
Abstract
L-Glutamine (Gln) functions physiologically to balance the carbon and nitrogen requirements of tissues. It has been proposed that in cancer cells undergoing aerobic glycolysis, accelerated anabolism is sustained by Gln-derived carbons, which replenish the tricarboxylic acid (TCA) cycle (anaplerosis). However, it is shown here that in glioblastoma (GBM) cells, almost half of the Gln-derived glutamate (Glu) is secreted and does not enter the TCA cycle, and that inhibiting glutaminolysis does not affect cell proliferation. Moreover, Gln-starved cells are not rescued by TCA cycle replenishment. Instead, the conversion of Glu to Gln by glutamine synthetase (GS; cataplerosis) confers Gln prototrophy, and fuels de novo purine biosynthesis. In both orthotopic GBM models and in patients, 13C–glucose tracing showed that GS produces Gln from TCA-cycle-derived carbons. Finally, the Gln required for the growth of GBM tumours is contributed only marginally by the circulation, and is mainly either autonomously synthesized by GS-positive glioma cells, or supplied by astrocytes.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout








Similar content being viewed by others

References
Moreadith, R. W. & Lehninger, A. L. The pathways of glutamate and glutamine oxidation by tumor cell mitochondria. Role of mitochondrial NAD(P) + -dependent malic enzyme. J. Biol. Chem. 259, 6215–6221 (1984).
Yuneva, M. O. et al. The metabolic profile of tumors depends on both the responsible genetic lesion and tissue type. Cell Metab. 15, 157–170 (2012).
Wise, D. R. et al. Myc regulates a transcriptional program that stimulates mitochondrial glutaminolysis and leads to glutamine addiction. Proc. Natl Acad. Sci. USA 105, 18782–18787 (2008).
DeBerardinis, R. J. et al. Beyond aerobic glycolysis: transformed cells can engage in glutamine metabolism that exceeds the requirement for protein and nucleotide synthesis. Proc. Natl Acad. Sci. USA 104, 19345–19350 (2007).
Tardito, S. et al. L-Asparaginase and inhibitors of glutamine synthetase disclose glutamine addiction of β-catenin-mutated human hepatocellular carcinoma cells. Curr. Cancer Drug Targets 11, 929–943 (2011).
Son, J. et al. Glutamine supports pancreatic cancer growth through a KRAS-regulated metabolic pathway. Nature 496, 101–105 (2013).
Gameiro, P. A. et al. In vivo HIF-mediated reductive carboxylation is regulated by citrate levels and sensitizes VHL-deficient cells to glutamine deprivation. Cell Metab. 17, 372–385 (2013).
Willems, L. et al. Inhibiting glutamine uptake represents an attractive new strategy for treating acute myeloid leukemia. Blood 122, 3521–3532 (2013).
Chiu, M. et al. Glutamine depletion by crisantaspase hinders the growth of human hepatocellular carcinoma xenografts. Br. J. Cancer 111, 1159–1167 (2014).
Wang, J. B. et al. Targeting mitochondrial glutaminase activity inhibits oncogenic transformation. Cancer Cell 18, 207–219 (2010).
Suzuki, S. et al. Phosphate-activated glutaminase (GLS2), a p53-inducible regulator of glutamine metabolism and reactive oxygen species. Proc. Natl Acad. Sci. USA 107, 7461–7466 (2010).
Dal Bello, B. et al. Glutamine synthetase immunostaining correlates with pathologic features of hepatocellular carcinoma and better survival after radiofrequency thermal ablation. Clin. Cancer Res. 16, 2157–2166 (2010).
Krebs, H. A. Metabolism of amino-acids: the synthesis of glutamine from glutamic acid and ammonia, and the enzymic hydrolysis of glutamine in animal tissues. Biochem. J. 29, 1951–1969 (1935).
van den Berg, C. J. & Garfinkel, D. A stimulation study of brain compartments. Metabolism of glutamate and related substances in mouse brain. Biochem. J. 123, 211–218 (1971).
Takano, T. et al. Glutamate release promotes growth of malignant gliomas. Nat. Med. 7, 1010–1015 (2001).
Scott, D. A. et al. Comparative metabolic flux profiling of melanoma cell lines: beyond the Warburg effect. J. Biol. Chem. 286, 42626–42634 (2011).
Metallo, C. M. et al. Reductive glutamine metabolism by IDH1 mediates lipogenesis under hypoxia. Nature 481, 380–384 (2012).
Kamphorst, J. J. et al. Hypoxic and Ras-transformed cells support growth by scavenging unsaturated fatty acids from lysophospholipids. Proc. Natl Acad. Sci. USA 110, 8882–8887 (2013).
