
Abstract
In eukaryotes, the anaphase-promoting complex (APC/C, also known as the cyclosome) regulates the ubiquitin-dependent proteolysis of specific cell-cycle proteins to coordinate chromosome segregation in mitosis and entry into the G1 phase1,2. The catalytic activity of the APC/C and its ability to specify the destruction of particular proteins at different phases of the cell cycle are controlled by its interaction with two structurally related coactivator subunits, Cdc20 and Cdh1. Coactivators recognize substrate degrons3, and enhance the affinity of the APC/C for its cognate E2 (refs 4, 5, 6). During mitosis, cyclin-dependent kinase (Cdk) and polo-like kinase (Plk) control Cdc20- and Cdh1-mediated activation of the APC/C. Hyperphosphorylation of APC/C subunits, notably Apc1 and Apc3, is required for Cdc20 to activate the APC/C7,8,9,10,11,12, whereas phosphorylation of Cdh1 prevents its association with the APC/C9,13,14. Since both coactivators associate with the APC/C through their common C-box15 and Ile-Arg tail motifs16,17, the mechanism underlying this differential regulation is unclear, as is the role of specific APC/C phosphorylation sites. Here, using cryo-electron microscopy and biochemical analysis, we define the molecular basis of how phosphorylation of human APC/C allows for its control by Cdc20. An auto-inhibitory segment of Apc1 acts as a molecular switch that in apo unphosphorylated APC/C interacts with the C-box binding site and obstructs engagement of Cdc20. Phosphorylation of the auto-inhibitory segment displaces it from the C-box-binding site. Efficient phosphorylation of the auto-inhibitory segment, and thus relief of auto-inhibition, requires the recruitment of Cdk–cyclin in complex with a Cdk regulatory subunit (Cks) to a hyperphosphorylated loop of Apc3. We also find that the small-molecule inhibitor, tosyl-l-arginine methyl ester, preferentially suppresses APC/CCdc20 rather than APC/CCdh1, and interacts with the binding sites of both the C-box and Ile-Arg tail motifs. Our results reveal the mechanism for the regulation of mitotic APC/C by phosphorylation and provide a rationale for the development of selective inhibitors of this state.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others

Accession codes
Primary accessions
Electron Microscopy Data Bank
Protein Data Bank
Data deposits
EM maps are deposited in the Electron Microscopy Data Bank with accession codes 3385 (APC/CCdc20-Hsl1), 3386 (apo unphosphorylated APC/C), 3387 (apo phosphorylated APC/C), 3388 (combined apo phosphorylated APC/C), 3389 (APC/C∆Apc1-300s) and 3390 (APC/C∆Apc1-300s-TAME). Protein coordinates are deposited in the Protein Data Bank under accession codes 5G04 (APC/CCdc20-Hsl1) and 5G05 (apo phosphorylated APC/C).
References
Pines, J. Cubism and the cell cycle: the many faces of the APC/C. Nat. Rev. Mol. Cell Biol. 12, 427–438 (2011)
Primorac, I. & Musacchio, A. Panta rhei: the APC/C at steady state. J. Cell Biol. 201, 177–189 (2013)
Chang, L. & Barford, D. Insights into the anaphase-promoting complex: a molecular machine that regulates mitosis. Curr. Opin. Struct. Biol. 29, 1–9 (2014)
Kimata, Y., Baxter, J. E., Fry, A. M. & Yamano, H. A role for the Fizzy/Cdc20 family of proteins in activation of the APC/C distinct from substrate recruitment. Mol. Cell 32, 576–583 (2008)
Van Voorhis, V. A. & Morgan, D. O. Activation of the APC/C ubiquitin ligase by enhanced E2 efficiency. Curr. Biol. 24, 1556–1562 (2014)
Chang, L., Zhang, Z., Yang, J., McLaughlin, S. H. & Barford, D. Molecular architecture and mechanism of the anaphase-promoting complex. Nature 513, 388–393 (2014)
Lahav-Baratz, S., Sudakin, V., Ruderman, J. V. & Hershko, A. Reversible phosphorylation controls the activity of cyclosome-associated cyclin-ubiquitin ligase. Proc. Natl Acad. Sci. USA 92, 9303–9307 (1995)
