
Abstract
Diabetic retinopathy is an important cause of blindness in adults1,2, and is characterized by progressive loss of vascular cells and slow dissolution of inter-vascular junctions, which result in vascular leakage and retinal oedema3. Later stages of the disease are characterized by inflammatory cell infiltration, tissue destruction and neovascularization4,5. Here we identify soluble epoxide hydrolase (sEH) as a key enzyme that initiates pericyte loss and breakdown of endothelial barrier function by generating the diol 19,20-dihydroxydocosapentaenoic acid, derived from docosahexaenoic acid. The expression of sEH and the accumulation of 19,20-dihydroxydocosapentaenoic acid were increased in diabetic mouse retinas and in the retinas and vitreous humour of patients with diabetes. Mechanistically, the diol targeted the cell membrane to alter the localization of cholesterol-binding proteins, and prevented the association of presenilin 1 with N-cadherin and VE-cadherin, thereby compromising pericyte–endothelial cell interactions and inter-endothelial cell junctions. Treating diabetic mice with a specific sEH inhibitor prevented the pericyte loss and vascular permeability that are characteristic of non-proliferative diabetic retinopathy. Conversely, overexpression of sEH in the retinal Müller glial cells of non-diabetic mice resulted in similar vessel abnormalities to those seen in diabetic mice with retinopathy. Thus, increased expression of sEH is a key determinant in the pathogenesis of diabetic retinopathy, and inhibition of sEH can prevent progression of the disease.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others
References
Yau, J. W. Y. et al. Global prevalence and major risk factors of diabetic retinopathy. Diabetes Care 35, 556–564 (2012)
Bourne, R. R. A. et al. Causes of vision loss worldwide, 1990–2010: a systematic analysis. Lancet Glob. Health 1, e339–e349 (2013)
Antonetti, D. A., Klein, R. & Gardner, T. W. Diabetic retinopathy. N. Engl. J. Med. 366, 1227–1239 (2012)
Robinson, R., Barathi, V. A., Chaurasia, S. S., Wong, T. Y. & Kern, T. S. Update on animal models of diabetic retinopathy: from molecular approaches to mice and higher mammals. Dis. Model. Mech. 5, 444–456 (2012)
Klaassen, I., Van Noorden, C. J. F. & Schlingemann, R. O. Molecular basis of the inner blood-retinal barrier and its breakdown in diabetic macular edema and other pathological conditions. Prog. Retin. Eye Res. 34, 19–48 (2013)
Barber, A. J. et al. The Ins2Akita mouse as a model of early retinal complications in diabetes. Invest. Ophthalmol. Vis. Sci. 46, 2210–2218 (2005)
Hu, J. et al. Müller glia cells regulate Notch signaling and retinal angiogenesis via the generation of 19,20-dihydroxydocosapentaenoic acid. J. Exp. Med. 211, 281–295 (2014)
Hwang, S. H., Tsai, H. J., Liu, J. Y., Morisseau, C. & Hammock, B. D. Orally bioavailable potent soluble epoxide hydrolase inhibitors. J. Med. Chem. 50, 3825–3840 (2007)
Gerhardt, H. & Betsholtz, C. Endothelial–pericyte interactions in angiogenesis. Cell Tissue Res. 314, 15–23 (2003)
Hammes, H. P. et al. Pericytes and the pathogenesis of diabetic retinopathy. Diabetes 51, 3107–3112 (2002)
Enge, M. et al. Endothelium-specific platelet-derived growth factor-B ablation mimics diabetic retinopathy. EMBO J. 21, 4307–4316 (2002)
Gerhardt, H., Wolburg, H. & Redies, C. N-cadherin mediates pericytic-endothelial interaction during brain angiogenesis in the chicken. Dev. Dyn. 218, 472–479 (2000)
Navaratna, D., McGuire, P. G., Menicucci, G. & Das, A. Proteolytic degradation of VE-cadherin alters the blood-retinal barrier in diabetes. Diabetes 56, 2380–2387 (2007)
Gavard, J. & Gutkind, J. S. VEGF controls endothelial-cell permeability by promoting the β-arrestin-dependent endocytosis of VE-cadherin. Nat. Cell Biol. 8, 1223–1234 (2006)
Huster, D., Arnold, K. & Gawrisch, K. Influence of docosahexaenoic acid and cholesterol on lateral lipid organization in phospholipid mixtures. Biochemistry 37, 17299–17308 (1998)
Wassall, S. R. & Stillwell, W. Polyunsaturated fatty acid-cholesterol interactions: domain formation in membranes. Biochim. Biophys. Acta 1788, 24–32 (2009)
Cai, J. et al. PEDF regulates vascular permeability by a γ-secretase-mediated pathway. PLoS One 6, e21164 (2011)
