
Abstract
It is widely assumed that mineral inclusions and their host diamonds are ‘syngenetic’ in origin, which means that they formed simultaneously and from the same chemical processes. Mineral inclusions that, instead, were formed earlier with respect to diamonds are termed protogenetic. However, minerals can have the same age as the diamonds in that they become enclosed in and isolated from any further isotopic exchange. But this is termed ‘synchronous’ not ‘syngenetic’. Here we demonstrate conclusively the protogenesis of inclusions in diamonds, based upon data from an exceptional fragment of a diamond-bearing peridotite, its clinopyroxene and a gem-quality diamond. Clinopyroxenes in the xenolith had the same chemistry and crystallographic orientation as those for inclusions in the diamond. With our results with garnets, olivines and sulfides, we can state that a major portion of the mineral inclusions in non-coated, monocrystalline-lithospheric diamonds are protogenetic. Our discovery here presented has implications for all genetic aspects of diamond growth, including their ages.
Similar content being viewed by others

Introduction
Diamonds and their mineral inclusions are among the most intriguing natural samples on Earth, as they represent a unique opportunity to probe the deepest regions of our planet. Indeed, diamonds are thought to be among the oldest available materials—the oldest diamonds have been dated at up to 3.5 billion years1,2,3,4,5). In addition, diamond is also able to capture mineral inclusions at depths from even greater than 500 km6,7. Such a combination makes diamond and its pristine mineral inclusions a virtual ‘window into the Earth’s mantle’, providing crucial information about the geological evolution of our planet. However, it is also well known and appreciated that the age and crystallization environment of diamond are only based upon its mineral inclusions; indeed, the age of diamonds are determined by dating the mineral inclusions trapped within them; similarly, the depth of diamond formation is determined by studying the depth of formation of the inclusions8,9,10.
The validity of the criteria for the ‘syngenetic’ co-crystallization of diamonds and their inclusions is at the very foundation of all diamond inclusion studies. Virtually every paper on mineral inclusions in diamonds is based upon an essential assumption that the mineral inclusions and their host diamond are syngenetic—that is, formed simultaneously and from the same formational process. However, supporting evidence for this assumption is rarely presented. There have been an extensive series of papers written with reviews of the criteria for ‘syngenesis’ of mineral inclusions and their host diamonds11,12,13,14, each coming to the conclusion that most, if not all, diamonds and their inclusions are syngenetic. Others believe the opposite—that most, if not all, inclusions in diamonds are protogenetic—formed before encapsulation in the diamonds15,16,17,18,19,20,21,22,23,24. The syngenesis versus protogenesis relationship between diamond and its mineral inclusions represents an extended scientific debate in diamond research; it is likely that the many decade-years old debate has not been well-addressed, at least, until recently. In general, we can count only a few publications where protogenesis is really invoked15,16,17,18,19,20,21,22,23,24. This illustrates how crucial the principle of ‘syngenesis’ is in diamond research. We will address the criteria for establishing the syngenesis of diamond and its mineral inclusions. This will be placed within the context of the evidence, presented in this paper, as well as in prior studies, that most mineral inclusions in diamonds are simply enclosed, pre-existing minerals—that is, protogenetic—and are synchronous, not syngenetic.
In this study, based upon the morphology, crystallography and chemistry of mineral inclusions in non-coated, monocrystalline-lithospheric diamonds, we present unequivocal evidence for the protogenesis of these inclusions, not syngenesis. In addition, we address the popular use of the term ‘syngenesis’ for such inclusions. Some mineral inclusions may be ‘synchronous’ (e.g., monosulfide solid solution); however, virtually no silicate inclusions are truly syngeneous—representing the same radiogenic isotopic ages. However, the possibility of syngenesis can still permit the age presented by the mineral inclusion to represent the same age as its diamond host.
Results
Syngenesis versus protogenesis and the case of diopsid
The two main arguments in favour of syngenesis are focused on the morphology of the inclusions, which often show a pseudocubo-octahedral morphology imposed by the diamond11,25,26, and on the assumption that diamond and its mineral inclusions show epitaxial growth relationships (coincidence of crystallographic planes and directions between the diamond and its inclusions12,13,14,27). There are other arguments based upon C and O isotopes, in coexisting diamond and silicate inclusions, which have been made by Ickert et al.28, but this approach is a subject of considerable debate29 and will not be addressed further.
