
Abstract
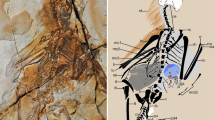
The wings of birds and their closest theropod relatives share a uniform fundamental architecture, with pinnate flight feathers as the key component1,2,3. Here we report a new scansoriopterygid theropod, Yi qi gen. et sp. nov., based on a new specimen from the Middle–Upper Jurassic period Tiaojishan Formation of Hebei Province, China4. Yi is nested phylogenetically among winged theropods but has large stiff filamentous feathers of an unusual type on both the forelimb and hindlimb. However, the filamentous feathers of Yi resemble pinnate feathers in bearing morphologically diverse melanosomes5. Most surprisingly, Yi has a long rod-like bone extending from each wrist, and patches of membranous tissue preserved between the rod-like bones and the manual digits. Analogous features are unknown in any dinosaur but occur in various flying and gliding tetrapods6,7,8,9,10, suggesting the intriguing possibility that Yi had membranous aerodynamic surfaces totally different from the archetypal feathered wings of birds and their closest relatives. Documentation of the unique forelimbs of Yi greatly increases the morphological disparity known to exist among dinosaurs, and highlights the extraordinary breadth and richness of the evolutionary experimentation that took place close to the origin of birds.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout



Similar content being viewed by others

References
Ji, Q., Currie, P. J., Norell, M. A. & Ji, S.-A. Two feathered dinosaurs from northeastern China. Nature 393, 753–761 (1998)
Xu, X. et al. Four-winged dinosaurs from China. Nature 421, 335–340 (2003)
Hu, D., Hou, L., Zhang, L. & Xu, X. A pre-Archaeopteryx troodontid from China with long feathers on the metatarsus. Nature 461, 640–643 (2009)
Xu, K. et al. Jurassic System in the North of China (VII): The Stratigraphic Region of Northeast China (Petroleum Industry Press, 2003)
Li, Q. et al. Melanosome evolution indicates a key physiological shift within feathered dinosaurs. Nature 507, 350–353 (2014)
Schutt, W. A., Jr & Simmons, N. B. Morphology and homology of the chiropteran calcar, with comments on the phylogenetic relationships of Archaeopteropus. J. Mamm. Evol. 5, 1–32 (1998)
Thorington, R. W., Jr & Darrow, K. A. C. J. Wing tip anatomy and aerodynamics in flying squirrels. J. Mamm. 79, 245–250 (1998)
Bennett, S. C. Articulation and function of the pteroid bone of pterosaurs. J. Vertebr. Paleontol. 27, 881–891 (2007)
Storch, G., Engesser, B. & Wuttke, M. Oldest fossil record of gliding in rodents. Nature 379, 439–441 (1996)
Simmons, N. B., Seymour, K. L., Habersetzer, J. & Gunnell, G. F. Primitive Early Eocene bat from Wyoming and the evolution of flight and echolocation. Nature 451, 818–821 (2008)
Sullivan, C. et al. The vertebrates of the Jurassic Daohugou Biota of northeastern China. J. Vertebr. Paleontol. 34, 243–280 (2014)
Irmis, R. B. Axial skeletal ontogeny in the Parasuchia (Archosauria: Pseudosuchia) and its implications for ontogenetic determination in Archosaurs. J. Vertebr. Paleontol. 27, 350–361 (2007)
Christiansen, P. & Fariña, R. A. Mass prediction in theropod dinosaurs. Hist. Biol. 16, 85–92 (2004)
Zhang, F., Zhou, Z., Xu, X., Wang, X. & Sullivan, C. A bizarre Jurassic maniraptoran from China with elongate ribbon-like feathers. Nature 455, 1105–1108 (2008)