Georgakopoulos, A. et al. Presenilin-1 forms complexes with the cadherin/catenin cell–cell adhesion system and is recruited to intercellular and synaptic contacts. Mol. Cell 4, 893–902 (1999)
Baki, L. et al. Presenilin-1 binds cytoplasmic epithelial cadherin, inhibits cadherin/p120 association, and regulates stability and function of the cadherin/catenin adhesion complex. Proc. Natl Acad. Sci. USA 98, 2381–2386 (2001)
Serban, G. et al. Cadherins mediate both the association between PS1 and β-catenin and the effects of PS1 on β-catenin stability. J. Biol. Chem. 280, 36007–36012 (2005)
Kouchi, Z . et al. p120 catenin recruits cadherins to γ-secretase and inhibits production of Ab peptide. J. Biol. Chem. 284, 1954–1961 (2009)
Saka, S. K. et al. Multi-protein assemblies underlie the mesoscale organization of the plasma membrane. Nat. Commun. 5, 4509 (2014)
Hulce, J. J., Cognetta, A. B., Niphakis, M. J., Tully, S. E. & Cravatt, B. F. Proteome-wide mapping of cholesterol-interacting proteins in mammalian cells. Nat. Methods 10, 259–264 (2013)
Morisseau, C. et al. Structural refinement of inhibitors of urea-based soluble epoxide hydrolases. Biochem. Pharmacol. 63, 1599–1608 (2002)
Pellissier, L. P. et al. Specific tools for targeting and expression in Müller glial cells. Mol. Ther. Methods Clin. Dev. 1, 14009 (2014)
Calabria, A. R., Weidenfeller, C., Jones, A. R., de Vries, H. E. & Shusta, E. V. Puromycin-purified rat brain microvascular endothelial cell cultures exhibit improved barrier properties in response to glucocorticoid induction. J. Neurochem. 97, 922–933 (2006)
Czupalla, C. J., Liebner, S. & Devraj, K. In vitro models of the blood-brain barrier. Methods Mol. Biol. 1135, 415–437 (2014)
Bess, E., Fisslthaler, B., Frömel, T. & Fleming, I. Nitric oxide-induced activation of the AMP-activated protein kinase α2 subunit attenuates IκB kinase activity and inflammatory responses in endothelial cells. PLoS One 6, e20848 (2011)
Kovacevic, I. et al. The F-BAR protein NOSTRIN participates in FGF signal transduction and vascular development. EMBO J. 31, 3309–3322 (2012)
Hammes, H. P., Martin, S., Federlin, K., Geisen, K. & Brownlee, M. Aminoguanidine treatment inhibits the development of experimental diabetic retinopathy. Proc. Natl Acad. Sci. USA 88, 11555–11558 (1991)
Barbosa-Sicard, E. et al. Inhibition of the soluble epoxide hydrolase by tyrosine nitration. J. Biol. Chem. 284, 28156–28163 (2009)
Rivera, J. et al. Measurement of 20-hydroxyeicosatetraenoic acid in human urine by gas chromatography-mass spectrometry. Clin. Chem. 50, 224–226 (2004)
Busse, R. & Lamontagne, D. Endothelium-derived bradykinin is responsible for the increase in calcium produced by angiotensin-converting enzyme inhibitors in human endothelial cells. Naunyn Schmiedebergs Arch. Pharmacol. 344, 126–129 (1991)
Paolinelli, R. et al. Wnt activation of immortalized brain endothelial cells as a tool for generating a standardized model of the blood brain barrier in vitro. PLoS One 8, e70233 (2013)
Taddei, A. et al. Endothelial adherens junctions control tight junctions by VE-cadherin-mediated upregulation of claudin-5. Nat. Cell Biol. 10, 923–934 (2008)
Xiao, K. et al. p120-Catenin regulates clathrin-dependent endocytosis of VE-cadherin. Mol. Biol. Cell 16, 5141–5151 (2005)
Fleming, I., Fisslthaler, B., Dixit, M. & Busse, R. Role of PECAM-1 in the shear-stress-induced activation of Akt and the endothelial nitric oxide synthase (eNOS) in endothelial cells. J. Cell Sci. 118, 4103–4111 (2005)
Vetrivel, K. S. et al. Association of γ-secretase with lipid rafts in post-Golgi and endosome membranes. J. Biol. Chem. 279, 44945–44954 (2004)
Acknowledgements
The authors thank K. Engel-Herbig, K. Bruch, N. Dietrich and I. Winter for expert technical assistance. This work was supported by the Deutsche Forschungsgemeinschaft (SFB-TR 23/A6 and B7, SFB 1118/C03, GRK 1874/1and Exzellenzcluster 147 ‘Cardio-Pulmonary Systems’) with partial support from NIEHS R01 ES002710. J.dM. was supported by the Deutsche Forschungsgemeinschaft Graduate School CLIC, and J.H. was the recipient of a research award from the Global Ophthalmology Awards Program from Bayer.