Relative to the inclusion morphologies, recent studies24,30,31 reported strong evidences for protogenesis of olivines included in diamonds from the Siberian (Russia) and Kaapvaal (South Africa) cratons. This is in spite of the olivines having pseudocubo-octahedral morphology. Indeed, the cubo-octahedral imposed morphology by diamond is extremely common for all types of inclusions and such a morphology has always been considered a strong argument in favour of syngenesis. However, this cubic super-imposed morphology, reported to indicate simultaneous crystallization from same fluid at the same time13, has no experimental and conclusive proof of this contention in spite of continued reference to Sunagawa32,33.
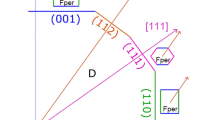
Relatively to the reported ‘epitaxy’ of diamond and its inclusionsrecent studies24,31 by single-crystal X-ray diffraction definitively showed that no epitaxial relationship between olivine and diamond was discovered on a huge population of inclusions in diamonds from different localities. Indeed, one of the most interesting discoveries, relative to olivine inclusions, is that they are totally randomly oriented within their diamond host. However, in single diamonds, with multiple olivine inclusions, some olivine crystals have an identical crystallographic orientation to each other24,31, yet not to their diamond hosts, as shown in Fig. 1 and represented in a cartoon in Fig. 2. This experimental evidence was interpreted as the main argument in favour of the protogenetic origin of such olivines, with respect their diamond hosts. The explanation for this conclusion was that the iso-oriented crystals could be only interpreted as the remaining ‘islands’ of a pre-existing olivine mono-crystal after a resorption process involved in the diamond crystallization. Additional experimental evidence for protogenesis is statistical in that of all the diamond hosts and the inclusions investigated (28 diamonds and 63 olivine inclusions), the authors24,31 could not find a single diamond with any set of crystallographic orientations in common with the olivine inclusion. Indeed, some diamonds contained up to seven olivine inclusions. Their interpretation was explained in terms of the crystallization of the diamond, which would be energetically favoured at a triple junction between pre-existing olivines in the peridotite host-rock, during introduction of the metasomatic fluid that formed the diamond. However, this is only an interpretation, and until the present study, quantitative evidence has been lacking to demonstrate it.

The diamond shows seven different mineral inclusions, modified after Milani et al.31. Six inclusions are olivines and the seventh one is a crystal of magnesio-chromite. The six inclusions of olivine belong to two different groups, group I and group II, respectively. Each group shows identical crystallographic orientation. The different colour of inclusion PR1-3 is likely due to an optical effect, due to a greater thickness. The inclusions, indeed, have all the same chemical compositions, as demonstrated by the identical unit-cell parameters31.

The cartoon in the figure shows a possible diamond–olivine growth relationship, as proposed by and modified from previous studies24,30. The host diamond is in yellow; the shapes of the olivine crystals used in this cartoon are not real but are simplified. The cartoon explains the experimental observations from different works in which multiple inclusions of olivines show different crystallographic orientations but at the same time, in the same diamond, it is also possible to find different olivines with similar crystallographic orientation. This orientational similarity can be obtained only if these olivines belonged to the same pre-existing olivine monocrystal. This is strong evidence of olivine protogenesis.
Evidence of protogenesis
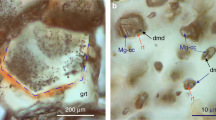
We have investigated, by single-crystal X-ray diffraction (hereafter XRD), a coarse-grained diamond-bearing garnet peridotite xenolith from the Finsch mine (Kaapvaal craton, South Africa). This peridotite contains a large millimetre-sized, gem-quality, octahedral diamond, still embedded in its host rock. This portion of the xenolith comes from the same sample34 and is shown in Fig. 3. The gem-quality octahedral diamond is well visible with its vertex pointing away from the image and is embedded in a matrix of serpentine, orthopyroxene, garnet and clinopyroxene. Owing to the intersection of the planar-growth planes, and its position with the majority of the diamond below the surface, it was not possible to detect any inclusions with a binocular microscopy. However, with the efficient use of a prototype, XRD instrument (see Methods), it was possible to perform a complete XRD scan of the internal part of the diamond, without extracting it from the host rock; this resulted in the detection and identification of two different inclusions, one an olivine and the other a peridotitic clinopyroxene. The experimental technique adopted in this work permits determination of the orientation matrixes of the two inclusions, as well as that of the diamond. In this manner, it was possible to determine reciprocal crystallographic orientations of the inclusions–diamond pairs24,31. The two inclusions (i.e., olivine and clinopyroxene) do not show any specific crystallographic orientation relative to each other, and importantly, they do not have any orientational relationships with the host diamond (see Table 1). Based upon prior detailed crystallographic studies, this result was expected for the diamond–olivine pair24,31. We are not familiar with any previous study of diamond–clinopyroxene orientation relationships. However, the presence of a clinopyroxene-bearing diamond embedded in a rock matrix containing a clinopyroxene presents yet another intriguing possibility.