Shteinberg, M., Protopopov, Y., Listovsky, T., Brandeis, M. & Hershko, A. Phosphorylation of the cyclosome is required for its stimulation by Fizzy/cdc20. Biochem. Biophys. Res. Commun. 260, 193–198 (1999)
Kramer, E. R., Scheuringer, N., Podtelejnikov, A. V., Mann, M. & Peters, J. M. Mitotic regulation of the APC activator proteins CDC20 and CDH1. Mol. Biol. Cell 11, 1555–1569 (2000)
Rudner, A. D. & Murray, A. W. Phosphorylation by Cdc28 activates the Cdc20-dependent activity of the anaphase-promoting complex. J. Cell Biol. 149, 1377–1390 (2000)
Golan, A., Yudkovsky, Y. & Hershko, A. The cyclin-ubiquitin ligase activity of cyclosome/APC is jointly activated by protein kinases Cdk1-cyclin B and Plk. J. Biol. Chem. 277, 15552–15557 (2002)
Kraft, C. et al. Mitotic regulation of the human anaphase-promoting complex by phosphorylation. EMBO J. 22, 6598–6609 (2003)
Zachariae, W., Schwab, M., Nasmyth, K. & Seufert, W. Control of cyclin ubiquitination by CDK-regulated binding of Hct1 to the anaphase promoting complex. Science 282, 1721–1724 (1998)
Jaspersen, S. L., Charles, J. F. & Morgan, D. O. Inhibitory phosphorylation of the APC regulator Hct1 is controlled by the kinase Cdc28 and the phosphatase Cdc14. Curr. Biol. 9, 227–236 (1999)
Schwab, M., Neutzner, M., Möcker, D. & Seufert, W. Yeast Hct1 recognizes the mitotic cyclin Clb2 and other substrates of the ubiquitin ligase APC. EMBO J. 20, 5165–5175 (2001)
Passmore, L. A. et al. Doc1 mediates the activity of the anaphase-promoting complex by contributing to substrate recognition. EMBO J. 22, 786–796 (2003)
Vodermaier, H. C., Gieffers, C., Maurer-Stroh, S., Eisenhaber, F. & Peters, J. M. TPR subunits of the anaphase-promoting complex mediate binding to the activator protein CDH1. Curr. Biol. 13, 1459–1468 (2003)
Patra, D. & Dunphy, W. G. Xe-p9, a Xenopus Suc1/Cks protein, is essential for the Cdc2-dependent phosphorylation of the anaphase- promoting complex at mitosis. Genes Dev. 12, 2549–2559 (1998)
Shteinberg, M. & Hershko, A. Role of Suc1 in the activation of the cyclosome by protein kinase Cdk1/cyclin B. Biochem. Biophys. Res. Commun. 257, 12–18 (1999)
Herzog, F., Mechtler, K. & Peters, J. M. Identification of cell cycle-dependent phosphorylation sites on the anaphase-promoting complex/cyclosome by mass spectrometry. Methods Enzymol. 398, 231–245 (2005)
Steen, J. A. et al. Different phosphorylation states of the anaphase promoting complex in response to antimitotic drugs: a quantitative proteomic analysis. Proc. Natl Acad. Sci. USA 105, 6069–6074 (2008)
Hegemann, B. et al. Systematic phosphorylation analysis of human mitotic protein complexes. Sci. Signal. 4, rs12 (2011)
Chang, L., Zhang, Z., Yang, J., McLaughlin, S. H. & Barford, D. Atomic structure of the APC/C and its mechanism of protein ubiquitination. Nature 522, 450–454 (2015)
Herzog, F. et al. Structure of the anaphase-promoting complex/cyclosome interacting with a mitotic checkpoint complex. Science 323, 1477–1481 (2009)
Izawa, D. & Pines, J. Mad2 and the APC/C compete for the same site on Cdc20 to ensure proper chromosome segregation. J. Cell Biol. 199, 27–37 (2012)
Yamaguchi, M. et al. Structure of an APC3-APC16 complex: insights into assembly of the anaphase-promoting complex/cyclosome. J. Mol. Biol. 427, 1748–1764 (2015)
van Zon, W. et al. The APC/C recruits cyclin B1-Cdk1-Cks in prometaphase before D box recognition to control mitotic exit. J. Cell Biol. 190, 587–602 (2010)
Sudakin, V., Shteinberg, M., Ganoth, D., Hershko, J. & Hershko, A. Binding of activated cyclosome to p13(suc1). Use for affinity purification. J. Biol. Chem. 272, 18051–18059 (1997)
Zeng, X. et al. Pharmacologic inhibition of the anaphase-promoting complex induces a spindle checkpoint-dependent mitotic arrest in the absence of spindle damage. Cancer Cell 18, 382–395 (2010)