O’Connor, J. & Sullivan, C. Reinterpretation of the Early Cretaceous maniraptoran (Dinosauria: Theropoda) Zhongornis haonae as a scansoriopterygid-like non-avian, and morphological resemblances between scansoriopterygids and basal oviraptorosaurs. Vertebr. Palasiat. 52, 3–30 (2014)
Zhang, F., Zhou, Z., Xu, X. & Wang, X. A juvenile coelurosaurian theropod from China indicates arboreal habits. Naturwissenschaften 89, 394–398 (2002)
Czerkas, S. A. & Yuan, C. X. in Feathered Dinosaurs and the Origin of Flight (ed. Czerkas, S. J.) 63–95 (The Dinosaur Museum, 2002)
Johnson-Murray, J. L. Myology of the gliding membranes of some Petauristine rodents (genera: Glaucomys, Pteromys, Petinomys, and Petaurista). J. Mamm. 58, 374–384 (1977)
Jackson, S. M. Glide angle in the genus Petaurus and a review of gliding mammals. Mammal Rev. 30, 9–30 (2000)
Feduccia, A. Bird origins anew. Auk 130, 1–12 (2013)
Moyer, A. E. et al. Melanosomes or microbes: testing an alternative hypothesis for the origin of microbodies in fossil feathers. Sci. Rep. 4, 4233 (2014)
Czerkas, S. A. & Feduccia, A. Jurassic archosaur is a non-dinosaurian bird. J. Ornithol. 155, 841–851 (2014)
Foth, C., Tischlinger, H. & Rauhut, O. W. M. New specimen of Archaeopteryx provides insights into the evolution of pennaceous feathers. Nature 511, 79–82 (2014)
Witmer, L. M. in Functional Morphology in Vertebrate Paleontology (ed. Thomason, J. J.) 19–33 (Cambridge Univ. Press, 1995)
Acknowledgements
We thank Z. Zhou, R. Dudley, J. Clarke and T. Stidham for discussion, X. Ding for specimen preparation, Y. Liu and Y. Han for providing illustrations, Z. Zhang for assistance with the EDS analysis, and Y. Hou for CT scanning the specimen. This research was funded by the National Natural Science Foundation of China (41372014, 41472023, 41120124002 and 41125008) and Major Basic Research Projects of the Ministry of Science and Technology, China (2012CB821900).
Author information
Authors and Affiliations
Contributions
X.X., X. Zheng and C.S. designed the project, X.X., X. Zheng, C.S., X.W., Y.W., L.X., X. Zhang, J.K.O’C., F.Z. and Y.P. performed the research, and X.X. and C.S. wrote the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Additional information
Data have been deposited in ZooBank under Life Science Identifier (LSID) http://zoobank.org/urn:lsid:zoobank.org:pub:5B765FD5-A534-45D5-AE14-62C25EADDD2D. The raw CT data have been deposited in the Dryad Digital Repository (http://dx.doi.org/10.5061/dryad.fp060).
Extended data figures and tables
Extended Data Figure 1 Fossil‐bearing beds of the Middle–Upper Jurassic Tiaojishan Formation in Qinglong County, Hebei.
The Mutoudeng Locality (top) and Gangou Locality (bottom) are geographically close to each other (approximately 20 km apart) and very similar in their sedimentology and fossil content. The bottom image shows an excavation we organized at the Gangou Locality.
Extended Data Figure 3 Close‐ups of the three counter slab fragments and corresponding slab regions of STM 31‐2.
Top row: isolated piece from counter slab preserving cranial region (left), and corresponding region of slab (right); middle row: isolated piece from counter slab preserving forelimb region (left), and corresponding region of slab (right); bottom row: isolated piece from counter slab preserving hindlimb region (left), and corresponding region of slab (right).
Extended Data Figure 4 Volumetric model generated from superimposed CT slices.
Because there is little density contrast between the fossil and the matrix, the outline of the fossil is somewhat vague.
