
Abstract
Marine sediments from the North Pacific document two episodes of expansion and strengthening of the subsurface oxygen minimum zone (OMZ) accompanied by seafloor hypoxia during the last deglacial transition1,2,3,4. The mechanisms driving this hypoxia remain under debate1,2,3,4,5,6,7,8,9,10,11. We present a new high-resolution alkenone palaeotemperature reconstruction from the Gulf of Alaska that reveals two abrupt warming events of 4–5 degrees Celsius at the onset of the Bølling and Holocene intervals that coincide with sudden shifts to hypoxia at intermediate depths. The presence of diatomaceous laminations and hypoxia-tolerant benthic foraminiferal species, peaks in redox-sensitive trace metals12,13, and enhanced 15N/14N ratio of organic matter13, collectively suggest association with high export production. A decrease in 18O/16O values of benthic foraminifera accompanying the most severe deoxygenation event indicates subsurface warming of up to about 2 degrees Celsius. We infer that abrupt warming triggered expansion of the North Pacific OMZ through reduced oxygen solubility and increased marine productivity via physiological effects; following initiation of hypoxia, remobilization of iron from hypoxic sediments could have provided a positive feedback on ocean deoxygenation through increased nutrient utilization and carbon export. Such a biogeochemical amplification process implies high sensitivity of OMZ expansion to warming.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others

References
Mix, A. C. et al. Rapid climate oscillations in the Northeast Pacific during the last deglaciation reflect Northern and Southern Hemisphere sources, in Mechanisms of global climate change at millennial time scales, American Geophysical Union, edited by P.U. Clark et al., Geophysical Monograph 112, 127–148 (1999)
Davies, M. H. et al. The deglacial transition on the southeastern Alaska Margin: Meltwater input, sea level rise, marine productivity, and sedimentary anoxia. Paleoceanography 26, PA2223 (2011)
Behl, R. J. & Kennett, J. P. Brief interstadial events in the Santa Barbara basin, NE Pacific, during the past 60 kyr. Nature 379, 243–246 (1996)
Jaccard, S. L. & Galbraith, E. D. Large climate-driven changes in oceanic oxygen concentrations during the last deglaciation. Nature Geosci. 5, 151–156 (2012)
Okazaki, Y. et al. Deepwater formation in the North Pacific during the Last Glacial Termination. Science 329, 200–204 (2010)
Crusius, J., Pedersen, T. F., Kienast, S., Keigwin, L. & Labeyrie, L. Influence of northwest Pacific productivity on North Pacific Intermediate Water oxygen concentrations during the Bølling-Allerød interval (14.7–12.9 ka). Geology 32, 633–636 (2004)
Hendy, I. L., Pedersen, T. F., Kennett, J. P. & Tada, R. Intermittent existence of a southern Californian upwelling cell during submillennial climate change of the last 60 kyr. Paleoceanography 19, PA3007 (2004)
Schmittner, A., Galbraith, E. D., Hostetler, S. W., Pedersen, T. F. & Zang, R. Large fluctuations of dissolved oxygen in the Indian and Pacific oceans during Dansgaard-Oeschger oscillations caused by variations of North Atlantic Deep Water subduction. Paleoceanography 22, PA3207 (2007)
Kohfeld, K. E. & Chase, Z. Controls on deglacial changes in biogenic fluxes in the North Pacific ocean. Quat. Sci. Rev. 30, 3350–3363 (2011)
Lam, P. J. et al. Transient stratification as the cause of the North Pacific productivity spike during deglaciation. Nat. Geosci. 6, 622–626 (2013)
Kuehn, H. et al. Laminated sediments in the Bering Sea reveal atmospheric teleconnections to Greenland climate on millennial to decadal timescales during the last deglaciation. Clim. Past 10, 2215–2236 (2014)