Fan, J., Kamphorst, J. J., Rabinowitz, J. D. & Shlomi, T. Fatty acid labeling from glutamine in hypoxia can be explained by isotope exchange without net reductive isocitrate dehydrogenase (IDH) flux. J. Biol. Chem. 288, 31363–31369 (2013).
Mullen, A. R. et al. Reductive carboxylation supports growth in tumour cells with defective mitochondria. Nature 481, 385–388 (2012).
Lewerenz, J. et al. The cystine/glutamate antiporter system x(c)(-) in health and disease: from molecular mechanisms to novel therapeutic opportunities. Antioxid. Redox Signal. 18, 522–555 (2013).
Duarte, N. C. et al. Global reconstruction of the human metabolic network based on genomic and bibliomic data. Proc. Natl Acad. Sci. USA 104, 1777–1782 (2007).
Gao, P. et al. c-Myc suppression of miR-23a/b enhances mitochondrial glutaminase expression and glutamine metabolism. Nature 458, 762–765 (2009).
Yuneva, M., Zamboni, N., Oefner, P., Sachidanandam, R. & Lazebnik, Y. Deficiency in glutamine but not glucose induces MYC-dependent apoptosis in human cells. J. Cell Biol. 178, 93–105 (2007).
Bax, D. A. et al. Molecular and phenotypic characterisation of paediatric glioma cell lines as models for preclinical drug development. PLoS ONE 4, e5209 (2009).
Filonov, G. S. et al. Bright and stable near-infrared fluorescent protein for in vivo imaging. Nat. Biotechnol. 29, 757–761 (2011).
Hock, A. K. et al. iRFP is a sensitive marker for cell number and tumor growth in high-throughput systems. Cell Cycle 13, 220–226 (2013).
Yin, Y. et al. Glutamine synthetase functions as a negative growth regulator in glioma. J. Neurooncol. 114, 59–69 (2013).
Reznik, E., Mehta, P. & Segre, D. Flux imbalance analysis and the sensitivity of cellular growth to changes in metabolite pools. PLoS Comput. Biol. 9, e1003195 (2013).
Calise, S. J. et al. Glutamine deprivation initiates reversible assembly of mammalian rods and rings. Cell. Mol. Life Sci. 71, 2963–2973 (2014).
Carruthers, R. et al. Abrogation of radioresistance in glioblastoma stem-like cells by inhibition of ATM kinase. Mol. Oncol. 9, 192–203 (2015).
Meyerand, M. E., Pipas, J. M., Mamourian, A., Tosteson, T. D. & Dunn, J. F. Classification of biopsy-confirmed brain tumors using single-voxel MR spectroscopy. AJNR Am. J. Neuroradiol. 20, 117–123 (1999).
Parmentier, J. H. et al. Glutaminase activity determines cytotoxicity of L-asparaginases on most leukemia cell lines. Leuk. Res. 39, 757–762 (2015).
Had-Aissouni, L. Toward a new role for plasma membrane sodium-dependent glutamate transporters of astrocytes: maintenance of antioxidant defenses beyond extracellular glutamate clearance. Amino Acids 42, 181–197 (2012).
Rosati, A. et al. Glutamine synthetase expression as a valuable marker of epilepsy and longer survival in newly diagnosed glioblastoma multiforme. Neuro-oncol. 15, 618–625 (2013).
Marin-Valencia, I. et al. Analysis of tumor metabolism reveals mitochondrial glucose oxidation in genetically diverse human glioblastomas in the mouse brain in vivo. Cell Metab. 15, 827–837 (2012).
Maher, E. A. et al. Metabolism of [U-13 C]glucose in human brain tumors in vivo. NMR Biomed. 25, 1234–1244 (2012).
Bagga, P. et al. Characterization of cerebral glutamine uptake from blood in the mouse brain: implications for metabolic modeling of 13C NMR data. J. Cereb. Blood Flow Metab. 34, 1666–1672 (2014).
Rosati, A. et al. Epilepsy in glioblastoma multiforme: correlation with glutamine synthetase levels. J. Neurooncol. 93, 319–324 (2009).
Noble, M. & Murray, K. Purified astrocytes promote the in vitro division of a bipotential glial progenitor cell. EMBO J. 3, 2243–2247 (1984).
Keunen, O. et al. Anti-VEGF treatment reduces blood supply and increases tumor cell invasion in glioblastoma. Proc. Natl Acad. Sci. USA 108, 3749–3754 (2011).
Sanzey, M. et al. Comprehensive analysis of glycolytic enzymes as therapeutic targets in the treatment of glioblastoma. PLoS ONE 10, e0123544 (2015).