Extended Data Figure 5 Photographs of preserved integumentary features of STM 31‐2.
a–cc, Feathers over the neck (a), along the humerus (b) and along the humerus and ulna (c); d, isolated basally converging feathers; e, sheet‐like soft tissue and feathers associated with the right forelimb and feathers along the right hindlimb; f, g, close‐ups of sheet‐like soft tissue associated with the right forelimb (f) and distal phalanges of manual digit IV (g).
Extended Data Figure 6 Elemental compositions of the styliform element and comparison samples based on EDS analyses.
EDS spectra derived from: a, right manual phalanx II‐1; b, right styliform element; c, feathers associated with right tibia; and d, sedimentary matrix.
Extended Data Figure 7 Aerodynamic apparatuses in Yi and other tetrapods.
a–cc, Wing of Yi, Bat Model (a), Maniraptoran Model (b), and Frog Model (c); d, bat wing; e, pigeon wing; f, pterosaur wing; g, giant Japanese flying squirrel wing. Yellow colour indicates styliform element in a, b, c, d, f, and g.
Extended Data Figure 8 Scanning electron microscopy sample locations.
Numbered circles indicate the locations of samples from the slab and counter slab. 1, distal part of cranial feathers; 2, middle part of tibial feathers; 3, distal part of humeral feathers; 4, distal part of ulnar feathers; 5, proximal part of cranial feathers; 6, proximal part of neck feathers; 7, distal part of humeral feathers; 8, distal part of humeral feathers; 9 and 10, membranous soft tissue near digit II; 11, membranous soft tissue near digit IV; 12, middle part of tibial feathers.
Extended Data Figure 9 Scanning electron microscopy images of melanosomes and melanosome impressions preserved in Yi qi holotype (STM 31‐2).
a–cm, Note the high morphological diversity of the preserved melanosomes in the feathers, which include: densely distributed, small round phaeomelanosomes and sparsely distributed, medium‐sized oval eumelanosomes in the feathers near the skull (a); medium‐sized elongate elliptical eumelanosomes in the feathers near the skull (b, c), neck (d, e), humerus (f, g), and ulna (h, i, j); and large oval and elliptical eumelanosomes in the feathers near the tibiotarsus (k, l). The subspherical phaeomelanosomes in the sheet‐like soft tissue (m) appear to be less densely distributed than the melanosomes in the feathers.
Related audio
41586_2015_BFnature14423_MOESM79_ESM.mp3
Kerri Smith talks to palaeontologist Xing Xu and editor Henry Gee about an unexpected find in the dinosaur lineage leading to birds.
Supplementary information
Supplementary Information
This file contains Supplementary Text and Data, Supplementary Tables 1-2 and Supplementary References. (PDF 621 kb)
CT data for holotype of Yi qi
This data archive contains CT slice images in .dcm format for the slab containing the holotype specimen (STM, Shandong Tianyu Museum, 31-2) of Yi qi, a scansoriopterygid dinosaur from the Middle-Upper Jurassic of Mutoudeng, Hebei Province, China. The CT data reveal the 3D structure of the specimen, and of the slab itself. (AVI 25715 kb)
Rights and permissions
About this article

Cite this article
Xu, X., Zheng, X., Sullivan, C. et al. A bizarre Jurassic maniraptoran theropod with preserved evidence of membranous wings. Nature 521, 70–73 (2015). https://doi.org/10.1038/nature14423
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature14423
This article is cited by
-
Origin of the propatagium in non-avian dinosaurs
Zoological Letters (2023)
-
Decoupling body shape and mass distribution in birds and their dinosaurian ancestors
Nature Communications (2023)
-
Sesamoids and Morphological Variation: a Hypothesis on the Origin of Rod-like Skeletal Elements in Aerial Mammals
Journal of Mammalian Evolution (2022)
-
‘Dinosaur-bird’ macroevolution, locomotor modules and the origins of flight
Journal of Iberian Geology (2021)
-
Archaeopteryx feather sheaths reveal sequential center-out flight-related molting strategy
Communications Biology (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
