
Abstract
The heat-shock response is a complex cellular program that induces major changes in protein translation, folding and degradation to alleviate toxicity caused by protein misfolding. Although heat shock has been widely used to study proteostasis, it remained unclear how misfolded proteins are targeted for proteolysis in these conditions. We found that Rsp5 and its mammalian homologue Nedd4 are important E3 ligases responsible for the increased ubiquitylation induced by heat stress. We determined that Rsp5 ubiquitylates mainly cytosolic misfolded proteins upon heat shock for proteasome degradation. We found that ubiquitylation of heat-induced substrates requires the Hsp40 co-chaperone Ydj1 that is further associated with Rsp5 upon heat shock. In addition, ubiquitylation is also promoted by PY Rsp5-binding motifs found primarily in the structured regions of stress-induced substrates, which can act as heat-induced degrons. Our results support a bipartite recognition mechanism combining direct and chaperone-dependent ubiquitylation of misfolded cytosolic proteins by Rsp5.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout








Similar content being viewed by others


References
Kim, Y. E., Hipp, M. S., Bracher, A., Hayer-Hartl, M. & Hartl, F. U. Molecular chaperone functions in protein folding and proteostasis. Annu. Rev. Biochem. 82, 323–355 (2013).
Powers, E. T. & Balch, W. E. Diversity in the origins of proteostasis networks—a driver for protein function in evolution. Nat. Rev. Mol. Cell Biol. 14, 237–248 (2013).
Chen, B., Retzlaff, M., Roos, T. & Frydman, J. Cellular strategies of protein quality control. Cold Spring Harb. Perspect. Biol. 3, a004374 (2011).
Comyn, S. A., Chan, G. T. & Mayor, T. False start: cotranslational protein ubiquitination and cytosolic protein quality control. J. Proteomics 100, 92–101 (2014).
Brodsky, J. L. Cleaning up: ER-associated degradation to the rescue. Cell 151, 1163–1167 (2012).
Vabulas, R. M., Raychaudhuri, S., Hayer-Hartl, M. & Hartl, F. U. Protein folding in the cytoplasm and the heat shock response. Cold Spring Harb. Perspect. Biol. 2, a004390 (2010).
Morimoto, R. I. The heat shock response: systems biology of proteotoxic stress in aging and disease. Cold Spring Harb. Symp. Quant. Biol. 76, 91–99 (2011).
Carlson, N., Rogers, S. & Rechsteiner, M. Microinjection of ubiquitin: changes in protein degradation in HeLa cells subjected to heat-shock. J. Cell Biol. 104, 547–555 (1987).
Parag, H. A., Raboy, B. & Kulka, R. G. Effect of heat shock on protein degradation in mammalian cells: involvement of the ubiquitin system. EMBO J. 6, 55–61 (1987).
Medicherla, B. & Goldberg, A. L. Heat shock and oxygen radicals stimulate ubiquitin-dependent degradation mainly of newly synthesized proteins. J. Cell Biol. 182, 663–673 (2008).
Fang, N. N., Ng, A. H., Measday, V. & Mayor, T. Hul5 HECT ubiquitin ligase plays a major role in the ubiquitylation and turnover of cytosolic misfolded proteins. Nat. Cell Biol. 13, 1344–1352 (2011).
Galan, J. M., Moreau, V., Andre, B., Volland, C. & Haguenauer-Tsapis, R. Ubiquitination mediated by the Npi1p/Rsp5p ubiquitin-protein ligase is required for endocytosis of the yeast uracil permease. J. Biol. Chem. 271, 10946–10952 (1996).
Huibregtse, J. M., Yang, J. C. & Beaudenon, S. L. The large subunit of RNA polymerase II is a substrate of the Rsp5 ubiquitin-protein ligase. Proc. Natl Acad. Sci. USA 94, 3656–3661 (1997).
Hoppe, T. et al. Activation of a membrane-bound transcription factor by regulated ubiquitin/proteasome-dependent processing. Cell 102, 577–586 (2000).
Zhao, Y., Macgurn, J. A., Liu, M. & Emr, S. The ART-Rsp5 ubiquitin ligase network comprises a plasma membrane quality control system that protects yeast cells from proteotoxic stress. Elife 2, e00459 (2013).
