
Abstract
Cyclin-dependent kinases 4 and 6 (CDK4/6) are fundamental drivers of the cell cycle and are required for the initiation and progression of various malignancies1,2. Pharmacological inhibitors of CDK4/6 have shown significant activity against several solid tumours3,4. Their primary mechanism of action is thought to be the inhibition of phosphorylation of the retinoblastoma tumour suppressor, inducing G1 cell cycle arrest in tumour cells5. Here we use mouse models of breast carcinoma and other solid tumours to show that selective CDK4/6 inhibitors not only induce tumour cell cycle arrest, but also promote anti-tumour immunity. We confirm this phenomenon through transcriptomic analysis of serial biopsies from a clinical trial of CDK4/6 inhibitor treatment for breast cancer. The enhanced anti-tumour immune response has two underpinnings. First, CDK4/6 inhibitors activate tumour cell expression of endogenous retroviral elements, thus increasing intracellular levels of double-stranded RNA. This in turn stimulates production of type III interferons and hence enhances tumour antigen presentation. Second, CDK4/6 inhibitors markedly suppress the proliferation of regulatory T cells. Mechanistically, the effects of CDK4/6 inhibitors both on tumour cells and on regulatory T cells are associated with reduced activity of the E2F target, DNA methyltransferase 1. Ultimately, these events promote cytotoxic T-cell-mediated clearance of tumour cells, which is further enhanced by the addition of immune checkpoint blockade. Our findings indicate that CDK4/6 inhibitors increase tumour immunogenicity and provide a rationale for new combination regimens comprising CDK4/6 inhibitors and immunotherapies as anti-cancer treatment.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others


Accession codes
References
Yu, Q. et al. Requirement for CDK4 kinase function in breast cancer. Cancer Cell 9, 23–32 (2006)
Choi, Y. J. et al. The requirement for cyclin D function in tumor maintenance. Cancer Cell 22, 438–451 (2012)
Patnaik, A. et al. Efficacy and safety of abemaciclib, an inhibitor of CDK4 and CDK6, for patients with breast cancer, non-small cell lung cancer, and other solid tumors. Cancer Discov. 6, 740–753 (2016)
Finn, R. S. et al. Palbociclib and letrozole in advanced breast cancer. N. Engl. J. Med. 375, 1925–1936 (2016)
Sherr, C. J. & Roberts, J. M. CDK inhibitors: positive and negative regulators of G1-phase progression. Genes Dev. 13, 1501–1512 (1999)
Goel, S. et al. Overcoming therapeutic resistance in HER2-positive breast cancers with CDK4/6 inhibitors. Cancer Cell 29, 255–269 (2016)
Cerami, E. et al. The cBio cancer genomics portal: an open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2, 401–404 (2012)
Campisi, J. & d’Adda di Fagagna, F. Cellular senescence: when bad things happen to good cells. Nat. Rev. Mol. Cell Biol. 8, 729–740 (2007)
Ma, C. X . et al. NeoPalAna: neoadjuvant palbociclib, a cyclin-dependent kinase 4/6 inhibitor, and anastrozole for clinical stage 2 or 3 estrogen receptor-positive breast cancer. Clin. Cancer Res. (2017)
Roulois, D. et al. DNA-demethylating agents target colorectal cancer cells by inducing viral mimicry by endogenous transcripts. Cell 162, 961–973 (2015)
Kimura, H., Nakamura, T., Ogawa, T., Tanaka, S. & Shiota, K. Transcription of mouse DNA methyltransferase 1 (Dnmt1) is regulated by both E2F-Rb-HDAC-dependent and -independent pathways. Nucleic Acids Res. 31, 3101–3113 (2003)
Chiappinelli, K. B. et al. Inhibiting DNA methylation causes an interferon response in cancer via dsRNA including endogenous retroviruses. Cell 162, 974–986 (2015)
