris-Descartes Ethical Committee for Animal Experimentation (decision CEEA34.CA.080.12).
Cell suspensions
pLNs, mLNs, spleen and thymus were homogenized and passed through a nylon cell strainer (BD Falcon) in RPMI 1640 Glutamax (Gibco) supplemented with 10% FCS (Biochrom) for adoptive transfer (LNs only), or in 5% FCS and 0.1% NaN3 (Sigma-Aldrich) in PBS for flow cytometry.
Adoptive transfer of CD4 TN cells
CD4 T cells were purified from LNs (pooled superficial cervical, axillary, brachial, inguinal and mLNs) or thymi of C57BL/6 Foxp3-GFP or DEREG mice by incubating cell suspensions on ice for 20 min with a mixture of anti-CD8 (53-6.7), anti-CD11b (Mac-1) and anti-CD19 (1D3) Abs obtained from hybridoma supernatants, and then with magnetic beads coupled to anti-rat immunoglobulins (Dynal Biotech). Purified CD4 T cells were then labelled with biotinylated anti-Ly-6C (AL-21), PE-conjugated anti-CD25 (PC61), APC-conjugated anti-CD44 (IM7), all from BD Biosciences, and Pacific Blue-conjugated streptavidin (Invitrogen). Ly-6C− and Ly-6C+ CD4 TN cells were sorted as GFP− CD25− CD44−/lo cells using a FACS-ARIA3 flow cytometer (BD Biosciences) and injected intravenously into sex-matched recipient mice. In the experiments depicted in Fig. 3d–f and Supplementary Fig. 7, recipient mice were injected intraperitoneally 2 consecutive days per week with 500 ng of DT (Sigma-Aldrich).
Adoptive transfer of BM cells
BM cells were incubated on ice for 20 min with anti-CD4 (GK1.5) and anti-CD8a (53-6.7) Abs, obtained from hybridoma supernatants, and then with magnetic beads coupled to anti-rat immunoglobulin (Dynal Biotech). We injected 5 × 106 T-cell-depleted BM cells intravenously into each lethally irradiated (9.5 Gy) recipient mouse.
Cell surface staining and flow cytometry
Cell suspensions were collected and dispensed into 96-well round-bottom microtiter plates (Greiner Bioscience; 6 × 106 cells per well). Surface staining was performed by incubating the cells on ice, for 15 min per step, with Abs in 5% FCS (Biochrom) and 0.1% NaN3 (Sigma-Aldrich) PBS. Each cell-staining reaction was preceded by a 15-min incubation with a purified anti-mouse CD16/32 Ab (FcγRII/III block; 2.4G2) obtained from hybridoma supernatants. For determination of intracellular cytokine production, cells were stimulated with 0.5 μg ml−1 phorbol myristate acetate (Sigma-Aldrich), 0.5 μg ml−1 ionomycin (Sigma-Aldrich) and 10 μg ml−1 Brefeldin A (Sigma-Aldrich) for 2 h at 37 °C. Cells were then stained for surface markers, fixed in 2% paraformaldehyde in PBS and permeabilized with 0.5% saponin, followed by labelling with specific cytokine Abs.
PerCP-conjugated anti-CD4 (RM4-5), phycoerythrin (PE)-conjugated anti-CD5 (53-7.3), anti-CD25 (PC61), anti-CD45.1 (A20), anti-CD69 (H1.2F3), anti-TCRβ (H57-597), anti-IL-2 (JES6-5H4), anti-IL-4 (11B11), anti-IL-10 (JES5-16E3) and anti-IL-17A (TC11-18H10), APC-conjugated anti-CD44 (IM7), anti-TCRβ (H57-597), anti-IFN-γ (XMG1.2), streptavidin, fluorescein isothiocyanate-conjugated anti-TNF-α (MP6-XT22), PE-Cy7-conjugated anti-Ly-6C (AL-21), streptavidin, PerCP-Cy5.5-conjugated anti-CD45.2 (104), APC-H7-conjugated anti-CD8 (53-6.7), Pacific Blue-conjugated anti-CD4 (RM4-5), and Alexa Fluor 700-conjugated anti-Ly-6C (AL-21), biotinylated anti-Ly-6C (AL-21), anti-CD5 (53-6.7), anti-CD45.1 (A20), anti-CD45.2 (104), anti-CD45RB (16A) and anti-CD62L (MEL14) were obtained from BD Biosciences. PE-conjugated anti-CD45.2 (104), PE-Cy5-conjugated anti-CD25 (PC61.5), Alexa Fluor 700-conjugated anti-CD45.2 (104), PerCP-Cy5.5-conjugated anti-TCRβ (H57-597) and biotinylated anti-CD127 (A7R34) were obtained from eBioscience. Pacific Blue-conjugated streptavidin was obtained from Invitrogen.