Keener, J. M. & Babst, M. Quality control and substrate-dependent downregulation of the nutrient transporter Fur4. Traffic 14, 412–427 (2013).
Crapeau, M., Merhi, A. & Andre, B. Stress conditions promote yeast Gap1 permease ubiquitylation and downregulation via the arrestin-like bul and aly proteins. J. Biol. Chem. 289, 22103–22116 (2014).
Shahsavarani, H., Sugiyama, M., Kaneko, Y., Chuenchit, B. & Harashima, S. Superior thermotolerance of Saccharomyces cerevisiae for efficient bioethanol fermentation can be achieved by overexpression of RSP5 ubiquitin ligase. Biotechnol. Adv. 30, 1289–1300 (2012).
Wang, G., Yang, J. & Huibregtse, J. M. Functional domains of the Rsp5 ubiquitin-protein ligase. Mol. Cell. Biol. 19, 342–352 (1999).
Krsmanovic, T. & Kolling, R. The HECT E3 ubiquitin ligase Rsp5 is important for ubiquitin homeostasis in yeast. FEBS Lett. 577, 215–219 (2004).
Haitani, Y. & Takagi, H. Rsp5 is required for the nuclear export of mRNA of HSF1 and MSN2/4 under stress conditions in Saccharomyces cerevisiae. Genes Cells 13, 105–116 (2008).
Rodriguez, M. S., Gwizdek, C., Haguenauer-Tsapis, R. & Dargemont, C. The HECT ubiquitin ligase Rsp5p is required for proper nuclear export of mRNA in Saccharomyces cerevisiae. Traffic 4, 566–575 (2003).
Zarrinpar, A. & Lim, W. A. Converging on proline: the mechanism of WW domain peptide recognition. Nat. Struct. Biol. 7, 611–613 (2000).
Wagner, S. A. et al. A proteome-wide, quantitative survey of ubiquitylation sites reveals widespread regulatory roles. in vivo 10, M111.013284 (2011).
Kim, W. et al. Systematic and quantitative assessment of the ubiquitin-modified proteome. Mol. Cell 44, 325–340 (2011).
Dunn, R., Klos, D. A., Adler, A. S. & Hicke, L. The C2 domain of the Rsp5 ubiquitin ligase binds membrane phosphoinositides and directs ubiquitination of endosomal cargo. J. Cell Biol. 165, 135–144 (2004).
Crosas, B. et al. Ubiquitin chains are remodeled at the proteasome by opposing ubiquitin ligase and deubiquitinating activities. Cell 127, 1401–1413 (2006).
Aviram, S. & Kornitzer, D. The ubiquitin ligase Hul5 promotes proteasomal processivity. Mol. Cell. Biol. 30, 985–994 (2010).
Kampinga, H. H. & Craig, E. A. The HSP70 chaperone machinery: J proteins as drivers of functional specificity. Nat. Rev. Mol. Cell Biol. 11, 579–592 (2010).
Lee, D. H., Sherman, M. Y. & Goldberg, A. L. Involvement of the molecular chaperone Ydj1 in the ubiquitin-dependent degradation of short-lived and abnormal proteins in Saccharomyces cerevisiae. Mol. Cell. Biol. 16, 4773–4781 (1996).
Metzger, M. B., Maurer, M. J., Dancy, B. M. & Michaelis, S. Degradation of a cytosolic protein requires endoplasmic reticulum-associated degradation machinery. J. Biol. Chem. 283, 32302–32316 (2008).
Gupta, R. et al. Ubiquitination screen using protein microarrays for comprehensive identification of Rsp5 substrates in yeast. Mol. Syst. Biol. 3, 116 (2007).
Ghaemmaghami, S. et al. Global analysis of protein expression in yeast. Nature 425, 737–741 (2003).
Kim, H. C. & Huibregtse, J. M. Polyubiquitination by HECT E3s and the determinants of chain type specificity. Mol. Cell. Biol. 29, 3307–3318 (2009).
Park, S. H. et al. PolyQ proteins interfere with nuclear degradation of cytosolic proteins by sequestering the Sis1p chaperone. Cell 154, 134–145 (2013).
Summers, D. W., Wolfe, K. J., Ren, H. Y. & Cyr, D. M. The Type II Hsp40 Sis1 cooperates with Hsp70 and the E3 ligase Ubr1 to promote degradation of terminally misfolded cytosolic protein. PLoS ONE 8, e52099 (2013).