Coppé, J. P., Desprez, P. Y., Krtolica, A. & Campisi, J. The senescence-associated secretory phenotype: the dark side of tumor suppression. Annu. Rev. Pathol. 5, 99–118 (2010)
Malumbres, M. et al. Mammalian cells cycle without the D-type cyclin-dependent kinases Cdk4 and Cdk6. Cell 118, 493–504 (2004)
Obata, Y. et al. The epigenetic regulator Uhrf1 facilitates the proliferation and maturation of colonic regulatory T cells. Nat. Immunol. 15, 571–579 (2014)
Bauer, C. A. et al. Dynamic TReg interactions with intratumoral APCs promote local CTL dysfunction. J. Clin. Invest. 124, 2425–2440 (2014)
Sakuishi, K. et al. Targeting Tim-3 and PD-1 pathways to reverse T cell exhaustion and restore anti-tumor immunity. J. Exp. Med. 207, 2187–2194 (2010)
Finn, R. S. et al. PD 0332991, a selective cyclin D kinase 4/6 inhibitor, preferentially inhibits proliferation of luminal estrogen receptor-positive human breast cancer cell lines in vitro. Breast Cancer Res. 11, R77 (2009)
Heng, T. S. et al. The Immunological Genome Project: networks of gene expression in immune cells. Nat. Immunol. 9, 1091–1094 (2008)
Bates, G. J. et al. Quantification of regulatory T cells enables the identification of high-risk breast cancer patients and those at risk of late relapse. J. Clin. Oncol. 24, 5373–5380 (2006)
McAllister, S. S. et al. Systemic endocrine instigation of indolent tumor growth requires osteopontin. Cell 133, 994–1005 (2008)
Wong, C. S. et al. Vascular normalization by loss of Siah2 results in increased chemotherapeutic efficacy. Cancer Res. 72, 1694–1704 (2012)
Bellone, M. et al. Relevance of the tumor antigen in the validation of three vaccination strategies for melanoma. J. Immunol. 165, 2651–2656 (2000)
Ni, J. et al. Combination inhibition of PI3K and mTORC1 yields durable remissions in mice bearing orthotopic patient-derived xenografts of HER2-positive breast cancer brain metastases. Nat. Med. 22, 723–726 (2016)
Wang, Y. et al. CDK7-dependent transcriptional addiction in triple-negative breast cancer. Cell 163, 174–186 (2015)
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550 (2014)
Subramanian, A. et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl Acad. Sci. USA 102, 15545–15550 (2005)
Mootha, V. K. et al. PGC-1α-responsive genes involved in oxidative phosphorylation are coordinately downregulated in human diabetes. Nat. Genet. 34, 267–273 (2003)
Gao, J. et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci. Signal. 6, pl1 (2013)
Gimenez, J. et al. Comparative methylation of ERVWE1/syncytin-1 and other human endogenous retrovirus LTRs in placenta tissues. DNA Res. 16, 195–211 (2009)
Acknowledgements
This work was supported by a Career Development Award from the DF/HCC SPORE in Breast Cancer (National Institutes of Health (NIH) 2015 P50 CA, to S.G.), a Landry Cancer Biology Research Fellowship (to M.J.D.), a DOD Era of Hope award (W81XWH-14-1-0191 to S.S.M.), NIH (NCI) RO1 CA166284 (to S.S.M.), a Presidential Early Career Award for Scientists and Engineers (to S.S.M.), the Breast Cancer Research Foundation (to J.J.Z.), the DF/HCC SPORE in Breast Cancer (P50 CA168504, to I.E.K., E.P.W., T.M.R. and J.J.Z.), and NIH awards CA187918-02 (to T.M.R. and J.J.Z.), CA210057-01 (to J.J.Z.), and CA172461-04 (to J.J.Z.). We thank T. Laszewski, A. Molineaux, J. Almeida, the Nikon Imaging Center at Harvard Medical School, and Boston Children’s Hospital Heme/Onc-HSCI Flow Cytometry facility for experimental assistance. We thank providers of reagents: pHAGE-deltaOVA-zsGreen plasmid (R. E. Tay, K. Wucherpfennig), LCMV-P14 transgenic mice (A. Cartwright, K. Wucherpfennig), CT-26 cell line (S. Elledge), and MMTV-PyMT-S2WTP3 cell line (A. Möller). We thank S. Stewart for advice.