Acknowledgements
This study has been supported by Cancer Research UK. S.T. is a recipient of an AIRC/Marie Curie International Fellowship for Cancer Research. The human and animal metabolomic studies were supported by The Norwegian Cancer Society, The Norwegian Research Council, Helse Vest, Haukeland University Hospital and the K.G-Jebsen Foundation. We acknowledge A. Golebiewska, V. Baus-Talko, N. Van Den Broek, G. MacKay, C. Nixon and E. MacKenzie for excellent technical assistance and A. King for excellent editorial work.
Author information
Authors and Affiliations
Contributions
S.T. conceived the study, designed and performed most experiments, interpreted the data, and wrote the manuscript, A.O. performed the experiments in orthotopic xenograft models, S.U.A. and A.J.C. provided the differentiated and stem-like primary glioblastoma cells, L.Z. supervised the analysis of LC-MS samples, O.K. performed the MRI analysis, F.F. processed the orthotopic and clinical GBM samples, H.M. provided the tissue microarray, A.K.H. designed and provided the iRFP and iRFP–GS constructs, A.Weinstock, A.Wagner and E.R. generated and employed the metabolic modelling, S.C.B. and S.L.L. provided the primary astrocytes, M.L.-J., S.H.M. and P.Ø.S. provided the surgical specimens from the patients, S.P.N. and R.B. conceived and supervised the experiments in orthotopic models and human patients, and E.G. conceived and supervised the study, interpreted the data, and revised the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Integrated supplementary information
Supplementary Figure 5 Glutamine starvation reduces GBM cell proliferation.
(a) Representative microscopic fields of cells incubated for 6 days with or without Gln. (b) Cell cycle distribution of cells incubated for 3 days with or without Gln. Mean ± S.E.M. n = 3 independent experiments. p value refers to a two-tailed t test for unpaired samples. Raw data of independent repeats are provided in the statistics source data Supplementary Table 5.
Supplementary Figure 6 Exchange rates of metabolites in GBM cell lines.
Cells were incubated for 24 h +/− U-13C5 Gln, secretion rates (positive bars) and consumption rates (negative bars) of the indicated metabolites are shown. The sum of all isotopologues is reported for clarity. Alanine (Ala) exchange rates are magnified in the inset. Mean ± S.E.M. n = 3 independent experiments.
Supplementary Figure 7 Effects of Gln availability and Glutaminase inhibition on GBM cell lines growth and redox state.
(a) Correlation between Gln consumption and growth inhibition caused by Gln starvation is shown in six cell lines. Mean ± S.E.M. n = 3 independent experiments. (b) Citrate isotopologues distribution of 13C5-Gln derived atoms. 13C6-Citrate was not detected. Mean ± S.E.M. n = 3 independent experiments. (c) Oxidized (GSSG) to reduced (GSH) glutathione ratio. LN18 cells were incubated for 24 h +/− Gln and supplemented with Glu, α-ketoglutarate dimethylester (dm-αKG), sulfasalazine (SSZ), or cystine, as indicated. GSSG/GSH ratio was assessed by HPLC-MS, and shown as % of untreated control. Data derive from one experiment performed once. Raw data of independent repeats are provided in the statistics source data Supplementary Table 5. (d–i) Cells were incubated with 0, 2.5, 5, 10, 15, 30 μM BPTES for 72 h, in the absence (solid line) or presence (dotted line) of 4 mM dm-αKG and counted. In all conditions cells were exposed to 0.3%DMSO. Mean ± S.E.M. n = 3 independent experiments.
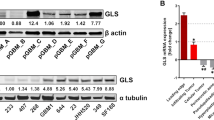
Supplementary Figure 8 GS expression and its effects on colony formation capacity, and glucosamine biosynthetic pathway in GBM cells.
(a) Cells were incubated for 24h +/− Gln and GLUL (GS), GLS, and MYC mRNA relative expression was assessed by qPCR. Actin expression was used for normalization. Data derive from one experiment performed once. Raw data of independent repeats are provided in the statistics source data Supplementary Table 5. (b) SF188 and U251 cells stably expressing a non-targeting control shRNA (shNTC) and two sequences targeting GS (shGS-1 and shGS-2) were cultured for 12–17 days +/− Gln in medium supplemented with glutamate (4 mM), and ammonia (0.8 mM) as indicated. The quantification of colonies surface area was obtained as described in the Methods section. Mean ± S.E.M., n = 4 independent experiments. (c) iRFP4 and iRFP-GS5 expressing LN18 cells were incubated +/− Gln for 24 h in medium supplemented with 0.8 mM 15NH4+. The intracellular isotopologues distribution of UDP N-acetylglucosamine is shown as % of control (value obtained for the metabolites in iRFP4 cells + Gln). Data derive from one experiment performed twice. Raw data of independent repeats are provided in the statistics source data Supplementary Table 5.