Barron, J. A., Bukry, D., Dean, W. E., Addison, J. A. & Finney, B. Paleoceanography of the Gulf of Alaska during the past 15,000 years: results from diatoms, silicoflagellates, and geochemistry. Mar. Micropaleontol. 72, 176–195 (2009)
Addison, J. A. et al. Productivity and sedimentary δ15N variability for the last 17,000 years along the northern Gulf of Alaska slope. Paleoceanography 27, PA1206 (2012)
Keeling, R. F., Kortzinger, A. & Gruber, N. Ocean deoxygenation in a warming world. Annu. Rev. Mar. Sci . 2, 199–229 (2010)
Schmittner, A., Oschlies, A., Matthews, H. D. & Galbraith, E. D. Future changes in climate, ocean circulation, ecosystems and biogeochemical cycling simulated for a business-as-usual CO2 emission scenario until year 4000 AD. Glob. Biogeochem. Cycles 22, GB1013 (2008)
Behrenfeld, M. J. et al. Climate-driven trends in contemporary ocean productivity. Nature 444, 752–755 (2006)
Lopes, C., Kucera, M. & Mix, A. C. Climate change decouples oceanic primary and export productivity and organic carbon burial. Proc. Natl Acad. Sci. USA 112, 332–335 (2014)
Diaz, R. J. & Rosenberg, R. Spreading dead zone and consequences for marine ecosystems. Science 321, 926–929 (2008)
Scholz, F., McManus, J., Mix, A. C., Hensen, C. & Schneider, R. R. The impact of ocean deoxygenation on iron release from continental margin sediments. Nat. Geosci. 7, 433–437 (2014)
Davies-Walczak, M. H. et al. Late glacial to Holocene radiocarbon constraints on North Pacific Intermediate Water ventilation and deglacial atmospheric CO2 sources. Earth Planet. Sci. Lett. 397, 57–66 (2014)
Takeda, S. Iron and phytoplankton growth in the subarctic North Pacific. Aqua-BioScience Monographs 4, 41–93 (2011)
Benson, B. B. & Krause, D. J. The concentration and isotopic fractionation of oxygen dissolved in freshwater and seawater in equilibrium with the atmosphere. Limnol. Oceanogr. 29, 620–632 (1984)
Hofmann, A. F. et al. Hypoxia by degrees: establishing definitions for a changing ocean. Deep Sea Res. Part I Oceanogr. Res. Pap . 58, 1212–1226 (2011)
Ingall, E. & Jahnke, R. Evidence for enhanced phosphorus regeneration from marine sediments overlain by oxygen depleted waters. Geochim. Cosmochim. Acta 58, 2571–2575 (1994)
Boyd, P. & Newton, P. Evidence of the potential influence of planktonic community structure on the interannual variability of particulate organic carbon flux. Deep Sea Res. Part I Oceanogr. Res. Pap . 42, 619–639 (1995)
Hoffmann, L. J., Peeken, I., Lochte, K., Assmy, P. & Veldhuis, M. Different reaction of Southern Ocean phytoplankton size classes to iron fertilization. Limnol. Oceanogr. 51, 1217–1229 (2006)
Kemp, A. E. S. & Villareal, T. A. High diatom production and export in stratified waters – A potential negative feedback to global warming. Prog. Oceanogr. 119, 4–23 (2013)
Eppley, R. W. Temperature and phytoplankton growth in the sea. Fish Bull. 70, 1063–1085 (1972)
Sverdrup, H. U. On conditions for the vernal blooming of phytoplankton. ICES J. Mar. Sci. 18, 287–295 (1953)
Wang, M., Overland, J. E. & Bond, N. A. Climate projections for selected large marine ecosystems. J. Mar. Syst . 79, 258–266 (2010)
Schlitzer, R. Electronic Atlas of WOCE Hydrographic and Tracer Data Now Available. Eos Trans. AGU 81, 45 (2000)
Brickley, P. J. & Thomas, A. C. Satellite-measured seasonal and inter-annual chlorophyll variability in the Northeast Pacific and Coastal Gulf of Alaska. Deep Sea Res. Part II Top. Stud. Oceanogr . 51, 229–245 (2004)