Tofaris, G. K. et al. Ubiquitin ligase Nedd4 promotes α-synuclein degradation by the endosomal-lysosomal pathway. Proc. Natl Acad. Sci. USA 108, 17004–17009 (2011).
Tardiff, D. F. et al. Yeast reveal a “Druggable” Rsp5/Nedd4 network that ameliorates α-synuclein toxicity in neurons. Science 342, 979–983 (2013).
Lu, K., Psakhye, I. & Jentsch, S. Autophagic clearance of polyQ proteins mediated by ubiquitin-Atg8 adaptors of the conserved CUET protein family. Cell 158, 549–563 (2014).
Li, Z. et al. Systematic exploration of essential yeast gene function with temperature-sensitive mutants. Nat. Biotechnol. 29, 361–367 (2011).
Mnaimneh, S. et al. Exploration of essential gene functions via titratable promoter alleles. Cell 118, 31–44 (2004).
Nikko, E. & Pelham, H. R. Arrestin-mediated endocytosis of yeast plasma membrane transporters. Traffic 10, 1856–1867 (2009).
Longtine, M. S. et al. Additional modules for versatile and economical PCR-based gene deletion and modification in Saccharomyces cerevisiae. Yeast 14, 953–961 (1998).
Saeki, Y., Isono, E. & Toh-E, A. Preparation of ubiquitinated substrates by the PY motif-insertion method for monitoring 26S proteasome activity. Methods Enzymol. 399, 215–227 (2005).
Gajewska, B. et al. WW domains of Rsp5p define different functions: determination of roles in fluid phase and uracil permease endocytosis in Saccharomyces cerevisiae. Genetics 157, 91–101 (2001).
Persaud, A. et al. Comparison of substrate specificity of the ubiquitin ligases Nedd4 and Nedd4-2 using proteome arrays. Mol. Syst. Biol. 5, 333 (2009).
Terrell, J., Shih, S., Dunn, R. & Hicke, L. A function for monoubiquitination in the internalization of a G protein-coupled receptor. Mol. Cell 1, 193–202 (1998).
Miyazaki, K. MEGAWHOP cloning: a method of creating random mutagenesis libraries via megaprimer PCR of whole plasmids. Methods Enzymol. 498, 399–406 (2011).
Persaud, A. et al. Tyrosine phosphorylation of NEDD4 activates its ubiquitin ligase activity. Sci. Signal. (in the press; 2014)
Fouladkou, F. et al. The ubiquitin ligase Nedd4-1 is dispensable for the regulation of PTEN stability and localization. Proc. Natl Acad. Sci. USA 105, 8585–8590 (2008).
Miller, M. J., Xuong, N. H. & Geiduschek, E. P. A response of protein synthesis to temperature shift in the yeast Saccharomyces cerevisiae. Proc. Natl Acad. Sci. USA 76, 5222–5225 (1979).
Ng, A. H., Fang, N. N., Comyn, S. A., Gsponer, J. & Mayor, T. System-wide analysis reveals intrinsically disordered proteins are prone to ubiquitylation after misfolding stress. Mol. Cell. Proteomics 12, 2456–2467 (2013).
Udeshi, N. D. et al. Refined preparation and use of anti-diglycine remnant (K-epsilon-GG) antibody enables routine quantification of 10,000s of ubiquitination sites in single proteomics experiments. Mol. Cell. Proteomics 12, 825–831 (2013).
Rappsilber, J., Mann, M. & Ishihama, Y. Protocol for micro-purification, enrichment, pre-fractionation and storage of peptides for proteomics using StageTips. Nat. Protoc. 2, 1896–1906 (2007).
Kristensen, A. R., Gsponer, J. & Foster, L. J. A high-throughput approach for measuring temporal changes in the interactome. Nat. Methods 9, 907–909 (2012).
Olsen, J. V. et al. Parts per million mass accuracy on an Orbitrap mass spectrometer via lock mass injection into a C-trap. Mol. Cell. Proteomics 4, 2010–2021 (2005).
Mayor, T., Graumann, J., Bryan, J., MacCoss, M. J. & Deshaies, R. J. Quantitative profiling of ubiquitylated proteins reveals proteasome substrates and the substrate repertoire influenced by the Rpn10 receptor pathway. Mol. Cell. Proteomics 6, 1885–1895 (2007).