Author information
Authors and Affiliations
Contributions
J.H. and I.F. designed the experiments and analysed the data. J.H., S.D., S.-I.B., K.D., S.L. and R.P. performed experiments, S.Z. and K.A. generated the epoxide and diol profiles. J.L. and H.-P.H. generated and characterized the retinal digest preparations. Z.C., L.W., C.G.E., A.S. and S.F. characterized and provided human samples. J.d.M. and C.G. performed the NMR studies using isolated membranes, and T.F., A.J. and O.J.M. generated the adenoviruses and adeno-associated viruses. B.D.H. provided advice, feedback and the sEH inhibitor. J.H. and I.F. wrote the manuscript.
Corresponding author
Ethics declarations
Competing interests
J.H., T.F., R.P. and I.F. are authors of patent applications for the use of sEH inhibitors in the treatment of non-proliferative diabetic retinopathy (German Patent Application no. 10 2016 109 709.8, International PCT Patent Application no. PCT/EP2017/062618). S.F. is an employee of F. Hoffmann-La Roche Ltd, Basel, Switzerland. B.D.H. is author of the University of California patents on sEH inhibitors licensed to EicOsis. None of the other authors has any disclosures to declare.
Additional information
Reviewer Information Nature thanks T. Hla, M. Weingarth and the other anonymous reviewer(s) for their contribution to the peer review of this work.
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Figure 1 Characterization of Ins2Akita mice.
a–d, Wild-type and Ins2Akita littermates were treated with vehicle (Veh; 0.3% ethanol) or the sEH inhibitor from the age of 6 weeks. a, b, Body weight (a) and fasting blood glucose levels (b) were recorded at monthly intervals. c, d, Systolic blood pressure (SBP) (c) and heart rate (HR) (d) were assessed at the age of 12 months. e, sEH (red), glutamine synthetase (green) and GFAP (blue) expression in retinas from 12-month-old Ins2Akita mice and their wild-type littermates. Retinas from Ephx2−/− mice were used to demonstrate the specificity of the sEH antibody used. f, Retinal digest preparations from 3-month-old Ins2Akita mice and their non-diabetic wild-type littermates; acellular capillaries are marked by arrows. g–j, Quantitative retinal morphometry images shown in f; that is, endothelial cells (g), pericytes (h), extravascular pericytes (i) and acellular capillaries (j). k, Quantification of leaked FITC–BSA. l, Retinal digest preparations from 6-month-old Ins2Akita mice and their non-diabetic wild-type littermates. Acellular capillaries are marked by arrows. m–p, Quantitative retinal morphometry of images in l. q, Quantification of leaked FITC–BSA. Scale bars, 50 μm. n = 4 (a, b), 6 (e–q) or 8 (c, d) mice per group. Data are mean ± s.e.m. P values determined by two-way ANOVA (a–d), or Student’s two-tailed t-test (g–k, m–q).