Multi-colour immunofluorescence was analysed using a BD-LSR2 cytometer (BD Biosciences). List-mode data files were analysed using Diva software (BD Biosciences). Data acquisition and cell sorting were performed on the Cochin Immunobiology facility.
Immunoblot analysis
Ly-6C− and Ly-6C+ CD4 TN cells (3 × 105 cells per condition) were flow-cytometry sorted as described above, washed in ice-cold PBS and lysed in lysis buffer (20 mM Tris-HCl (pH 7.5), 150 mM NaCl, 5 mM EDTA (pH 8.0), 5 mM NaPiP, 1 mM, Na3VO4, 20 mM NaPO4 (pH 7.6), 3 mM β-glycerophosphate, 10 mM NaF, 1% Triton X-100, complete protease and phosphatase inhibitor cocktail (Roche)) for 15 min on ice. Samples were centrifugated, and whole-cell lysates were collected. Unreduced proteins were separated on a 12% SDS-PAGE, transferred on a nitrocellulose membrane and immunoblotted first with an anti-Phosphotyrosine Ab (Clone 4G10; Millipore) and, after stripping, with an anti-CD3ζ Ab (Clone 6B10.2; Santa Cruz Biotechnology). Signals were quantified using the Image J software. The ratio pζ/ζ was estimated as previously described54.
Immunofluorescence microscopy
Freshly isolated LN cells were first fixed in 4% paraformaldehyde in PBS for 5 min at 37 °C. Ly-6C− and Ly-6C+ CD4 TN cells were then sorted using flow cytometry as described above and stained with fluorescein isothiocyanate-conjugated anti-CD4 Ab. T cells were then permeabilized and incubated with an anti-ZAP-70 Ab (Transduction Laboratories) followed by anti-mouse Ab conjugated to Alexa Fluor 647 (Molecular Probes). Stained T cells were photographed on a Yokogawa spinning-disk confocal microscope with a × 100 oil immersion objective connected to a digital video camera and analysed using Image J software. Briefly, automated analysis was performed to divide each cell into quadrants and then to measure the average intensity in each quadrant. The CD4 polarization index was calculated by dividing the highest quadrant by the lowest.
Colitis scoring
For the colitis model, colons were removed and fixed in PBS containing 10% formaldehyde. Five-micrometer paraffin-embedded sections were cut and stained with haematoxylin and eosin and then blindly analysed. Each segment was given a score of 0–4: grade 0, no significant changes; grade 1, minimal scattered mucosal inflammatory cell infiltrates, with or without minimal epithelial hyperplasia; grade 2, mild scattered to diffuse inflammatory cell infiltrates, sometimes extending into the submucosa and associated with erosions, with mild to moderate epithelial hyperplasia and mild to moderate mucin depletion from goblet cells; grade 3, moderate inflammatory cell infiltrates that were sometimes transmural, with moderate to severe epithelial hyperplasia and mucin depletion; grade 4, marked inflammatory cell infiltrates that were often transmural and associated with crypt abscesses and occasional ulceration, with marked epithelial hyperplasia, mucin depletion and loss of intestinal glands.