Stabeno, P. J. et al. Meteorology and oceanography of the Northern Gulf of Alaska. Cont. Shelf Res. 24, 859–897 (2004)
Parnell, A. C. et al. A flexible approach to assessing synchroneity of past events using Bayesian reconstructions of sedimentation history. Quat. Sci. Rev. 27, 1872–1885 (2008)
Reimer, P. J. et al. INTCAL13 and MARINE13 radiocarbon age calibration curves, 0–50,000 years cal BP. Radiocarbon 55, 1869–1887 (2013)
Praetorius, S. K. & Mix, A. C. Synchronization of North Pacific and Greenland climates preceded abrupt deglacial warming. Science 345, 444–448 (2014)
Adkins, J. F. & Boyle, E. A. Changing atmospheric Δ14C and the record of deep water paleoventilation ages. Paleoceanography 12, 337–344 (1997)
Bemis, B. E., Spero, H. J., Bijima, J. & Lea, D. W. Reevaluation of the oxygen isotopic composition of planktonic foraminifera: Experimental results and revised paleotemperature equations. Paleoceanography 13, 150–160 (1988)
Waelbroeck, C. L. et al. Sea-level and deep water temperature changes derived from benthic foraminifera isotopic records. Quat. Sci. Rev. 21, 295–305 (2002)
Walinsky, S. E. et al. Distribution and composition of organic matter in surface sediments of coastal Southeast Alaska. Cont. Shelf Res. 29, 1565–1579 (2009)
Christie, W. W. & Han, X. Lipid Analysis: Isolation, Separation, Identification and Structural Analysis of Lipids 4th edn (Oily Press, 2003)
Prahl, F. G., Muehlhausen, L. A. & Zahnle, D. L. Further evaluation of long-chain alkenones as indicators of paleoceanographic conditions. Geochim. Cosmochim. Acta 52, 2303–2310 (1988)
Prahl, F. G. et al. Systematic pattern in – Temperature residuals for surface sediments from high latitude and other oceanographic settings. Geochim. Cosmochim. Acta 74, 131–143 (2010)
Rontani, J. F., Volkman, J. K., Prahl, F. G. & Wakeham, S. G. Biotic and abiotic degradation of alkenones and implications on paleoproxy application. A review. Org. Geochem . 59, 95–113 (2013)
Prahl, F. G., Cowie, G. L., De Lange, G. J. & Sparrow, M. G. Selective organic matter preservation in “burn-down” turbidites on the Madeira Abyssal Pla in. Paleoceanography 18, 1052 (2003)
Rontani, J. F. et al. Degradation of alkenones by aerobic heterotrophic bacteria: selective or not? Org. Geochem. 39, 34–51 (2008)
Barron, J. A., Heusser, L., Herbert, T. & Lyle, M. High-resolution climatic evolution of coastal northern California during the past 16,000 years. Paleoceanography 18, 1020 (2003)
Roselle-Melé, A. & Comes, P. Evidence for a warm Last Glacial Maximum in the Nordic seas or an example of shortcomings in UK37′ and UK37 to estimate low seas surface temperature? Paleoceanography 14, 770–776 (1999)
Bendle, J. & Roselle-Mele, A. Distributions of UK37 and UK37′ in the surface waters and sediments of the Nordic Seas: Implications for paleoceanography. Geochem. Geophys. Geosyst. 5, Q11013 (2004)
Prahl, F. G. et al. Assessment of sea-surface temperature at 42°N in the California Current over the last 30,000 years. Paleoceanography 10, 763–773 (1995)
Bernhard, J. M. Characteristic assemblages and morphologies of benthic foraminifera from anoxia, organic-rich deposits: Jurassic through Holocene. J. Foraminiferal Res. 16, 207–215 (1986)
Bernhard, J. & Reimers, C. Benthic foraminiferal population fluctuation related to anoxia: Santa Barbara Basin. Biochemistry 15, 127–149 (1991)
Kaiho, K. Benthic foraminiferal dissolved-oxygen index and dissolved-oxygen levels in the modern ocean. Geology 22, 719–722 (1994)
Jorissen, F. J., Fontanier, C. & Thomas, E. Paleoceanographical proxies based on deep-sea benthic foraminiferal assemblage characteristics. Proxies in Late Cenozoic Paleoceanography: Pt. 2: Biological tracers and biomarkers (eds Hillaire-Marcel C. & de Vernal, A. ) 263–326 (Elsevier, 2007)