This exceptional rock fragment, with a super-imposed gem-quality, millimetre-sized diamond, contains olivine and clinopyroxene inclusions identical in chemistry and crystallographic orientation to those outside the diamond, a clear evidence for protogenesis of the olivine inclusions, with a genesis similar that is illustrated in Fig. 2.
The presence of a single crystal of clinopyroxene positioned externally to the diamond is shown in Fig. 3. This ‘groundmass’ clinopyroxene is set within a matrix of serpentine alteration and is located at around 0.1 mm from the external surface of the diamond. As evident in Fig. 3, the clinopyroxene crystal is not optically distinguishable. However, XRD analysis definitively identified this external clinopyroxene. Unexpectedly, the crystallographic orientation of this groundmass clinopyroxene outside the diamond is identical to the clinopyroxene included within the diamond. Minor angular mismatches between the crystallographic axes are only 0.8°, 0.6° and 1.6° degrees, for a, b and c axes, respectively (see Table 1); these angular values are within one experimental uncertainty. This means that the two crystals have an identical crystallographic orientation—that is, clinopyroxene outside and inside the diamond are, in our interpretation, part of one crystal. The significance of this observation is paramount to the question of syngenetic versus protogenetic diamond mineral inclusions, at least for non-gem-quality, coated, poly-crystalline diamonds, for which different interpretations have been reported35,36,37,38.
The surprising first experimental confirmation of co-incidence of the crystallographic orientation of the pyroxene outside and inside a diamond was effectively predicted as ‘remaining islands’ of pre-existing mono-crystals24. The significance of this observation is monumental to the genetic relations between diamond and its mineral inclusions. These exciting results from our present study indicate that such inclusions are definitively protogenetic. This is ‘proof-positive’ for protogenesis for the diamond and its inclusion.
Discussion
Formation age of diamonds is determined by measurements of their mineral inclusions: K–Ar in pyroxene, only reliable for dating the kimberlite; Rb–Sr and/or Sm–Nd in garnet and pyroxene; and Re–Os and U–Th–Pb in sulfide phases. The application of the Sm–Nd and Rb–Sr isotopic schemes, due to limiting mineral masses necessary for instrumental sensitivity, requires putting together literally hundreds of inclusions from hundreds of diamonds, whereas the sulfide isotopic schemes permit ages on single diamonds. To date, the most commonly used and agreed upon mineral system is the Re–Os of sulfide inclusions. However, the major system that has received the most attention, until recently, is that of the Sm–Nd in garnet and pyroxene39,40,41.
The Sm–Nd system for age-dating is restricted for use if an isochron can be constructed from two or more minerals in isotopic equilibrium or one mineral and its corresponding whole rock. A major assumption is thereby made that the garnets and clinopyroxenes in the host rocks were in diffusional equilibrium and encapsulated above their ‘closure temperatures’ (TC≈600–800 °C) for these radiogenic isotopes. This isolation effectively ‘freezes-in’ the exchange of isotopic components, and starts their radiogenic clocks. However, for this system, the minerals in the host rock can continue to isotopically inter-diffuse (of course only when they are touching, many parageneses are non-touching), thereby re-equilibrating until they reach their TC—usually the date of kimberlite eruption42. The key limitations on the application of isotopic decay pairs are the availability and size of the inclusions, the abundance levels of the radionuclides and instrumental sensitivity.