In vitro polarization assays
Flow-cytometry sorted Ly-6C− and Ly-6C+ CD4 TN cells from LNs of C57BL/6 Foxp3-GFP mice were stimulated for 4 days with immobilized anti-CD3 (clone 145.2C11; 4 μg ml−1; obtained from hybridoma supernatants) and anti-CD28 (clone 37.51; eBioscience; 4 μg ml−1) Abs, in the presence of graded concentrations of exogenous recombinant human TGFβ1 (Invitrogen) in the presence (TH17) or absence (iTreg) of 20 ng ml−1 of recombinant mouse IL-6 (Invitrogen). To study the ability of CD4 TN cells to differentiate into TH1 effector cells, stimulation was performed in the presence of graded concentrations of recombinant mouse IL-12 (R&D).
In vitro suppression assay
Purified Ly-6C− or Ly-6C+ CD4 TN cells were stimulated for 3 days with coated anti-CD3 and anti-CD28 Abs in the presence of 1 ng ml−1 of TGFβ1 and then let to rest for 3 more days in the presence of recombinant human IL-2 (13 ng ml−1; R&D). GFP-expressing cells were then sorted using flow cytometry, and the suppressive capacities of these highly purified Foxp3-expressing cells were then assessed as previously described31. Briefly, conventional CD4 T cells (GFP− CD4+ T cells) were purified from LNs of C57BL/6 Foxp3-GFP mice, labelled with Cell Trace Violet (Invitrogen) and stimulated for 64 h, alone or together with Treg cells at various Treg/Tconv-cell ratios.
Suboptimal T-cell activation in vitro assay
Flow-cytometry sorted Ly-6C− and Ly-6C+ CD4 TN cells from LNs of C57BL/6 Foxp3-GFP were stimulated for 3 days with irradiated splenocytes from C57BL/6 CD3ε−/− mice in the presence of graded concentrations of soluble anti-CD3 Ab (145.2C11). These experiments were conducted in the absence of addition of exogenous cytokines.
Blocking T-cell entry into LNs
C57BL/6 Foxp3-GFP mice were injected or not intraperitoneally with 200 μg of anti-αL and anti-α4 Abs (Bio-Xcell) to block LN entry. Twenty-one hours later, LNs were collected, and CD4 T cells were analysed. In some experiments (Fig. 6i,j), 15 × 106 T cells from CD45.2 C57BL/6 Foxp3-GFP mice were transferred into C57BL/6 CD45.1 mice lacking (MHC-II−) or not lacking (MHC-II+) MHC class II molecule expression. Two hours later, LN entry was blocked as described above. LNs were collected 21 h after entry blockade, and CD45.2+ CD4 T cells were analysed.
Additional information
How to cite this article: Martin, B. et al. Highly self-reactive naive CD4 T cells are prone to differentiate into regulatory T cells. Nat. Commun. 4:2209 doi: 10.1038/ncomms3209 (2013).
References
Mosmann, T. R., Cherwinski, H., Bond, M. W., Giedlin, M. A. & Coffman, R. L. Two types of murine helper T cell clone. I. Definition according to profiles of lymphokine activities and secreted proteins. J. Immunol. 136, 2348–2357 (1986).
Abbas, A. K., Murphy, K. M. & Sher, A. Functional diversity of helper T lymphocytes. Nature 383, 787–793 (1996).
Ye, P. et al. Requirement of interleukin 17 receptor signaling for lung CXC chemokine and granulocyte colony-stimulating factor expression, neutrophil recruitment, and host defense. J. Exp. Med. 194, 519–527 (2001).
Liang, S. C. et al. Interleukin (IL)-22 and IL-17 are coexpressed by Th17 cells and cooperatively enhance expression of antimicrobial peptides. J. Exp. Med. 203, 2271–2279 (2006).
Bilate, A. M. & Lafaille, J. J. Induced CD4(+)Foxp3(+) regulatory T cells in immune tolerance. Annu. Rev. Immunol. 30, 733–758 (2012).
Szabo, S. J. et al. A novel transcription factor, T-bet, directs Th1 lineage commitment. Cell 100, 655–669 (2000).