Hermelin, J. O. R. & Shimmield, G. B. The importance of the oxygen minimum zone and sediment geochemistry on the distribution of recent benthic foraminifera from the NW Indian ocean. Mar. Geol. 91, 1–29 (1990)
Kienast, S. S., Calvert, S. E. & Pedersen, T. F. Nitrogen isotope and productivity variations along the northeast Pacific margin over the last 120 kyr: surface and subsurface paleoceanography. Paleoceanography 17, 7-1–7-17 (2002)
Thomson, R. E. & Krassovski, M. V. Poleward reach of the California Undercurrent extension. J. Geophys. Res. 115, C09027 (2010)
Robinson, R. S., Martinez, P., Pena, L. D. & Cacho, I. Nitrogen isotopic evidence for deglacial changes in nutrient supply in the eastern equatorial Pacific. Paleoceanography 24, PA4213 (2009)
Schmitt, J. et al. Carbon isotope constraints on the deglacial CO2 rise from ice cores. Science 336, 711–714 (2012)
Zhang, J., Quay, P. D. & Wilbur, D. O. Carbon isotope fractionation during gas water exchange and dissolution of CO2 . Geochim. Cosmochim. Acta 59, 107–114 (1995)
Fitzsimmons, J. N., Boyle, E. A. & Jenkins, W. J. Distal transport of dissolved hydrothermal iron in the deep South Pacific Ocean. Proc. Natl Acad. Sci. USA 111, 16654–16661 (2014)
Chang, A. S., Pichevin, L., Pedersen, T. F., Gray, V. & Ganeshram, R. New insights into productivity and redox-controlled trace element (Ag, Cd, Re, and Mo) accumulation in a 55 kyr long sediment record from Guaymas Basin, Gulf of California. Paleoceanography 30, 77–94 (2015)
Conley, D. J., Carstensen, J., Vaquer-Sunyer, R. & Duarte, C. M. Ecosystem thresholds with hypoxia. Hydrobiologia 629, 21–29 (2009)
Moffitt, S. E., Hill, T. M., Ohkushi, K., Kennett, J. P. & Behl, R. Vertical oxygen minimum zone oscillations since 20 ka in Santa Barbara Basin: A benthic foraminiferal community perspective. Paleoceanography 29, 44–57 (2014)
Schilt, A. et al. Atmospheric nitrous oxide during the last 140,000 years. Earth Planet. Sci. Lett. 300, 33–43 (2010)
Monnin, E. et al. Atmospheric CO2 concentrations over the last glacial termination. Science 291, 112–114 (2001)
Marcott, S. A. et al. Centennial-scale changes in the global carbon cycle during the last deglaciation. Nature 514, 616–619 (2014)
Hill, T. M. et al. Pre-Bølling warming in Santa Barbara Basin, California: surface and intermediate water records of early deglacial warmth. Quat. Sci. Rev. 25, 2835–2845 (2006)
Acknowledgements
We thank J. Padman for assistance with faunal counts, K. Brewster for assistance with alkenone sample preparation and analysis, and A. Guiheneuf for preliminary alkenone measurements and faunal assemblage data. This work was supported by NSF grants AGS-0602395 (Project PALEOVAR, A.C.M.) and OCE-1204204 (A.C.M. and F.G.P.), and an NSF graduate research fellowship for S.K.P.; J.A.A. was supported by the USGS Climate and Land Use Change Research and Development Program and the Volcano Science Center.
Author information
Authors and Affiliations
Contributions
S.K.P. and A.C.M. designed the study and wrote the paper. S.K.P., M.D.W., and F.G.P. contributed to alkenone palaeotemperature measurements and analysis. M.H.W. assisted with the chronology. J.A.A. provided insights on the trace metal and δ15N records. All authors contributed to interpretation of the data and provided comments on the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Extended data figures and tables
Extended Data Figure 1 Depth transect of oxygen isotopes from the Northeast Pacific.