The main assumption for radiogenic isotopic age determinations for diamonds, as well as rocks and minerals, is that they reach conditions where the minerals have stopped ‘isotopic communication’ (diffusion) with their host rock and each other. As mentioned above, this can occur when they reach their isotopic closure temperature (TC); or when they are otherwise isolated from further isotopic diffusion with each other and their surroundings—for example, encapsulated in a diamond. All minerals are encapsulated in diamonds above their TC. In this encapsulation case, the minerals that are used for their radiogenic isotopes must have been the same ones in contact before isolation—for example, clinopyroxene and garnet. Therein is one of the major assumptions in age-dating of diamonds. Were the minerals that are being used in the isotopic-partitioning studies actually in contact prior to and during their encapsulation by the diamonds? Or, did they become diffusionally isolated while still in the rock, before encapsulation? In the case of a single-phase isotopic system, the encapsulation starts the isotopic system at a time=zero. This is the case for sulfide inclusions for Re–Os age determinations2,3,4,5, where the encapsulated immiscible-sulfide phase1 cools to an assemblage of pentlandite, pyrrhotite and chalcopyrite, and is isolated from any outside inter-action.
Landmark publications39,43 used the assemblage of garnet and clinopyroxene occurring as separate inclusions in diamonds—that is, non-touching=no elemental isotopic exchanges. Because there was a minimum mass necessary for instrumental sensitivity for accurate measurement of the isotopes in the minerals, literally hundreds of each mineral inclusion, from hundreds of diamonds, were accumulated together for the elaborate chemistry necessary for the isotopes—typically Sm–Nd and Rb–Sr—of the garnet and the clinopyroxene aggregations. In later studies40, the garnets were further separated into individual groups, depending upon colour—four in this specific study. They then only used the clinopyroxene and one of the four garnet groups for determining the diamond age, picking the garnet aggregation with the highest 143Nd/144Nd contents, and using the other three garnet groups to calculate the precision. The inherent assumption made in these studies is that all the diamonds formed instantaneously from a distinct event, and throughout the entire kimberlite. But the huge array of compositions for the garnets and clinopyroxenes testifies to a large variation in chemistry and time for the encapsulation into the diamonds.
A modification of the principles presented by Navon44 on the formation of diamonds is shown in Fig. 4, based upon the criticisms of studies that were conducted of 100s of garnets and clinopyroxenes for diamond age determinations. This addresses the mis-conception of using piles of mineral inclusions for such conclusions. It is our contention that the assumption is false that these hundreds of garnet and clinopyroxene inclusion grains are all represented by one equilibrated pair of minerals, yet in an entire kimberlite. In reality, the diamond formation occurred over 10 to 100 s of million years, and such Sm–Nd two-mineral isochrons39 give but some indication of an ‘average diamond age’, with large values of variability, and the acceptance of assumptions mentioned above. Such data were the best available at that time. However, in our opinion, the authors oversimplified the complications intrinsic to such practice. Abundant studies have shown that multiple silicate inclusions in diamonds represent multi-generations of diamonds that have experienced different metasomatic alterations18,20.

Based upon the discussion by Navon44, and with modifications, the principles of this graph caution against the usage of multitudes of diamond inclusions for dating of ‘one diamond event’. It demonstrates that the nature of the two-mineral isochron is obviously a ‘grossly weighted-average age’ and should only to be addressed and used with caution.
In summary, for the last couple of decades, culminating in the present study, mounting evidences have accrued that most, if not all, mineral inclusions are not syngenetic, but are protogenetic—that is, formed before the diamonds, perhaps billions of years before18,19,20. Several authors18,20,45 have presented evidence that questions and is considered to negate the assumption that the inclusions in diamonds from one kimberlite are of the similar composition or forming at the same time in isotopic equilibrium (syngenetic); this negated assumption is evidenced even within one xenolith, or even within one-single diamond. This was based upon some of the first diamondiferous eclogite tomography, with up to 78 macrodiamonds in one 65 g xenolith; each diamond was mapped as to its position relative to the others, and each diamond then examined and polished to reveal 1–6 mineral inclusions in situ on one polished diamond surface. Electron microprobe and secondary ion mass spectrometry (SIMS) analyses of the clinopyroxene inclusions in one diamond, for example, showed distinct chemical differences, including positive Eu anomalies. In fact, the clinopyroxene and garnet compositions in many diamonds possess inter-granular heterogeneities—they were completely different, gain to grain. Indeed, some authors15 examined multiple inclusions in one diamond; the 35 individual garnets inclusions in one diamond had compositions that covered the wide array of all eclogitic garnets from xenoliths recovered from kimberlites in Yakutia—all in one diamond. The additional five clinopyroxenes from this single diamond also exhibited inter-granular heterogeneities. Recently21,46, based upon water in olivine, pyroxene and garnets as diamond inclusions, caution was again presented for assuming syngenesis of simultaneous formation of diamonds and their inclusions.