Zheng, W. & Flavell, R. A. The transcription factor GATA-3 is necessary and sufficient for Th2 cytokine gene expression in CD4 T cells. Cell 89, 587–596 (1997).
Ivanov, I. I. et al. The orphan nuclear receptor RORgammat directs the differentiation program of proinflammatory IL-17+ T helper cells. Cell 126, 1121–1133 (2006).
Hori, S., Nomura, T. & Sakaguchi, S. Control of regulatory T-cell development by the transcription factor Foxp3. Science 299, 1057–1061 (2003).
Fontenot, J. D., Gavin, M. A. & Rudensky, A. Y. Foxp3 programs the development and function of CD4+CD25+ regulatory T cells. Nat. Immunol. 4, 330–336 (2003).
Groux, H. et al. A CD4+ T-cell subset inhibits antigen-specific T-cell responses and prevents colitis. Nature 389, 737–742 (1997).
Veldhoen, M. et al. Transforming growth factor-beta 'reprograms' the differentiation of T helper 2 cells and promotes an interleukin 9-producing subset. Nat. Immunol. 9, 1341–1346 (2008).
Nurieva, R. I. et al. Generation of T follicular helper cells is mediated by interleukin-21 but independent of T helper 1, 2, or 17 cell lineages. Immunity 29, 138–149 (2008).
Hsieh, C. S. et al. Development of TH1 CD4+ T cells through IL-12 produced by Listeria-induced macrophages. Science 260, 547–549 (1993).
Kopf, M. et al. Disruption of the murine IL-4 gene blocks Th2 cytokine responses. Nature 362, 245–248 (1993).
Chen, W. et al. Conversion of peripheral CD4+CD25- naive T cells to CD4+CD25+ regulatory T cells by TGF-beta induction of transcription factor Foxp3. J. Exp. Med. 198, 1875–1886 (2003).
Veldhoen, M., Hocking, R. J., Atkins, C. J., Locksley, R. M. & Stockinger, B. TGFbeta in the context of an inflammatory cytokine milieu supports de novo differentiation of IL-17-producing T cells. Immunity 24, 179–189 (2006).
Bettelli, E. et al. Reciprocal developmental pathways for the generation of pathogenic effector TH17 and regulatory T cells. Nature 441, 235–238 (2006).
Corse, E., Gottschalk, R. A. & Allison, J. P. Strength of TCR-peptide/MHC interactions and in vivo T cell responses. J. Immunol. 186, 5039–5045 (2011).
Schlueter, A. J. et al. Distribution of Ly-6C on lymphocyte subsets: I. Influence of allotype on T lymphocyte expression. J. Immunol. 158, 4211–4222 (1997).
McHeyzer-Williams, L. J. & McHeyzer-Williams, M. G. Developmentally distinct Th cells control plasma cell production in vivo. Immunity 20, 231–242 (2004).
Powrie, F., Leach, M. W., Mauze, S., Caddle, L. B. & Coffman, R. L. Phenotypically distinct subsets of CD4+ T cells induce or protect from chronic intestinal inflammation in C. B-17 scid mice. Int. Immunol. 5, 1461–1471 (1993).
Martin, B. et al. Suppression of CD4+ T lymphocyte effector functions by CD4+CD25+ cells in vivo. J. Immunol. 172, 3391–3398 (2004).
Pandiyan, P. et al. CD4(+)CD25(+)Foxp3(+) regulatory T cells promote Th17 cells in vitro and enhance host resistance in mouse Candida albicans Th17 cell infection model. Immunity 34, 422–434 (2011).
Chen, Y. et al. Foxp3(+) regulatory T cells promote T helper 17 cell development in vivo through regulation of interleukin-2. Immunity 34, 409–421 (2011).
Lahl, K. et al. Selective depletion of Foxp3+ regulatory T cells induces a scurfy-like disease. J. Exp. Med. 204, 57–63 (2007).
Gabrysova, L. et al. Integrated T-cell receptor and costimulatory signals determine TGF-beta-dependent differentiation and maintenance of Foxp3+ regulatory T cells. Eur. J. Immunol. 41, 1242–1248 (2011).