Huh, W. K. et al. Global analysis of protein localization in budding yeast. Nature 425, 686–691 (2003).
Ward, J. J., Sodhi, J. S., McGuffin, L. J., Buxton, B. F. & Jones, D. T. Prediction and functional analysis of native disorder in proteins from the three kingdoms of life. J. Mol. Biol. 337, 635–645 (2004).
Bryson, K. et al. Protein structure prediction servers at University College London. Nucleic Acids Res. 33, W36–W38 (2005).
Wagner, M., Adamczak, R., Porollo, A. & Meller, J. Linear regression models for solvent accessibility prediction in proteins. J. Comput. Biol. 12, 355–369 (2005).
Adamczak, R., Porollo, A. & Meller, J. Combining prediction of secondary structure and solvent accessibility in proteins. Proteins 59, 467–475 (2005).
Kota, P., Summers, D. W., Ren, H. Y., Cyr, D. M. & Dokholyan, N. V. Identification of a consensus motif in substrates bound by a Type I Hsp40. Proc. Natl Acad. Sci. USA 106, 11073–11078 (2009).
Jurica, M. S. et al. The allosteric regulation of pyruvate kinase by fructose-1,6-bisphosphate. Structure 6, 195–210 (1998).
Pieper, U. et al. ModBase, a database of annotated comparative protein structure models, and associated resources. Nucleic Acids Res. 39, D465–D474 (2011).
Vizcaino, J. A. et al. ProteomeXchange provides globally coordinated proteomics data submission and dissemination. Nat. Biotechnol. 32, 223–226 (2014).
Acknowledgements
We thank N. Stoynov for his support for the mass spectrometry analyses, all colleagues cited in Methods who provided reagents and a previous anonymous reviewer who suggested investigating Rsp5. This work was supported by a CIHR grant. R.J.D. is an HHMI investigator and T.M. is a CIHR and MSFHR new investigator.
Author information
Authors and Affiliations
Contributions
N.N.F. designed most of the experiments through discussions with T.M. and additional inputs from J.G., R.J.D. and D.R.; N.N.F. carried out most experiments; G.T.C. carried out the computational analyses with additional participation from J.G.; M.Z. and S.A.C. prepared several plasmids and strains; A.P. and N.N.F carried out the mammalian cells experiments together. D.R. provided reagents. Development of the conjugate assay was initiated in R.J.D.’s laboratory. T.M. supervised the study. N.N.F. and T.M. wrote the paper and all other authors edited or commented on the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Integrated supplementary information
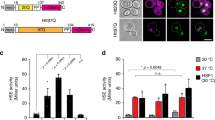
Supplementary Figure 1 Rsp5 is required for the increased ubiquitination induced by HS.
(a) Dot-blot images used for the quantification in Fig. 1a. (b) Normalized ubiquitination levels quantified by dot-blot in WT and rsp5-1 cells incubated at the indicated temperatures for 15 min. Increases in ubiquitination from 25 °C to the HS conditions in rsp5-1 cells were each compared to WT with a two-tailed student t-test (∗∗P < 0.01; ∗∗∗P < 0.001). (c) Ubiquitination levels in WT and rsp5-1 cells incubated at 45 °C for the indicated times were assessed by western-blot. (d) Normalized ubiquitination levels quantified by dot-blot in WT, rsp5-1, rsp5-sm1 and rps5-3 cells under the indicated conditions. Increase in ubiquitination from 37 °C to 45 °C was compared to WT with a two-tailed student t-test (∗∗P < 0.01; ∗P < 0.05). (e) Ubiquitination levels of Tetp::Rsp5 cells treated with (OFF) or without (ON) 100 μg/ml doxycycline before and after heat-shock were assessed by both dot-blot assay (bottom; quantified in Fig. 1b) and western-blot (top). Experiments in a, b, d and e were done with three biological replicates and when shown the averaged values are shown with standard deviations. a.u.: denotes arbitrary units (each value is relative to the reference sample). The source values are listed in Supplementary Table 4. (f) Ubiquitination levels in WT and rsp5-1 cells with an empty plasmid (-) or a plasmid that expressed His8-tagged ubiquitin from the GPD1 promoter were assessed by western-blot. Before analysis cells were subjected or not to a HS (45 °C for 15 min). (g) In vitro ubiquitination assay performed with MYC-ubiquitin in WT and rsp5-1 cell extracts incubated at 25 °C, 42 °C or 45 °C for 10 min and analysed by western blot using anti-MYC antibodies. Pgk1 loading control is also shown. Lower increased in poly-ubiquitination was observed at 45 °C, presumably due to the observed increase in protein precipitation in these conditions (data not shown).