Zhang, Z. et al. Recombinant expression, reconstitution and structure of human anaphase-promoting complex (APC/C). Biochem. J. 449, 365–371 (2013)
Perkins, D. N., Pappin, D. J., Creasy, D. M. & Cottrell, J. S. Probability-based protein identification by searching sequence databases using mass spectrometry data. Electrophoresis 20, 3551–3567 (1999)
Keller, A., Nesvizhskii, A. I., Kolker, E. & Aebersold, R. Empirical statistical model to estimate the accuracy of peptide identifications made by MS/MS and database search. Anal. Chem. 74, 5383–5392 (2002)
Bai, X. C., Fernandez, I. S., McMullan, G. & Scheres, S. H. Ribosome structures to near-atomic resolution from thirty thousand cryo-EM particles. eLife 2, e00461 (2013)
Li, X. et al. Electron counting and beam-induced motion correction enable near-atomic-resolution single-particle cryo-EM. Nat. Methods 10, 584–590 (2013)
Mindell, J. A. & Grigorieff, N. Accurate determination of local defocus and specimen tilt in electron microscopy. J. Struct. Biol. 142, 334–347 (2003)
Zhang, K. Gctf: real-time CTF determination and correction. J. Struct. Biol. 193, 1–12 (2016)
Scheres, S. H. RELION: implementation of a Bayesian approach to cryo-EM structure determination. J. Struct. Biol. 180, 519–530 (2012)
Scheres, S. H. Semi-automated selection of cryo-EM particles in RELION-1.3. J. Struct. Biol. 189, 114–122 (2015)
Scheres, S. H. Beam-induced motion correction for sub-megadalton cryo-EM particles. eLife 3, e03665 (2014)
Emsley, P. & Cowtan, K. Coot: model-building tools for molecular graphics. Acta Crystallogr. D 60, 2126–2132 (2004)
Tian, W. et al. Structural analysis of human Cdc20 supports multisite degron recognition by APC/C. Proc. Natl Acad. Sci. USA 109, 18419–18424 (2012)
Yang, Z. et al. UCSF Chimera, MODELLER, and IMP: an integrated modeling system. J. Struct. Biol. 179, 269–278 (2012)
Murshudov, G. N. et al. REFMAC5 for the refinement of macromolecular crystal structures. Acta Crystallogr. D 67, 355–367 (2011)
Fernández, I. S., Bai, X. C., Murshudov, G., Scheres, S. H. & Ramakrishnan, V. Initiation of translation by cricket paralysis virus IRES requires its translocation in the ribosome. Cell 157, 823–831 (2014)
Waterhouse, A. M., Procter, J. B., Martin, D. M., Clamp, M. & Barton, G. J. Jalview Version 2–a multiple sequence alignment editor and analysis workbench. Bioinformatics 25, 1189–1191 (2009)
Acknowledgements
This work was funded by the MRC Laboratory of Molecular Biology and a Cancer Research UK grant to D.B. PhD funding for S.Z. was from the Gates Cambridge Scholarship and Boehringer Ingelheim Fonds. C.A. is an EMBO Fellow. We are grateful to members of the Barford group for discussion; S. Chen, C. Savva and G. McMullan for EM facilities; J. Grimmet and T. Darling for computing; K. Zhang for advice on data processing; G. Murshudov for help with REFMAC and S. Aibara for advice on cloning.
Author information
Authors and Affiliations
Contributions
S.Z. cloned the substrates and Cdh1 mutant, purified proteins and performed biochemical analysis. S.Z. and L.C. prepared grids, collected and analysed EM data and determined the three-dimensional reconstructions. S.Z. fitted coordinates, built models and made figures with help of L.C. C.A. cloned kinases and Cdc20 and established in vitro phosphorylation of the APC/C. Z.Z. and J.Y. cloned the APC/C mutants and the chimaeric proteins and prepared viruses. S.M. and M.S. performed mass spectrometry. D.B. directed the project and designed experiments with S.Z. S.Z. and D.B. wrote the manuscript with input from authors.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Extended data figures and tables
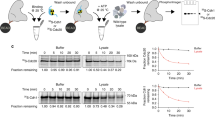
Extended Data Figure 1 Preparations and EM images of different APC/C samples used for structural studies.