The present study may help negate the use of ‘syngenesis’ for mineral inclusions in diamonds. This term has been used for the formation of the inclusion and its host diamond at the same instant and from the same formational process. This is the incorrect word for use with respect to diamond mineral inclusions. The isotopic systems of immiscible sulfide phases can be started by the isolation of encapsulation, thereby resulting in the inclusions giving the time of encapsulation. This is synchronous. However, to assume that pairs of silicate minerals were touching or otherwise in isotopic equilibrium before encapsulation, and give synchronous ages, is incorrect. The age of the diamond and that of the inclusions may be synchronous, even if the inclusions existed before the encapsulation, but they are not syngenetic in origin.
Methods
Single-crystal XRD
The identification of the minerals investigated in this work and the determination of the orientation matrixes of diamond and its mineral inclusions, in addition to the mineral external to the diamond, were made possible with a prototype instrument at the Department of Geosciences at the University of Padova. The instrument is a single-crystal X-ray diffractometer Supernova (Rigaku-Oxford Diffraction) equipped with a brilliant X-ray micro-source (X-ray radiation wavelength=0.71073 Å; spot-size at the sample=0.12 mm) and with the Pilatus 200K detector (Dectris). Such an instrument permits measurement of small crystals down to 5 μm47. The orientation matrixes were then treated by OrientXplot software48, which allows one to calculate easily the reciprocal crystallographic orientations of the diamond-inclusions system.
Data availability
All data generated or analysed during this study are included in this published article and incorporated in Table 1.
Additional information
How to cite this article: Nestola, F. et al. Mineral inclusions in diamonds may be synchronous but not syngenetic. Nat. Commun. 8, 14168 doi: 10.1038/ncomms14168 (2017).
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
Taylor, L. A. & Liu, Y. Sulfide inclusions in diamonds: not monosulfide solid solution. Russ. Geol. Geophys. 50, 1201–1211 (2009).
Pearson, D. G. et al. Archean Re-Os age for Siberian eclogites and constraints on Archean tectonics. Nature 374, 711–713 (1995).
Pearson, D. G., Davies, R. M., Shirey, S. B., Carlson, R. W. & Griffin, W. L. in Proc. Seventh Int. Kimberlite Conf., Vol. 2, 664–666 (Red Roof Design, Capetown, 1998a).
Pearson, D. G., Shirey, S. B., Harris, J. W. & Carlson, R. W. Sulphide inclusions in diamonds from the Koffiefontein kimberlite, S. Africa: constraints on diamond ages and mantle Re-Os systematics. Earth Planet. Sci. Lett. 160, 311–326 (1998b).
Shirey, S. B. et al. in Reviews in Mineralogy and Geochemistry Vol. 75, eds Hazen R. M., Jones A. P., Baross J. A. 355–421Mineralogical Society of America (2013).
Walter, M. J. et al. Deep mantle cycling of oceanic crust: evidence from diamonds and their mineral inclusions. Science 334, 54–57 (2011).
Pearson, D. G. et al. A hydrous mantle transition zone indicated by ringwoodite included within diamond. Nature 507, 221–224 (2014).
Izraeli, E. S., Harris, J. W. & Navon, O. Raman barometry of diamond formation. Earth Planet. Sci. Lett. 173, 351–360 (1999).
Sobolev, N. V. et al. Fossilized high pressure from the Earth’s deep interior: the coesite-in-diamond barometer. Proc. Natl Acad. Sci. USA 97, 11875–11879 (2000).
Nestola, F. et al. First crystal-structure determination of olivine in diamond: composition and implications for provenance in the Earth’s mantle. Earth Planet. Sci. Lett. 305, 249–255 (2011).
Harris, J. W. The recognition of diamond inclusions, Pt. 1: syngenetic inclusions. Ind. Diamond Rev. 28, 402–410 (1968).
Sobolev, V. S., Sobolev, N. V. & Lavrent’ev, Y. G. Inclusions in diamonds from a diamond-bearing eclogite. Dokl. Akad. Nauk. SSR 207, 164–167 (1972).
Harris, J. W. & Gurney, J. J. in The Properties of Diamonded. Field J. E. 555–591Academic Press (1979).