Oliveira, V. G., Caridade, M., Paiva, R. S., Demengeot, J. & Graca, L. Sub-optimal CD4+ T-cell activation triggers autonomous TGF-beta-dependent conversion to Foxp3+ regulatory T cells. Eur. J. Immunol. 41, 1249–1255 (2011).
Boursalian, T. E., Golob, J., Soper, D. M., Cooper, C. J. & Fink, P. J. Continued maturation of thymic emigrants in the periphery. Nat. Immunol. 5, 418–425 (2004).
Poitrasson-Riviere, M. et al. Regulatory CD4+ T cells are crucial for preventing CD8+ T cell-mediated autoimmunity. J. Immunol. 180, 7294–7304 (2008).
Delpoux, A. et al. Foxp3-independent loss of regulatory CD4(+) T-cell suppressive capacities induced by self-deprivation. Eur. J. Immunol. 42, 1237–1249 (2012).
Martin, B., Becourt, C., Bienvenu, B. & Lucas, B. Self-recognition is crucial for maintaining the peripheral CD4+ T-cell pool in a nonlymphopenic environment. Blood 108, 270–277 (2006).
Le Campion, A. et al. Lymphopenia-induced spontaneous T-cell proliferation as a cofactor for autoimmune disease development. Blood 114, 1784–1793 (2009).
Azzam, H. S. et al. CD5 expression is developmentally regulated by T cell receptor (TCR) signals and TCR avidity. J. Exp. Med. 188, 2301–2311 (1998).
Smith, K. et al. Sensory adaptation in naive peripheral CD4 T cells. J. Exp. Med. 194, 1253–1261 (2001).
Mandl, J. N. et al. Quantification of lymph node transit times reveals differences in antigen surveillance strategies of naive CD4+ and CD8+ T cells. Proc. Natl Acad. Sci. USA 109, 18036–18041 (2012).
Mandl, J. N., Monteiro, J. P., Vrisekoop, N. & Germain, R. N. T cell-positive selection uses self-ligand binding strength to optimize repertoire recognition of foreign antigens. Immunity 38, 263–274 (2012).
Stefanova, I., Dorfman, J. R. & Germain, R. N. Self-recognition promotes the foreign antigen sensitivity of naive T lymphocytes. Nature 420, 429–434 (2002).
Grandjean, I. et al. Are major histocompatibility complex molecules involved in the survival of naive CD4+ T cells? J. Exp. Med. 198, 1089–1102 (2003).
Kassiotis, G., Zamoyska, R. & Stockinger, B. Involvement of avidity for major histocompatibility complex in homeostasis of naive and memory T cells. J. Exp. Med. 197, 1007–1016 (2003).
Lo, W. L. et al. An endogenous peptide positively selects and augments the activation and survival of peripheral CD4+ T cells. Nat. Immunol. 10, 1155–1161 (2009).
Ebert, P. J., Jiang, S., Xie, J., Li, Q. J. & Davis, M. M. An endogenous positively selecting peptide enhances mature T cell responses and becomes an autoantigen in the absence of microRNA miR-181a. Nat. Immunol. 10, 1162–1169 (2009).
Hochweller, K. et al. Dendritic cells control T cell tonic signaling required for responsiveness to foreign antigen. Proc. Natl Acad. Sci. USA 107, 5931–5936 (2010).
Cho, J. H., Kim, H. O., Surh, C. D. & Sprent, J. T cell receptor-dependent regulation of lipid rafts controls naive CD8+ T cell homeostasis. Immunity 32, 214–226 (2010).
Stoklasek, T. A., Colpitts, S. L., Smilowitz, H. M. & Lefrancois, L. MHC class I and TCR avidity control the CD8 T cell response to IL-15/IL-15Ralpha complex. J. Immunol. 185, 6857–6865 (2010).
Palmer, M. J., Mahajan, V. S., Chen, J., Irvine, D. J. & Lauffenburger, D. A. Signaling thresholds govern heterogeneity in IL-7-receptor-mediated responses of naive CD8(+) T cells. Immunol. Cell. Biol. 89, 581–594 (2011).