a, Recombinant human APC/C was phosphorylated in vitro using Cdk2–cyclin A3, Cdk2–cyclin A3–Cks2 or Plk1 alone or with both Cdk2–cyclin A3–Cks2 and Plk1. The phosphorylated APC/C samples are shown after SDS–PAGE. b, In vitro phosphorylated recombinant human APC/C can be fully activated by Cdc20 to ubiquitylate a native substrate Cdk2–cyclin A2–Cks2 when both kinases were added (lanes 9, 10). Without Cks2 (lanes 3, 4) or with Plk1 alone (lanes 7, 8) no activation of the APC/C could be observed, whereas treating with Cdk2–cyclin A3–Cks2 alone (lanes 5, 6) resulted in its partial activation. Samples were recorded at 15 and 30 min of the reaction and 20 nM of Cdc20 was used. This experiment was replicated three times. Anti-Apc3 antibodies (BD Bioscience, cat. code: 610454) were used as a loading control. c, Purified wild-type (WT) APC/C and mutant samples with and without kinase treatment (both Cdk2–cyclin A3–Cks2 and Plk1). Upon deletion of the Apc3 loop, no association of the Cdk2–cyclin A3–Cks2 kinase to the APC/C could be observed (lanes 6 and 8). d, SDS–PAGE of purified APC/CCdc20-Hsl1 ternary complex. e, A typical cryo-EM micrograph of APC/CCdc20-Hsl1 representative of 15,582 micrographs. f, Gallery of two-dimensional averages of APC/CCdc20-Hsl1 showing different views; representative of 100 two-dimensional averages. g, Gold-standard FSC curves of all APC/C reconstructions in this work. See Supplementary Fig. 1 for gel source data.
Extended Data Figure 2 Three-dimensional classification of APC/CCdc20-Hsl.
The initial particles after two-dimensional classification were divided into six classes by three-dimensional classification using RELION. The resultant classes were grouped into four categories: (i) 9.0% in the active ternary state with coactivator and substrate bound; (ii) 11.3% in a hybrid state with coactivator bound, but the APC/C in the inactive conformation; (iii) 71.6% in the inactive apo state; (iv) 8.1% with poorer reconstruction owing to some bad particles. The first class in the active ternary state containing 179,660 particles was used for three-dimensional refinement and movie correction to obtain the final reconstruction at 3.9 Å.
Extended Data Figure 3 Comparison of Cdc20 and Cdh1 association to the APC/C.
a, The catalytic module (Apc2-Apc11) of the APC/CCdc20-Hsl1 complex is flexible and almost no density accounting for Apc11 (pink, modelled based on the structure of APC/CCdh1-Emi1, PDB 4UI9)23 could be observed. b, The WD40 domain of Cdc20 (purple) occupies a similar position as Cdh1WD40 (grey), but it is displaced from the APC/C by as much as 10 Å. c, d, EM density for Cdc20C box allowed for ab initio model building and the C-box interaction with Apc8B (cyan) is well conserved between the two coactivators. e, Both Cdc20IR (right) and Cdh1IR (left) associates with Apc3A (orange), although the EM density for Cdc20IR is much weaker (not shown) and the C-terminal α-helix in Cdh1IR is absent.
Extended Data Figure 4 Conformational changes of the APC/C between the inactive apo and the active ternary states and domain and sequence analysis of Cdc20.
a, b, Subunits that undergo conformational changes upon coactivator and substrate binding are highlighted in their ternary state and coloured as in Fig. 1, while the corresponding proteins in the inactive apo state are in lighter shades. In the active conformation, the platform subdomain containing subunits Apc1, Apc4 and Apc5 is shifted upward, inducing a large movement of the catalytic module to enable E2 access. c, Domain organization of Cdc20. d, Sequence alignment of Cdc20NTD and Cdh1NTD with α-helices represented as cylinders (purple and grey for Cdc20NTD and Cdh1NTD, respectively) underneath the sequences and the C-box and KILR/KLLR motif highlighted.
Extended Data Figure 5 Comparison of apo APC/C in unphosphorylated and phosphorylated states.
a, b, Superposition of the apo unphosphorylated (magenta) and phosphorylated (cyan) APC/C EM maps revealed little conformational differences except in the vicinity of the C-box binding site. c, Apc3A is in an equilibrium between open (light blue) and closed (orange) conformations. While in the inactive apo state, the majority of Apc3A is in the closed state, association of Cdc20IR stabilizes the open state. d, Sequence alignment of the Apc1 300s loop across different species human, mouse, Xenopus tropicalis (western clawed frog) and Danio rerio (zebrafish). Phosphorylation sites are indicated and residues accounting for the Apc1 auto-inhibitory segment (361–380) are boxed.