Extended Data Figure 2 Epoxide and diol profile in retinas from wild-type and Ins2Akita mice and sEH expression in mice fed a high-fat diet.
a, Effect of diabetes and sEH inhibition on metabolites of polyunsaturated fatty acids in 12-month-old wild-type and Ins2Akita mice treated with either vehicle (0.3% ethanol) or sEH inhibitor. DHDP, dihydroxydocosapentaenoic acid; DHET, dihydroxyeicosatrienoic acid; DiHETE, dihydroxyeicosatetraenoic acid; DiHOME, dihydroxyoctadecenoic acid; EDP, epoxydocosapentaenoic acid; EET, epoxyeicosatrienoic acid; EpETE, epoxyeicosatetraenoic acid; EpOME, epoxyoctadecenoic acid; HEPE, hydroxyeicosapentaenoic acid; HODE, hydroxyoctadecadienoic acid; nd, not detectable. n = 6 biologically independent samples per group. b, Expression of epoxide hydrolase (Ephx) genes in retinas from 12-month-old wild-type and Ins2Akita littermates. c, Retinal sEH expression in wild-type mice given a normal diet (ND; n = 6) or a high fat, high carbohydrate (HFHC; n = 8) diet for 20 weeks. For gel source data, see Supplementary Fig. 1. d, sEH activity in retinas from mice given a normal diet (n = 6) or a high-fat, high-carbohydrate diet (n = 8). e, Retinal digest preparations from animals fed a normal diet or a high-fat, high-carbohydrate diet for 20 weeks. Acellular capillaries are marked by an arrow. f–i, Quantitative retinal morphometry of the images shown in e; that is, endothelial cells (f), pericytes (g), extravascular pericytes (h) and acellular capillaries (i). n = 6 mice per group (b, e–i). Data are mean ± s.e.m. P values determined by one-way ANOVA (a), or Student’s two-tailed t-test (b–d, f–i).
Extended Data Figure 3 Growth factor expression in the retina and lipid raft preparations.
a–m, Growth factor expression in retinas from wild-type and Ins2Akita littermates treated with vehicle (0.3% ethanol) or the sEH inhibitor. n = 6 biologically independent samples per group. n, o, Endothelial cells and pericytes were co-cultured and treated with solvent, 19,20-EDP or 19,20-DHDP in the presence of sEH inhibitor. Lipid rafts (LR) were isolated by gradient centrifugation (see Methods), and the distribution of VE-cadherin (n) and N-cadherin (o) were assessed by western blotting. Flotillin-1 (Flot1) was included as a marker of lipid-raft-enriched fractions. Similar results were obtained in three additional cell batches. p, q, Co-precipitation of p120 catenin with N-cadherin (p) and VE-cadherin (q) in endothelial cell–pericyte co-cultures after treatment with solvent, 19,20-EDP or 19,20-DHDP. VEGF was included as a positive control. Similar results were obtained in three additional experiments. r, VE-cadherin phosphorylation in endothelial cells treated with solvent, 19,20-EDP, 19,20-DHDP or VEGF for 5 or 15 min. For gel source data, see Supplementary Fig. 1. Comparable results were obtained in three additional experiments. Data are mean ± s.e.m. P values determined by two-way ANOVA.
Extended Data Figure 4 Mural cell coverage and N-cadherin localization in endothelial cell–pericyte co-cultures.
a, α-Smooth muscle actin (α-SMA) expression, showing mural cell coverage of 20-μm, 15-μm and 10-μm diameter retinal vessels. Scale bar, 50 mm. Comparable results were obtained in retinas from five additional animals in each group. b, Permeability of human endothelial cells to dextrans of different molecular mass; n = 6 independent cell preparations. c, VE-cadherin and N-cadherin expression in endothelial cells, pericytes or endothelial cell–pericyte co-cultures. Similar results were obtained in three different cell batches. For gel source data, see Supplementary Fig. 1. d, Cartoon showing the filter-based co-culture system studied. e, VE-cadherin (red), N-cadherin (green), desmin (cyan) and DAPI (grey) expression on transwell membranes with endothelial cell–pericyte co-cultures. Scale bars, 5 μm (top view) and 2 μm (side views). Similar observations were made in two additional experiments. f, g, Internalized N-cadherin (yellow) in pericytes treated with solvent, 19,20-EDP or 19,20-DHDP in the presence of sEH inhibitor; n = 5 independent experiments. Scale bar, 10 μm. Data are mean ± s.e.m. P values determined by one-way ANOVA.