Planktonic oxygen isotopes (Nps) from core EW0408-85JC (dark blue)2, benthic oxygen isotopes from core MD02-2503 in the Santa Barbara basin (Uvigerina peregrina; light blue, Bolivina argentea; bright blue)68, benthic Uvigerina peregrina oxygen isotopes from core EW0408-85JC (green)2, ODP Site 1019 ((41° 68′ N, 124° 93′ W, 978 m; light blue)1, cores EW0408-26JC/TC, and core EW0408-87JC (Cibicidoides). Data from the Gulf of Alaska cores (EW0408) are used to make the depth-time map shown in Fig. 4.
Extended Data Figure 2 Comparison of SST estimates based on the 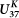 (purple) and
(purple) and 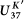 (black) indices.
(black) indices.
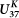 (purple) and
(purple) and 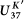 (black) indices.
(black) indices.Temperatures based on the  index show a larger glacial-interglacial change, with colder SSTs during the late glacial period and warmer SST for the Holocene. Temperature estimates for the deglacial period, including the two hypoxic intervals and the Younger Dryas, are virtually identical between methods, giving confidence that diagenetic effects are not influencing the alkenone ratios during the hypoxic warm events. Alkenone concentrations are high during the two hypoxic events (blue), consistent with other proxy evidence for high productivity and/or excellent preservation of organic matter during these events.
index show a larger glacial-interglacial change, with colder SSTs during the late glacial period and warmer SST for the Holocene. Temperature estimates for the deglacial period, including the two hypoxic intervals and the Younger Dryas, are virtually identical between methods, giving confidence that diagenetic effects are not influencing the alkenone ratios during the hypoxic warm events. Alkenone concentrations are high during the two hypoxic events (blue), consistent with other proxy evidence for high productivity and/or excellent preservation of organic matter during these events.
Extended Data Figure 3 Rate of SST change in the Gulf of Alaska.
The alkenone palaeotemperature record (purple) was interpolated on a 200-yr time step and the average rate of temperature change (°C per century) was calculated over a 400-yr window (black).
Extended Data Figure 4 Relative abundances of benthic species and genera in core EW0408-85JC.
Bulimina exilis is the most tolerant of low-oxygen conditions, and is often associated with near-anoxic bottom waters. Bulimina and Bolivina genera are typically found in strongly hypoxic waters, whereas Epistominella pacifica is associated with intermediate hypoxia, and Uvigerina peregrina is associated with more well-oxygenated conditions. Grey shaded bars represent the two laminated intervals, which are almost exclusively comprised of Bolivina and Bulimina genera.
Extended Data Figure 5 Records of surface and export productivity from EW0408-85JC.
The sedimentary δ15N record (violet) and δ15N corrected for terrestrial organic matter (light violet)13 show elevated values during the two hypoxic intervals (grey bars), which also coincide with enhanced organic matter deposition, including total organic carbon (light green)13 and an increase in the alkenone K37 abundance (dark blue). High planktonic δ13C values are observed during these intervals (bright blue)2, consistent with an increase in surface productivity rather than upwelling of deep waters exported from low-latitudes. The progressive increase in planktonic δ13C through the Holocene is accompanied by an increase in the relative abundance of U. peregrina (purple), likely indicating a better ventilated water column in the Holocene.
Extended Data Figure 6 Comparison of planktonic δ13C with sedimentary δ15N in core EW0408-85JC.
a, δ13C of planktonic foraminifera, Nps (blue), Gb (red), average of the two species (black). b, Comparison of the average planktonic foraminiferal δ13C (black) with changes in δ13C of atmospheric CO2 (blue)57 and estimated surface ocean δ13C of CO3=. (red, calculated from the smooth atmospheric values using the temperature relationship of Zhang et al.60 c, Relationship between δ15N in organic matter and δ13C in planktonic foraminifera (average of Gb and Nps). The δ15N and δ13C measurements were in most cases made in adjacent samples, typically separated by 1 cm. To prevent directional bias in the scatter plot, the two variables were first interpolated linearly onto the depth of the other variable. In no cases were interpolations allowed over an interval >5 cm or 200 years. The positive correlation between organic δ15N and planktonic foraminiferal δ13C during the deglacial interval (17–9 ka, red points, r2 = 0.51) supports an interpretation of increased nutrient utilization and carbon export from near-surface waters associated with the high δ15N events. In contrast, within Holocene time (9–0 ka) the relationship between δ15N and δ13C reverses, suggesting no systematic variations in nutrient utilization. d, Time series of organic δ15N (organic) and δ13C (average of the planktonic foraminifera Gb and Nps) as a time series; these data form the comparison in c.