Garbi, N., Hammerling, G. J., Probst, H. C. & van den Broek, M. Tonic T cell signalling and T cell tolerance as opposite effects of self-recognition on dendritic cells. Curr. Opin. Immunol. 22, 601–608 (2010).
Hosken, N. A., Shibuya, K., Heath, A. W., Murphy, K. M. & O'Garra, A. The effect of antigen dose on CD4+ T helper cell phenotype development in a T cell receptor-alpha beta-transgenic model. J. Exp. Med. 182, 1579–1584 (1995).
Tao, X., Constant, S., Jorritsma, P. & Bottomly, K. Strength of TCR signal determines the costimulatory requirements for Th1 and Th2 CD4+ T cell differentiation. J. Immunol. 159, 5956–5963 (1997).
Gottschalk, R. A., Corse, E. & Allison, J. P. TCR ligand density and affinity determine peripheral induction of Foxp3 in vivo. J. Exp. Med. 207, 1701–1711 (2010).
Le Campion, A. et al. IL-2 and IL-7 determine the homeostatic balance between the regulatory and conventional CD4+ T cell compartments during peripheral T cell reconstitution. J. Immunol. 189, 3339–3346 (2012).
Wang, Y. et al. Th2 lymphoproliferative disorder of LatY136F mutant mice unfolds independently of TCR-MHC engagement and is insensitive to the action of Foxp3+ regulatory T cells. J. Immunol. 180, 1565–1575 (2008).
Yu, W. et al. Continued RAG expression in late stages of B cell development and no apparent re-induction after immunization. Nature 400, 682–687 (1999).
Dorfman, J. R., Stefanova, I., Yasutomo, K. & Germain, R. N. CD4+ T cell survival is not directly linked to self-MHC-induced TCR signaling. Nat. Immunol. 1, 329–335 (2000).
Acknowledgements
We greatly acknowledge K. Labroquère, L. Stouvenel and M. Andrieu from the Cochin Immunobiology facility, and B. Durel and P. Bourdoncle from the Cochin Cellular Imaging facility. We would like to thank A. Bandeira, E. Donnadieu, C. A. Dutertre and A. Henino for providing tools and advice, and T. Guilbert for his invaluable help in image analysis. This work was supported by grants from the ‘Ligue contre le Cancer’ and the ‘Association pour la Recherche contre le Cancer’. A.P. was supported by a PhD fellowship from the ‘Association pour la Recherche sur le Cancer’. A.A. was supported by a master fellowship from the ‘Fondation pour la Recherche Médicale’.
Author information
Authors and Affiliations
Contributions
Br.M., C.A., B.S. and B.L. designed experiments. Br.M., C.A., A.D. A.P., C.C., P.Y., H.d.B., N.B. and A.A. did the experiments. T.S. and Be.M. provided tools. Br.M., C.A. and B.L. wrote the paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Supplementary Information
Supplementary Figures S1-S13 (PDF 1307 kb)
Rights and permissions
About this article
Cite this article
Martin, B., Auffray, C., Delpoux, A. et al. Highly self-reactive naive CD4 T cells are prone to differentiate into regulatory T cells. Nat Commun 4, 2209 (2013). https://doi.org/10.1038/ncomms3209
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/ncomms3209
This article is cited by
-
Type 1 interferons and Foxo1 down-regulation play a key role in age-related T-cell exhaustion in mice
Nature Communications (2024)
-
TCR signal strength controls thymic differentiation of iNKT cell subsets
Nature Communications (2018)
-
Stepwise B-cell-dependent expansion of T helper clonotypes diversifies the T-cell response
Nature Communications (2016)
-
Positive and negative selection of the T cell repertoire: what thymocytes see (and don't see)
Nature Reviews Immunology (2014)
-
Intrinsic CD4+ T cell sensitivity and response to a pathogen are set and sustained by avidity for thymic and peripheral complexes of self peptide and MHC
Nature Immunology (2014)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