Pearson, D. G. & Shirey, S. B. in Application of Radiogenic Isotopes to Ore Deposit Research and Exploration. eds Lambert D. D., Ruiz J. 143–171Society of Economic Geologists (1999).
Sobolev, N. V. et al. Extreme chemical diversity in the mantle during eclogite diamond formation: evidence from 35 garnet and 5 pyroxene inclusions in a single diamond. Int. Geol. Rev. 40, 567–578 (1998).
Spetsius, Z. V., Belousova, E. A., Griffin, W. L., O’Reilly, S. Y. & Pearson, N. J. Archean sulfide inclusions in Paleozoic zircon megacrysts from the Mir kimberlite, Yakutia: implications for the dating of diamonds. Earth Planet. Sci. Lett. 199, 111–126 (2002).
Spetsius, Z. V. & Taylor, L. A. In in Proc. Int. Workshop—Plumes and Problems of Deep Sources of Alkaline Magmatism, 5–19 (Khabarovsk, Siberia, 2003).
Taylor, L. A. et al. Diamonds and their mineral inclusions and what they tell us: a detailed ‘pull-apart’ of a diamondiferous eclogite. Int. Geol. Rev. 42, 959–983 (2000).
Taylor, L. A., Anand, M. & Promprated, P. in Eighth Int. Kimberlite Conf., Long Abstract Volume (Victoria, Canada, 22-27 June 2003).
Taylor, L. A. & Anand, M. Diamonds: time capsules from the Siberian Mantle. Chem. Erde 64, 1–74 (2004).
Taylor, L. A. et al. Low water contents in diamond mineral inclusions: proto-genetic origin in a dry cratonic lithosphere. Earth Planet. Sci. Lett. 433, 125–132 (2016).
Thomassot, E. et al. Metasomatic diamond growth: a multi-isotope study (13C, 15N, 33S, 34S) of sulphide inclusions and their host diamonds from Jwaneng (Botswana). Earth Planet. Sci. Lett. 282, 79–90 (2009).
Jacob, D.E., Piazolo, S., Schreiber, A. & Trimby, P. Redox-freezing and nucleation of diamond via magnetite formation in the Earth’s mantle. Nat. Commun. 7, 11891 (2016).
Nestola, F. et al. Olivine with diamond-imposed morphology included in diamonds. Syngenesis or protogenesis? Int. Geol. Rev. 56, 1658–1667 (2014).
Meyer, H. O. A. Genesis of diamond: a mantle saga. Am. Mineral. 70, 344–355 (1985).
Meyer, H. O. A. in Mantle Xenolithsed. Nixon P. H. 501–523Wiley (1987).
Mitchell, R. S. & Giardini, A. A. Oriented olivine inclusions in diamond. Am. Mineral. 38, 136–138 (1953).
Ickert, R.B., Stachel, T., Stern, R.A. & Harris, J.W. Diamond from recycled crustal carbon documented by coupled δ18O–δ13C measurements of diamonds and their inclusions. Earth Planet. Sci. Lett. 364, 85–97 (2013).
Cartigny, P., Palot, M., Thomassot, E. & Harris, J. W. Diamond formation: a stable isotope perspective. Annu. Rev. Earth Planet. Sci. 42, 699–732 (2014).
Bruno, M., Rubbo, M., Aquilano, D., Massaro, F. R. & Nestola, F. Diamond and its olivine inclusions: a strange relation revealed by ab initio simulations. Earth Planet. Sci. Lett. 435, 31–35 (2016).
Milani, S., Nestola, F., Angel, R. J., Nimis, P. & Harris, J. W. Crystallographic orientations of olivine inclusions in diamonds. Lithos 265, 312–316 (2016).
Sunagawa, I. in Materials Science of the Earth’s Interior (ed. Sunagawa, I.) 61–103 (Terra Scientific Publishing Company, 1984).
Sunagawa, I. in Materials Science of the Earth’s Interior (ed. Sunagawa, I.) 303–331 (Terra Scientific Publishing Company, 1984).
Lee, J. & Jung, H. Lattice-preferred orientation of olivine found in diamond-bearing garnet peridotites in Finsch, South Africa and implications for seismic anisotropy. J. Struct. Geol. 70, 12–22 (2015).
Weiss, Y., McNeill, J., Pearson, D. G., Nowell, G. M. & Ottley, C. J. Highly saline fluids from a subducting slab as the source for fluid-rich diamonds. Nature 524, 339–342 (2015).