Extended Data Figure 5 Association of cholesterol with cadherins and PS1.
a, Experimental pipeline for studying the association of proteins with cholesterol in Fig. 3n, o. b, Reverse immunoprecipitation for studying the association of proteins with cholesterol. Note the difference in steps 5 and 6 from experimental procedure in a. c–e, Representative LI-COR system images. Proteins clicked with cholesterol were visualized in green; N-cadherin (c), VE-cadherin (d) and PS1 (e) were detected on the same membrane and visualized in red. Comparable results were observed in three additional experiments.
Extended Data Figure 6 1H MAS NMR spectra from native brain membranes and epoxide-diol profile of retinas 14 days after adenovirus injection.
a, 1H MAS NMR spectra of membranes from wild-type and Ephx2−/− mouse brains. At low temperatures, ordered phases are predominant. They are characterized by a high chemical shift anisotropy and strong dipolar couplings resulting in broad components with low spectral resolution and pronounced spinning sidebands. With increasing temperatures, narrow components caused by fast axial motions and gauche-trans isomerization of the acyl chains dominate the centre band of the 1H MAS NMR spectrum. Therefore, the centre band CH2 resonance is a qualitative indicator of membrane dynamics and lipid order. b, Fatty acid epoxide and diol levels in retinas from wild-type animals 14 days after administration of adenovirus encoding GFP, wild-type sEH or an sEH mutant without epoxide hydrolase activity (∆EH); some of animals were treated with sEH inhibitor immediately after virus injection. n = 6 biologically independent samples per group. Data are mean ± s.e.m. P values determined by one-way ANOVA.
Extended Data Figure 7 sEH-induced retinopathy.
Desmin staining of non-diabetic wild-type retinas 14 days after administration of adenoviruses encoding GFP, wild-type sEH or sEH mutant. Some mice were treated with sEH inhibitor. Arrows indicate desmin-negative regions along a retinal artery. Comparable observations were made using five additional animals in each group.
Extended Data Figure 8 Retinopathy induced by adenovirus-mediated sEH overexpression in non-diabetic mice.
a, sEH (red) expression in retinas from wild-type mice 3 months after intravitreal injection of 1010 genomic copies of adenovirus encoding wild-type sEH (with Myc-tag, green). b, Retinal digest preparations from wild-type mice 3 months after intravitreal injection of 1010 genomic copies of adenovirus encoding GFP (CTL) or wild-type sEH. sEH inhibitor treatment began directly after adenovirus injection. Acellular capillaries are marked by arrows. c–f, Quantitative retinal morphometry of the images shown in b; that is, endothelial cells (c), pericytes (d), extravascular pericytes (e) and acellular capillaries (f). g, Desmin (red) and CD31 (cyan) expression in retinas from wild-type mice 3 months after intravitreal injection of adenoviruses. Scale bars, 50 μm. n = 6 (a, g) or 8 (b–f) mice per group. Data are mean ± s.e.m. P values determined by one-way ANOVA.
Supplementary information
Supplementary Figure 1
This file contains source data (gels) for Figures 1- 3, and Extended Data Figures 2- 4. (PDF 1690 kb)
Source data
Rights and permissions
About this article

Cite this article
Hu, J., Dziumbla, S., Lin, J. et al. Inhibition of soluble epoxide hydrolase prevents diabetic retinopathy. Nature 552, 248–252 (2017). https://doi.org/10.1038/nature25013
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature25013
This article is cited by
-
Modeling early pathophysiological phenotypes of diabetic retinopathy in a human inner blood-retinal barrier-on-a-chip
Nature Communications (2024)
-
Oxidative stress and epigenetics in ocular vascular aging: an updated review
Molecular Medicine (2023)
-
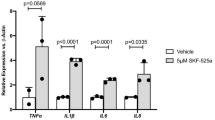
Cytochrome P450-epoxygenated fatty acids inhibit Müller glial inflammation
Scientific Reports (2021)
-
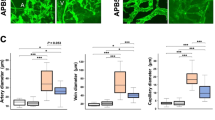
Altered bioavailability of epoxyeicosatrienoic acids is associated with conduit artery endothelial dysfunction in type 2 diabetic patients
Cardiovascular Diabetology (2019)
-
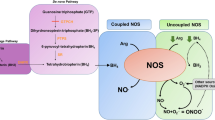
Role of ω3 polyunsaturated fatty acids in diabetic retinopathy: a morphological and metabolically cross talk among blood retina barriers damage, autoimmunity and chronic inflammation
Lipids in Health and Disease (2019)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