Supplementary Figure 2 Lower levels of Nedd4 affect the increased ubiquitination but not cell viability on HS.
(a) Levels of Nedd4 were visualized by western blot after loading the same protein amounts derived from stably transfected HeLa cells with the indicated shRNA (− stands for scrambled control, #A and #B express two different hairpins that target Nedd4). (b) The lysates in Fig. 2a, were re-analysed by western blot on a 15% SDS-PAGE to visualize free mono-ubiquitin. (c) Viability of the indicated cells was assessed by mixing cells with 0.4% trypan blue stain and then counting with a CountessTM Automated Cell Counter (Invitrogen) after the indicated heat-stress in three independent HS assays. (d) Nedd4 (+/+) or (−/−) MEFs were incubated at the indicated temperatures at the noted times and ubiquitination levels were analysed by immunoblotting. On quantification, ubiquitination levels were higher at 45 °C (data not shown), and hence that temperature was used for the subsequent experiments.
Supplementary Figure 3 Rsp5 ubiquitinates mainly cytosolic proteins on HS without the help of arrestin-containing proteins.
(a) Summary table of ubiquitination sites (GG peptides) and proteins in two independent SILAC experiments. Experiment 1 (Exp1) analyses are also shown in Fig. 3b and d. (b) Comparison of SILAC ratios for proteins identified in both the whole cell lysate (WCL) and after the GG peptide enrichment. 523 proteins with at least two unique peptides were identified and 446 proteins quantified in WCL (only lysine was SILAC-labelled in this experiment). Protein levels were quantified in WCL for 137 ubiquitinated peptides. The log2 ratio values of light (WT, HS) versus medium (WT, no HS) SILAC-labelled proteins are compared in the top graph, and light versus heavy (rsp5-1, HS) in the lower graph. The ratio values of the ubiquitinated peptides are shown in the x axis, while the ratio values of the corresponding proteins in the WCL are shown in the y-axis. There was one Rsp5 candidate substrate protein (Sec27) that was more abundant in WT cells in comparison to rsp5-1 cells; in this case the observed decrease in ubiquitination in rsp5-1 cells may be attributed to lower protein levels. A few more Rsp5 candidate substrate proteins were also slightly more abundant in WCL (New1, Pbp1, Glt1, Gln1 and Cdc48), but their GG ratios were greater, thus the changes in ubiquitination levels cannot be fully attributed to changes in protein levels and correspond to lesser conjugation in rsp5-1 cells. (c) Immunoprecipitation (IP) of the indicated C-terminally 13xMYC tagged proteins expressed from a plasmid under a GDP promoter in heat-shock (20 min, 45 °C) treated Tetp::Rsp5 cells pre-treated for about 12 h with (OFF) or without (ON) 100 μg/ml doxycycline. Samples were analysed by western blotting as indicated. Ubiquitination signals were quantified in the molecular weight regions indicated by grey dotted lines. Signals were normalized to Pgk1 levels after background subtraction from the control lane. For comparison the Hul5 substrate Lsm7 was also included. Ubiquitination of Cdc19 and Pdc1 was also assessed using a MYC tagged ubiquitin, for which there was a lower background in control lanes (Fig. 4c, d). (d) Localization of HS-induced Rsp5 candidate substrates identified in the second SILAC-diGly mass spectrometry experiment shown in a. Distribution of proteins in each compartment is shown in percentage. C: Cytoplasm; N: Nuclear; M: Membrane; Mit: Mitochondria. (e) Tet::RSP5 cells expressing N-terminal GFP tagged Rsp5 with or without a NLS or an empty HIS plasmid were pre-treated for about 12 h with 100 μg/ml doxycycline (OFF). Ubiquitination levels of cells heat-shocked or not for 15 min at 45 °C were assessed by dot-blot (left). Fluorescent microscopy images of representative GFPRsp5 and GFP–NLSRsp5 expressing cells are shown on the right with the DNA stained with Hoechst. Scale bar represents 2 μm. (f) Increased ubiquitination levels in WT, art1-10Δ, and art1-8,10Δ cells from 25 °C to 45°C were assessed by dot-blot. Ubiquitination levels were normalized to Pgk1 levels. (g) Normalized ubiquitination levels before and after heat-shock in WT and rsp5-1 cells containing the indicated plasmids (− denotes presence of a control empty plasmid) were quantified by dot-blot. Both RSP5 and rsp5-C2Δ were expressed under a Gal-promoter induced for 60 min with 2% Galactose at 37°C. Experiments in e–g were done with three biological replicates and the averaged values are shown with standard deviations; a.u. denotes arbitrary units (each value is relative to the reference sample). The source values are listed in Supplementary Table 5.