Author information
Authors and Affiliations
Contributions
S.G., M.J.D., H.-J. K., S.S.M., and J.J.Z. designed the study. S.G. and M.J.D. performed all experiments with help as follows: A.C.W. and H.B. helped with in vitro tumour cell experiments, qPCR, in vivo treatments, and tumour tissue collection; H.-J. K. helped with TReg isolation from mice; J.S. and J.M.U. helped with processing mouse tissues for flow cytometry; N.K. helped with qPCR and combined bisulfite restriction analysis; B.B.L. analysed mouse transcriptomic data; S.R. conducted digital image analysis; S.X. conducted transcriptomic assays; I.E.K., E.P.W, O.M.-F., and T.M.R contributed to scientific discussion that guided the project’s direction; M.E., C.X.M., and J.H. conducted the NeoPalAna trial and provided gene expression data for analysis. S.G. and M.J.D. wrote the paper.
Corresponding authors
Ethics declarations
Competing interests
S.G. has served as a paid scientific adviser to Eli Lilly, and conducts laboratory research funded by Eli Lilly. Eli Lilly did not fund the present study. M.J.E. has performed ad hoc consulting for Novartis, Pfizer, and AstraZeneca, receives royalties for PAM50-based diagnostics including Prosigna, and holds stock in Bioclassifier LLC for PAM50-based diagnostics. C.X.M. receives research funding from Pfizer and has served as a paid scientific adviser to Pfizer and Novartis.
Additional information
Reviewer Information Nature thanks J. Balko, C. Klebanoff and the other anonymous reviewer(s) for their contribution to the peer review of this work.
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Figure 1 Tumour cell proliferation and expression of cell-cycle-related genes after CDK4/6 inhibition.
a, Immunohistochemistry for Ki-67 in MMTV-rtTA/tetO-HER2 tumours treated for 12 days with abemaciclib or vehicle; representative images (scale bar, 100 μm) and quantification (n = 7 tumours per group). b, Expression of E2F transcription factors, S phase genes, and G2/M genes in MMTV-rtTA/tetO-HER2 tumours treated with abemaciclib for 12 days compared with vehicle (vehicle, n = 11; abemaciclib, n = 12 tumours). c, d, GO terms with P < 0.05 (c) or GSEA terms significantly downregulated (d) by abemaciclib compared with vehicle in MMTV-rtTA/tetO-HER2 tumours (vehicle, n = 11; abemaciclib, n = 12 tumours). Unpaired two-tailed t-tests (a, b). Error bars, s.d. *P < 0.05, **P < 0.01.
Extended Data Figure 2 CDK4/6 inhibition enhances antigen presentation.
a, Antigen processing and presentation gene expression in MDA-MB-361 cells treated with 500 nM abemaciclib or DMSO (7 days; n = 3). b, Gene expression in cell lines treated with DMSO or palbociclib (7 days; n = 3). c, β2M/MHC-I flow cytometry in cell lines; grey, FMO control. (For MDA-MB-453, vehicle and abemaciclib, n = 2; palbociclib, n = 3. For MDA-MB-361, n = 3.) d, Gene expression in TCGA samples (CCND1 shallow deletion, n = 101; CCND1 diploid, n = 503; CCND1 gain, n = 203; CCND1 amplified, n = 153). e, H-2Kb SIINFEKL flow cytometry after 7 days of abemaciclib or DMSO (B16-OVA, n = 9; MMTV-PyMT-S2WTP3-OVA, n = 3). f, CD8+ T-cell proliferation in response to abemaciclib-pretreated tumour cells (n = 3). g, IFN-γ and TNF-α production in tumour cell/OT-I assay by ELISA (n = 3). One-way ANOVA adjusted for multiple comparisons (c, d, f), unpaired two-tailed t-tests (a, b, e, g). Error bars, s.d. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001.
Extended Data Figure 3 Effects of CDK4/6 inhibition on breast cancer cell proliferation and apoptosis in vitro.
a, Relative numbers of breast cancer cells cultured in 250 nM (MDA-MB-453) or 500 nM (MDA-MB-361, BT474) abemaciclib or DMSO for 11 days, followed by drug withdrawal (arrow). b, Representative senescence-associated β-galactosidase staining of MDA-MB-453 cells (left) and BT474 cells (right) treated with DMSO or abemaciclib (MDA-MB-453, 250 nM; BT474, 500 nM) for 0, 4, and 7 days. c, Western blot of SKBR3, BT474, MDA-MB-453, and MDA-MB-361 cells treated with DMSO, lapatinib, or abemaciclib for 48 h. d, Western blot of MDA-MB-453 cells pretreated with DMSO or abemaciclib (500 nM) for 0, 1, or 7 days before exposure to staurosporine (500 nM) for 4 h. For western blot source images, see Supplementary Fig. 1.