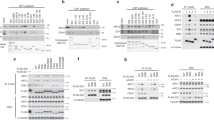
Extended Data Figure 6 Analytical gel filtration and activity assays.
a, With equal amount of input Cdc20, phosphorylated APC/C could form a stable binary complex with Cdc20 after a gel-filtration purification step (lane 5), whereas unphosphorylated APC/C could not (lane 4). b, Both unphosphorylated and phosphorylated APC/C associate with Cdh1 stably on gel filtration, as well as APC/C∆Apc1-300s. Anti Cdc20 antibody (Santa Cruz Biotechnology, cat. code: sc-8358) and anti Cdh1 antibody (Sigma, cat. code: C7855) were used for detection; antibody to Apc4 (ref. 6) served as a loading control and unphosphorylated APC/C alone is used as a negative control for western blotting. c, Point mutations of peptide 7 (residues 361–380), either when Arg368 was mutated to glutamate or when the four neighbouring serines were mutated to phospho-mimics (Glu), caused the peptide to abolish its inhibition effect and restored the APC/C activity (lanes 4, 5). Phosphorylation of a single Ser377 only resulted in partial activation of the APC/C (lane 6). d, Chimaeric proteins composed of the NTD, the WD40 domain and the IR tail of either Cdc20 or Cdh1 were purified to study their differences in APC/C activation. Both the NTD and the CTD of the coactivators are essential for their association with the APC/C. Swapping both NTD and CTD of Cdh1 with Cdc20 makes it phosphorylation sensitive (lanes 7, 8), similar to Cdc20 (lanes 9, 10) and vice versa. e, Top, Cdh1 can activate both unphosphorylated and phosphorylated APC/C similarly, whereas Cdc20 requires APC/C phosphorylation for its activity. Bottom, a titration of Cdh1 against unphosphorylated APC/C and APC/C∆Apc1-300s showed enhanced activity in the absence of the Apc1 auto-inhibitory segment at low Cdh1 concentration (≤10 nM), whereas Cdc20 requires displacement of the auto-inhibitory segment for its activity. f, Deletion of the Cdh1 α3 helix resulted in reduced activation of the APC/C and makes Cdh1 more phosphorylation sensitive. The substrate Cdk2–cyclin A2–Cks2 was used for assay in c and Hsl1 for the assays in d–f. 20 nM Cdc20 was used in c, 10 nM chimaeric coactivators in d and 30 nM coactivators in f. Experiments in a and b were replicated two times, in c, e and f three times and in d four times. See Supplementary Fig. 1 for gel source data.
Extended Data Figure 7 TAME competes with Cdc20 to bind at the IR-tail and the C-box binding sites.
a, TAME (C atoms in lime green) is superimposed with Cdc20IR (purple) and the arginine motif in both structures engages the same binding site on Apc3A (orange). b, The tosyl-Arg motif of TAME overlaps with Arg78–Tyr79 of Cdc20C box at the C-box binding site to out-compete Cdc20. c, A density for TAME was also observed within a pocket of the Apc8A TPR super-helix, similar to that of Apc8B.
Supplementary information
Supplementary Figure 1
Original source images for all data obtained by electrophoretic separation: Coomassie stained SDS PAGE and western blots. (PDF 5046 kb)
Structure of phosphorylated APC/CCdc20.Hsl1 and unphosphorylated apo APC/C (a narrative of the main results of the manuscript).
The chronology follows Figures 1 to 3. Overall structure of the phosphorylated APC/CCdc20. Detailed views showing interactions between Cdc20 and the APC/C (C box, KILR motif and IR tail) and comparison with Cdh1. The apo unphosphorylated APC/C is shown revealing the Apc1 auto-inhibitory (AI) segment that overlaps with the C box motif of Cdc20. Red spheres on the Apc1 AI segment indicate mitotic phosphorylation sites. (MP4 24230 kb)
Rights and permissions
About this article

Cite this article
Zhang, S., Chang, L., Alfieri, C. et al. Molecular mechanism of APC/C activation by mitotic phosphorylation. Nature 533, 260–264 (2016). https://doi.org/10.1038/nature17973
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature17973
This article is cited by
-
CDK9-55 guides the anaphase-promoting complex/cyclosome (APC/C) in choosing the DNA repair pathway choice
Oncogene (2024)
-
Alternative CDC20 translational isoforms tune mitotic arrest duration
Nature (2023)
-
Single-molecule analysis of specificity and multivalency in binding of short linear substrate motifs to the APC/C
Nature Communications (2022)
-
Ubiquitin signaling in cell cycle control and tumorigenesis
Cell Death & Differentiation (2021)
-
Structural basis of human separase regulation by securin and CDK1–cyclin B1
Nature (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