Supplementary Figure 4 Rsp5 and Hul5 are mainly required for the increased ubiquitination of different proteins.
(a) Increased ubiquitination levels quantified by dot-blot after a 15 min HS at 45 °C in the indicated cells. Cells were pre-incubated at 37 °C for 30 min before the HS. Experiments were done with three biological replicates and the averaged values are shown with standard deviations; a.u. denotes arbitrary units (each value is relative to the reference sample). The source values are listed in Supplementary Table 4. (b) IMAC of samples from WT, hul5Δ and rsp5-1 cells that expressed His8-ubiquitin and Lsm7TAP from the same plasmid after a HS at 45 °C for 20 min were analysed by western-blot using anti-TAP antibodies. Relative levels of the poly-ubiquitin signal are indicated below. (c) IMAC-enriched ubiquitin conjugates were analysed by western blots. Samples were derived from indicated cells (rsp5-1 designated by green circles) expressing endogenously tagged candidates (3xHA) and H8-Ubiquitin. HS was applied to all samples in b and c.
Supplementary Figure 5 The heat-induced degradation of short-lived proteins requires functional proteasome.
(a–b) Degradation of 35S labelled proteins in WT (grey) and indicated mutant or deletion strains (green) cells at 25 °C (dotted lines) or 38 °C (solid lines). The portion of proteins degraded at the indicated times was measured for short-lived proteins (5 min labelling). The same values for WT samples are shown in a and b as all samples were analysed together. Experiments were done in three independent experiments and averaged values are shown with standard deviations. The source values are listed in Supplementary Table 5. (c) Increased ubiquitination levels from 25 °C to 45 °C (15 min) in WT and rsp5-1 cells treated with or without 100 μg/ml cycloheximide were analysed by dot-blot assay. Three biological replicates were analysed, averaged values are shown with standard deviation and source values are listed in Supplementary Table 4. a.u. denotes arbitrary units (each value is relative to the reference sample). (d) Protein localization of the cytoplasmic fraction sample obtained by native lysis and analysed by mass spectrometry. 348 proteins were identified in this experiment. The same lysis method was used for samples analysed in Fig. 5d. C: Cytoplasm; N: Nuclear; M: Membrane; Mit: Mitochondria.
Supplementary Figure 6 The Ydj1 Hsp40 is required for the increased ubiquitination induced by HS.
(a) Histograms of proteins with at least 1, 2 or 3 Ydj1 binding motifs among HS-induced Rsp5 candidate substrates and in the proteome. The Ydj1 binding motif GX[LMQ]{P}X{P}{CIMPVW} was previously identified57, P-values calculated using a Fishers exact test are shown, and assigned motifs are listed in Supplementary Table 4. (b) Western blot analysis of one of three lysates quantified by dot-blot in Fig. 5a. Ubiquitination levels were assessed after HS (45 °C, 15 min) in the indicated cells that carried a LEU2 plasmid that was empty or with YDJ1 or ydj1 (PP/GG) expressed from the YDJ1 promoter. Ubiquitination levels normalized to Pgk1 levels are indicated in grey and were quantified in the region indicated by the dotted line and. c. IMACs from samples derived from WT, ydj1Δ or rsp5-1 cells expressing His8-ubiquitin from a first plasmid and CDC19 (with C-terminal 13xMYC) in a second plasmid. Cells were HS (45 °C, 20 min) or not before lysis and IMACs were analysed by western blot using anti-MYC antibodies. The relative level of poly-ubiquitination is shown for each lane. Inputs and Pgk1 are also shown.