Extended Data Figure 4 CDK4/6 inhibition increases IFN signalling.
a, b, Top-ranked GO terms (a) and expression of IFN-responsive transcription factors (b) in MDA-MB-361 cells treated with 500 nM abemaciclib or DMSO (7 days; n = 3). c, Expression of ISGs in MMTV-PyMT-S2WTP3 cells treated with DMSO or abemaciclib (500 nM, 7 days) (n = 3). d, Expression of ISGs in MDA-MB-361 and MCF7 cells treated with abemaciclib (100 nM), fulvestrant (100 nM), or the combination for 7 days (n = 3). e, Expression of ISGs in MDA-MB-453, MCF7, and MDA-MB-231 cells treated with abemaciclib or DMSO (7 days; n = 3). f, Expression of ISGs in PDX 14-07 tumours treated with abemaciclib or vehicle (21–28 days; vehicle, n = 4; abemaciclib, n = 2 tumours per group). g, Phospho- and total STAT1 in cells treated with 500 nM abemaciclib as indicated. h, Confirmation of p16-Flag overexpression in MDA-MB-453 and BT474 cells (left) and gene expression in these cell lines by qPCR (right) (n = 6). i, j, Gene expression changes in MMTV-rtTA/tetO-HER2 tumours from mice treated with vehicle or abemaciclib for 12 days (vehicle, n = 11; abemaciclib, n = 12 tumours per group). Relative expression of IFN-responsive T-cell chemoattractants (i); relative expression of ISGs (j). k, Correlation of expression of Stat1 and Nlrc5 with genes involved in antigen processing and presentation in MMTV-rtTA/tetO-HER2 tumours. Blue dots, vehicle-treated tumours; red dots, abemaciclib-treated tumours. (Symbol r is the Pearson product-moment correlation coefficient.) Unpaired two-tailed t-tests (b, d–f, h–j) adjusted for multiple comparisons (c). Error bars, s.d. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001. For western blot source images, see Supplementary Fig. 1.
Extended Data Figure 5 CDK4/6 inhibitors enhance expression of immune-related signatures in breast cancer patients.
a–c, NeoPalAna schema (a). ‘Endo. Rx’, endocrine therapy for breast cancer. Downregulated GSEA signatures after 2 weeks of palbociclib treatment (b); nominal P < 0.001, false discovery rate q < 0.001. Upregulated GSEA signatures after 2 weeks palbociclib treatment (c); nominal P < 0.001, false discovery rate q < 0.001 (C1D1, n = 34; C1D15, n = 29; surgery, n = 23).
Extended Data Figure 6 CDK4/6 inhibition mediates type III IFN production.
a, Phospho- and total STAT1 in MDA-MB-453 cells treated with abemaciclib with or without ruxolitinib for 7 days. b, Effect of neutralization of IFN-α or IFN-γ on STAT1 mRNA expression (n = 2–4). c, d, Impact of neutralization of IFN-α (c) and IFN-γ (d) on phospho- and total STAT1 protein in indicated cell lines. e, Expression of type III IFN genes in indicated cell lines treated with abemaciclib for 7 days compared with DMSO (n = 3). f, Type III IFN production measured by ELISA (7 days; n = 2). Unpaired two-tailed t-tests (e, f) adjusted for multiple comparisons (b). Error bars, s.d. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001. For western blot source images, see Supplementary Fig. 1.