Mikhail, S., Dobosi, G., Verchovsky, A. B., Kurat, G. & Jones, A. P. Peridotitic and websteritic diamondites provide new information regarding mantle melting and metasomatism induced through the subduction of crustal volatiles. Geochim. Cosmochim. Acta 107, 1–11 (2013).
Jacob, D. E., Dobrzhinetskaya, L. & Wirth, R. New insight into polycrystalline diamond genesis from modern nanoanalytical techniques. Earth Sci. Rev. 136, 21–35 (2014).
Stachel, T. & Harris, J. W. The origin of cratonic diamonds—constraints from mineral inclusions. Ore Geol. Rev. 34, 5–32 (2008).
Richardson, S. H., Gurney, J. J., Erlank, A. J. & Harris, J. W. Origin of diamond from old continental mantle. Nature 310, 198–202 (1984).
Richardson, S. H., Erlank, A. J., Harris, J. W. & Hart, S. R. Eclogitic diamonds of Proterozoic age from Cretaceous kimberlites. Nature 346, 54–56 (1990).
Shirey, S. B. & Richardson, S. H. Start of the Wilson Cycle at 3 Ga shown by diamonds from subcontinental mantle. Science 333, 434–436 (2011).
Snyder, G. A. et al. Archean mantle heterogeneity and the origin of diamondiferous eclogites, Siberia—evidence from stable isotopes and hydroxyl in garnet. Am. Mineral. 80, 799–809 (1995).
Richardson, S. H. Latter-day origin of diamonds of eclogitic paragenesis. Nature 322, 623–626 (1986).
Navon, O. in Proceedings of Seventh International Kimberlite Conference, Vol. 2 (eds Gurney, J. J., Gurney, J. L., Pascoe, M. D. & Richardson, S. H.) 584–605 (Red Roof Design, 1999).
Taylor, L. A. et al. Petrogenesis of Group A eclogites: observations from the Obnazhennaya kimberlite, Yakutia. Contrib. Miner. Petrol. 145, 424–443 (2003).
Novella, D., Bolfan-Casanova, N., Nestola, F. & Harris, J. W. H2O in olivine and garnet inclusions still trapped in diamonds from the Siberian craton: Implications for the water content of cratonic lithosphere peridotites. Lithos 230, 180–183 (2015).
Angel, R. J. & Nestola, F. A century of mineral structures: how well do we know them? Am. Mineral. 101, 1036–1045 (2016).
Angel, R. J., Milani, S., Alvaro, M. & Nestola, F. OrientXplot: a program to analyse and display relative crystal orientations. J. Appl. Crystallogr 48, 1330–1334 (2015).
Acknowledgements
The research was supported by the ERC Starting Grant INDIMEDEA to F.N. (No. 307322), the NRF grant to H.J. (NRF-2015R1A2A1A15052305) and an NSF grant to L.A.T. (EAR-1523303). We are especially grateful to Steve Shirey, Graham Pearson and Pierre Cartigny for useful and invigorating scientific discussions, and the two referees, S. Mikhail and a second anonymous one, for very constructive comments that led to significant improvement in this paper.
Author information
Authors and Affiliations
Contributions
F.N. wrote the initial manuscript and performed the X-ray diffraction experiments; H.J. wrote part of the manuscript and provided the sample; L.A.T. re-wrote the entire manuscript and contributed to overall discussion of synchronous diamond inclusions.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Nestola, F., Jung, H. & Taylor, L. Mineral inclusions in diamonds may be synchronous but not syngenetic. Nat Commun 8, 14168 (2017). https://doi.org/10.1038/ncomms14168
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/ncomms14168
This article is cited by
-
A critique of using epitaxial criterion to discriminate between protogenetic and syngenetic mineral inclusions in diamond
Scientific Reports (2024)
-
Imperfections in natural diamond: the key to understanding diamond genesis and the mantle
La Rivista del Nuovo Cimento (2023)
-
Has Axial Spin Decline Affected Earth’s Geologic and Tectonic History?
Journal of Earth Science (2022)
-
Helium in diamonds unravels over a billion years of craton metasomatism
Nature Communications (2021)
-
The role of elastic anisotropy in determining the depth of formation for diamonds and their inclusions
Rendiconti Lincei. Scienze Fisiche e Naturali (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