Supplementary Figure 7 A destabilizing mutation near a PY-motif of Cdc19 induces proteasome degradation.
(a) The predicted percentage of relative solvent accessibility was plotted for the PY motifs within structured regions of Rsp5 candidate substrates that are HS-induced (grey) or identified in a protein array (green). Solvent accessibility of motifs was predicted using Sable (http://sable.cchmc.org/) by averaging relative accessibility for the four PY residues (source data are listed in Supplementary Table 4). The median solvent accessibility of PY motifs in disordered regions in both datasets combined (presumably accessible) is marked with a dotted line for comparison. A box plot of the data is also shown above the plot. (b) Surface contour (upper left) of the crystal structure of the Cdc19 homo-dimer (1A3W): chains are depicted in green and cyan, PY motif (LPNY) in magenta (with the tyrosine side chain embedded in the structure), and D367 in orange. Enlargement (lower right) of the WT Cdc19 homo-dimer interface near D367 is showed. Backbones are depicted in cyan and pale green, sidechains in bright green and blue, and D367 in red. The predicted change caused by the D367R mutation in the interaction energy of the homo-dimer is also indicated. (c,d) IMACs from samples derived from ydj1Δ (c) or YDJ1 (d) cells expressing His8-ubiquitin from a first plasmid and the indicated Pdc1 constructs (with C-terminal 13xMYC) in a second plasmid. Cells were HS (45 °C, 20 min) before lysis and IMACs were analysed by western blot using anti-MYC antibodies. Inputs and Pgk1 are also shown. Note that Pdc1 may still be slightly ubiquitinated in rsp5-1 cells, presumably by Hul5 (Fig. 3f). (e) Western blots of samples derived from wild-type cells (BY4741) expresseing the indicated C-terminally 13xMYC tagged Cdc19 and that were shifted to 42 °C in the presence of 100 μg/ml cycloheximide (CHX) at the indicated time. The MYC band signals, normalized to the corresponding Pgk1 signals, are indicated below in grey. (f) Representative western blots used for quantification in Fig. 7g. (g–h) Western blots of samples derived from indicated cells that expressed the C-terminally 13xMYC tagged Cdc19(D367R) and that were shifted to 42 °C in the presence of 100 μg/ml cycloheximide (CHX) at the indicated time. The MYC band signals, normalized to the corresponding Pgk1 signals, are indicated below in grey. (i) The proportion of PY and PY-like motifs ([PLV]PxY, [PSV]PxF, PPPP or PPR) in proteins encoded in the yeast genome that are located in regions predicted to be disordered (IDP) or not (that is, structured) are indicated.
Supplementary Figure 8
Uncropped images of Western blots of main figures.
Supplementary information
Supplementary Information
Supplementary Information (PDF 1846 kb)
Supplementary Table 1
Supplementary Information (XLSX 64 kb)
Supplementary Table 2
Supplementary Information (XLSX 46 kb)
Supplementary Table 3
Supplementary Information (XLSX 11 kb)
Supplementary Table 4
Supplementary Information (XLSX 12 kb)
Supplementary Table 5
Supplementary Information (XLSX 548 kb)
Rights and permissions
About this article

Cite this article
Fang, N., Chan, G., Zhu, M. et al. Rsp5/Nedd4 is the main ubiquitin ligase that targets cytosolic misfolded proteins following heat stress. Nat Cell Biol 16, 1227–1237 (2014). https://doi.org/10.1038/ncb3054
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ncb3054
This article is cited by
-
Mitochondrial complexome reveals quality-control pathways of protein import
Nature (2023)
-
Mechanisms of genotoxicity and proteotoxicity induced by the metalloids arsenic and antimony
Cellular and Molecular Life Sciences (2023)
-
Global ubiquitinome profiling identifies NEDD4 as a regulator of Profilin 1 and actin remodelling in neural crest cells
Nature Communications (2022)
-
Yeast polyubiquitin unit regulates synaptonemal complex formation and recombination during meiosis
Journal of Microbiology (2022)
-
Disome-seq reveals widespread ribosome collisions that promote cotranslational protein folding
Genome Biology (2021)