Extended Data Figure 7 DNMT1 suppression mediates dsRNA response.
a, DNMT1 expression after treatment with abemaciclib (n = 3). b, DNMT1 protein expression after treatment with abemaciclib. c, DNMT3A expression after treatment with abemaciclib (500 nM) for 7 days (n = 3, except abemaciclib 24 h, n = 2). d, DNMT1 expression after abemaciclib or control in MMTV-rtTA/tetO-HER2 tumours, P = 0.05 (12 days; vehicle, n = 11; abemaciclib, n = 12 tumours per group). e, RB1 knockdown in MDA-MB-453 cells and its effect on indicated gene expression after 7 days of abemaciclib (500 nM) (two biological replicates each associated with three technical replicates). f, ERV3-1 methylation. g, ERV expression after abemaciclib or DMSO (7 days; n = 3). h, Relative dsRNA expression after 7 days of abemaciclib compared with DMSO (n = 3). i, j, Cytosolic pattern recognition receptors in cells (i) (7 days; n = 3) or PDX tumours (j) (21–28 days; vehicle, n = 4; abemaciclib, n = 2 tumours per group). k, Western blot of overexpression of DNMT1 in MDA-MB-453 cells and quantification of mRNA expression (n = 3). Unpaired two-tailed t-tests (d, e, g–k) adjusted for multiple comparisons (a). Error bars, s.d. *P ≤ 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001. For western blot source images, see Supplementary Fig. 1.
Extended Data Figure 8 Abemaciclib induces a ‘senescence-like’ phenotype without evidence of SASP.
a, Representative senescence-associated β-galactosidase staining (left) of MMTV-rtTA/tetO-HER2 tumours treated with vehicle or abemaciclib for 12 days (scale bar, 500 μm). Quantification of relative senescence-associated β-galactosidase positive area (right, n = 6 tumours per group). b, mRNA expression of SASP factors in MMTV-rtTA/tetO-HER2 tumours treated as in Fig. 1a. Il6 expression determined by qPCR (n = 10 tumours per group); Il1a and Il1b by transcriptome analysis (vehicle, n = 11; abemaciclib, n = 12 tumours per group). c, MDA-MB-453 and BT474 cells treated with DMSO or abemaciclib (500 nM) for 7 days, and expression determined by qPCR. d, mRNA expression of IL6 upon doxorubicin-induced senescence (n = 3). MDA-MB-453 and BT474 cells were treated with doxorubicin (200 nM) for 24 h, and mRNA extracted 3 days later for qPCR. Unpaired two-tailed t-tests (a, d). Error bars, s.d. **P < 0.01.
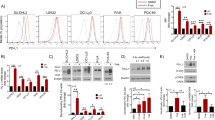
Extended Data Figure 9 Impact of CDK4/6 inhibition on immune cell populations and TReg biology.
a, Flow cytometric analysis of immune cell populations in MMTV-rtTA/tetO-HER2 tumours treated with vehicle or abemaciclib for 12 days (vehicle, n = 15; abemaciclib, n = 17 tumours per group). b, Peripheral blood TReg cells in MMTV-rtTA/tetO-HER2 mice (n = 4 mice per group). c, d, TReg cells (CD4+FoxP3+) quantified by flow cytometry of MMTV-PyMT tumours (c, vehicle, n = 18; abemaciclib, n = 16 tumours per group) and CT-26 tumours (d, n = 12 tumours per group) treated as indicated for 12 days. e, TReg cells in lymph nodes of tumour-free mice (12 days; one-way ANOVA corrected for multiple comparisons, vehicles and palbociclib, n = 8; abemaciclib, n = 7 mice per group). f, TReg:CD8 ratio in the spleens and lymph nodes (LN) of tumour-free FVB mice (12 days; vehicles and palbociclib, n = 8; abemaciclib, n = 7 mice per group). g, Plasma autoantibodies in tumour-free and tumour-bearing mice treated with vehicle or abemaciclib for 12 days (tumour-free vehicle, n = 8; tumour-free abemaciclib, n = 7; tumour-bearing vehicle, n = 7; tumour-bearing abemaciclib, n = 6 mice per group). h–l, Tumour-free FVB mice treated with abemaciclib or vehicle for 12 days. Thymic mass (h). Thymic cell populations were quantified by flow cytometry. CD4+CD8+ double-positive (DP) thymocytes (i); CD4+ single-positive (SP) thymocytes (j); CD8+ SP thymocytes (k); CD4+FoxP3+ regulatory T cells (l). (Vehicles and palbociclib, n = 5; abemaciclib, n = 4 mice per group.) m, Effect of abemaciclib or DMSO on ex vivo differentiation of CD4+CD25− T cells into TReg cells in the presence of TGF-β for 72 h (n = 4). n, Effect of DMSO or abemaciclib treatment for 72 h on TReg apoptosis measured by Annexin V staining (n = 2). o, Quantification of immunofluorescent staining of MMTV-rtTA/tetO-HER2 tumours (n = 7 tumours per group). p, q, Dnmt1 (p) and Cdkn1a (q) in T cells from tumour-free mice (12 days; two-way ANOVA corrected for multiple comparisons, n = 7 mice per group (pooled)). Unpaired two-tailed t-tests (b–d), one-way ANOVA corrected for multiple comparisons (e, f, h–l). Error bars, s.d. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001.