Extended Data Figure 7 Expanded view of proxy data (plotted as depth in core) for EW0408-85JC during the two hypoxic intervals.
CT grey scale (black) reflects changes in the biogenic:lithogenic fraction of sediment, with low values indicating times of high biogenic input (primarily diatoms)2. The laminated intervals (pink shading) coincide with high diatom abundance and SSTs near or exceeding 10 °C, whereas evidence for low-oxygen conditions appears to extend both before (blue shading) and after (grey shading) the BA laminated zone based on trace metal concentrations12,13, benthic faunal assemblages, and preservation of TOC13, coinciding with the initial increase in SST and decrease in benthic δ18O (dark blue)2.
Extended Data Figure 8 Surface climate proxies compared with changes in benthic δ18O and fauna from different depth sites in the Gulf of Alaska.
The alkenone palaeotemperature record from core EW0408-85JC (purple), a composite record of planktonic δ18O from cores EW0408-26JC and EW0408-66JC (black)36, benthic δ18O (dark blue) and biogenic silica (bright blue) from core EW0408-85JC2, the combined abundance of low-oxygen tolerant Bolivina and Bulimina species from cores EW0408-85JC (682 m; green) and EW0408-26JC (1,620 m; violet). An increase in low-oxygen benthic fauna is apparent in the deeper site (EW0408-26JC) commencing at 16 ka, which coincides with the pre-Bølling warming in the SST record and an increase in the planktonic and benthic δ18O records. This initial decrease (blue shading) in sedimentary oxygen content at the base of the OMZ clearly precedes the large increase in biogenic silica and the shift to hypoxic conditions in core EW0408-85JC near the onset of sedimentary laminations (pink shading). Sediment laminations in core EW0408-26JC occur from 15–14 ka (shaded bar on x-axis).
Extended Data Figure 9 Northeast Pacific SSTs, productivity indices, and atmospheric greenhouse gases.
Data from top: CO2 record from EDC (dark blue)66 and WAIS (light blue)67, a record of N2O from TALOS Dome (bright blue)65, the Gulf of Alaska  SST record (purple), a
SST record (purple), a  SST record from the California margin (black)47, records of total organic carbon (TOC: light green), total nitrogen (TN: dark green), and δ15N records on bulk organic matter from core EW0408-85JC13. Grey shaded bars indicate the two intervals in which the deglacial rise in CO2 plateaus/reverses, which generally correspond to the episodes of widespread North Pacific hypoxia, high SSTs, enhanced nitrate utilization, and increased export productivity.
SST record from the California margin (black)47, records of total organic carbon (TOC: light green), total nitrogen (TN: dark green), and δ15N records on bulk organic matter from core EW0408-85JC13. Grey shaded bars indicate the two intervals in which the deglacial rise in CO2 plateaus/reverses, which generally correspond to the episodes of widespread North Pacific hypoxia, high SSTs, enhanced nitrate utilization, and increased export productivity.
Source data
Rights and permissions
About this article

Cite this article
Praetorius, S., Mix, A., Walczak, M. et al. North Pacific deglacial hypoxic events linked to abrupt ocean warming. Nature 527, 362–366 (2015). https://doi.org/10.1038/nature15753
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature15753
This article is cited by
-
Spatial and seasonal variations in coastal water physicochemical parameters in a southeastern Brazilian watershed
International Journal of Environmental Science and Technology (2024)
-
A global synthesis of high-resolution stable isotope data from benthic foraminifera of the last deglaciation
Scientific Data (2023)
-
Volcanic trigger of ocean deoxygenation during Cordilleran ice sheet retreat
Nature (2022)
-
A palaeothermometer of ancient Indigenous fisheries reveals increases in mean temperature of the catch over five millennia
Environmental Biology of Fishes (2022)
-
Enhanced North Pacific subtropical gyre circulation during the late Holocene
Nature Communications (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