Supplementary Figure 9 Heterogeneous GS expression in human GBM biopsies.
The whole bioptic tissues of three patients included in the tissue microarray were stained for GS, and Hematoxilin and Eosin [H&E], as indicated. Low magnifications of whole biopsies are reported in panels (a,b,h,i,n,o). For each biopsy, tissue regions framed in blue and green are magnified in d–g,j–m,q–t. Biopsy 1 shows necrotic tissue [nec] surrounded by a solid tumour core [TC] with uniformly low GS staining (d,f). (d,e) Invasive tumour areas [TI] show infiltrative cells with heterogeneous immunoreactivity for GS. (c) Normal astrocytes [black arrows] in the adjacent normal brain [Br] were positive for GS, while neurons [white arrows] were GS-negative. Biopsy 2 shows distinct areas with low/absent (k) and high (j) immunoreactivity for GS. GS-negative inflammatory tissue [i] surrounding a necrotic region [nec] is evident in panels h,i,k–m. Biopsy 3 shows distinct areas with low/absent (q) and high (r) immunoreactivity for GS. (p) GS-positive reactive astrocytes [arrows] with extensive cellular processes forming a network of GS positive fibers surrounding GS-negative tumour cells.
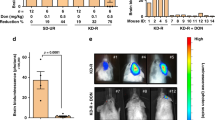
Supplementary Figure 10 Glucose and glutamine metabolic fates in GBM patients, and orthotopic mouse models.
(a) Serum levels of 13C6 and 13C0 glucose in three GBM patients injected with 13C6 glucose before surgical intervention for tumour removal. The dashed line represents the time at which glucose infusion starts. Black arrow points at the time of surgical resection. Values are reported as % of the basal values obtained before infusion. (b) 13C Lactate and (c) 13C Glutamate enrichment in serum at time of tumour resection, in tumour tissue, and in adjacent edematous tissue, of seven (1-7) GBM patients injected with 13C6-glucose. The % of lactate or glutamate incorporating one or more 13C carbons over the total amount of lactate or glutamate detected is reported; na: not available, nd: not detectable. Values were corrected for the natural abundance of 13C. (d,e) Isotopologue distribution of metabolites obtained in the liver of mice orthotopically xenografted with human P3 GBM and injected with 13C6 Glucose (d) or 13C5-Gln (e). The values are mean ± S.E.M. n = 3 mice. (f,g) Isotopologues of Gln (f) and glucose (g) found in the serum of mice orthotopically xenografted with the human GS-positive T16 GBM, and infused for 4h into the carotid artery with 13C5-Gln. Mean ± S.E.M. n = 3 mice. (h–j) Isotopologue distributions of Gln, Glu and α-kg in T16 tumours, contralateral brain, and liver tissues of mice infused as in (f,g). Mean ± S.E.M. n = 3 mice. (k) Astrocytes were incubated for 24 hours +/− Gln, and asparagine consumption is reported. Data derive from one experiment performed twice. Raw data of independent repeats are provided in the statistics source data Supplementary Table 5.
Supplementary Figure 11 A toy network used for metabolic modelling.
A toy network (a) and its optimal flux distribution (b).
Supplementary Figure 12 Unprocessed scans of western blots accompanied by size markers.
Red dotted lines delineate the region presented in the respective Figures as indicated. Images were obtained using a Licor Odyssey scanner and acquired using Image Studio 2.0.
Supplementary information
Supplementary Information
Supplementary Information (PDF 2806 kb)
Supplementary Table 5
Supplementary Information (XLSX 130 kb)
Rights and permissions
About this article

Cite this article
Tardito, S., Oudin, A., Ahmed, S. et al. Glutamine synthetase activity fuels nucleotide biosynthesis and supports growth of glutamine-restricted glioblastoma. Nat Cell Biol 17, 1556–1568 (2015). https://doi.org/10.1038/ncb3272
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ncb3272
This article is cited by
-
Metabolic pathway analysis using stable isotopes in patients with cancer
Nature Reviews Cancer (2023)
-
BCL2L13 promotes mitophagy through DNM1L-mediated mitochondrial fission in glioblastoma
Cell Death & Disease (2023)
-
A collaborative synthetase
Nature Chemical Biology (2023)
-
STAT proteins in cancer: orchestration of metabolism
Nature Reviews Cancer (2023)
-
Long-term 1,2-dimethylhydrazine triggers pathological remodeling of colon mucosa through repression of sestrin2, nuclear factor (erythroid-derived 2)-like 2, and sirtuin4 stimulating mitochondrial stress and metabolic reprogramming
Naunyn-Schmiedeberg's Archives of Pharmacology (2023)