Extended Data Figure 10 Role of adaptive immunity in response to abemaciclib and combination with immunotherapy.
a, Relative change in volume of MMTV-rtTA/tetO-HER2 tumours implanted into Foxn1nu mice and then treated with vehicle (n = 11 tumours) or abemaciclib (n = 9 tumours) (one-way ANOVA). b, Immunohistochemistry of tumours in a for Ki-67. Representative images (left), and quantification of percentage of Ki-67+ cells (right) (vehicle, n = 7; abemaciclib, n = 10 per group; scale bar, 100 μm). c, Flow cytometric analysis of absolute number (left) and percentage (right) of CD8+ T cells in peripheral blood after administration of anti-CD8 or isotype control antibodies (isotype, n = 6; anti-CD8, n = 4 mice per group). d–g, Expression of inhibitory co-receptors on intratumoural CD8+ T cells in MMTV-rtTA/tetO-HER2 tumours after 6 days of treatment with abemaciclib (n = 6 tumours) or vehicle (n = 5 tumours). PD-1 cell-surface expression (d); representative flow cytometry plots (left), quantification (right). Representative flow cytometry plots for CTLA-4 (e) and LAG3 (f). g, Quantification of e and f. h, Quantification of number of inhibitory receptors per cell on intratumoural CD8+ T cells in MMTV-PyMT tumours after treating mice with abemaciclib or vehicle (12 days; n = 18 tumours per group). i, Ifng in MMTV-rtTA/tetO-HER2 tumours (12 days; Mann–Whitney test, n = 10 tumours per group). j, Experimental schema for Fig. 4j. k, Quantification of mean change in tumour volume at d19 (top) and maximal reduction in tumour volume (bottom) of curves in Fig. 4j. l, Spider plots of CT-26 tumour growth with indicated treatments (n = 4 per group, one experiment). m, Kaplan–Meier curves of incidence of CT-26 tumour formation after re-challenge (n = 4 per group). n, Schematic. Unpaired two-tailed t-tests (b, c), two-way ANOVA corrected for multiple comparisons (d, g). Error bars, s.d.; except (a), s.e.m. *P < 0.05, ***P < 0.001, ****P < 0.0001. For source data, see Supplementary Table 2.
Supplementary information
Supplementary information
This file contains the raw images of western blots and gating strategies for flow cytometry Supplementary Methods 1-3. (PDF 19115 kb)
Supplementary Table 1
Expression of SASP genes in MDA-MB-453 and MDA-MB-361 cells treated with abemaciclib (500 nM) for 7d relative to DMSO, and expression of SASP genes in MMTV-rtTA/tetO-HER2 tumors treated with abemaciclib for 12d relative to vehicle as determined by transcriptomic analysis. (XLSX 36 kb)
Supplementary Table 2
This table contains tumor volume and fold-change data from in vivo experiments. (XLSX 67 kb)
Rights and permissions
About this article

Cite this article
Goel, S., DeCristo, M., Watt, A. et al. CDK4/6 inhibition triggers anti-tumour immunity. Nature 548, 471–475 (2017). https://doi.org/10.1038/nature23465
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature23465
This article is cited by
-
ESCO2’s oncogenic role in human tumors: a pan-cancer analysis and experimental validation
BMC Cancer (2024)
-
MYC induces CDK4/6 inhibitors resistance by promoting pRB1 degradation
Nature Communications (2024)
-
Recent progress of CDK4/6 inhibitors’ current practice in breast cancer
Cancer Gene Therapy (2024)
-
The CDK4/6 inhibitor revolution — a game-changing era for breast cancer treatment
Nature Reviews Clinical Oncology (2024)
-
Unique vulnerability of RAC1-mutant melanoma to combined inhibition of CDK9 and immune checkpoints
Oncogene (2024)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
