
Abstract
Cancer-associated genetic alterations induce expression of tumour antigens that can activate CD8+ cytotoxic T cells (CTLs), but the microenvironment of established tumours promotes immune tolerance through poorly understood mechanisms1,2. Recently developed therapeutics that overcome tolerogenic mechanisms activate tumour-directed CTLs and are effective in some human cancers1. Immune mechanisms also affect treatment outcome, and certain chemotherapeutic drugs stimulate cancer-specific immune responses by inducing immunogenic cell death and other effector mechanisms3,4. Our previous studies revealed that B cells recruited by the chemokine CXCL13 into prostate cancer tumours promote the progression of castrate-resistant prostate cancer by producing lymphotoxin, which activates an IκB kinase α (IKKα)-BMI1 module in prostate cancer stem cells5,6. Because castrate-resistant prostate cancer is refractory to most therapies, we examined B cell involvement in the acquisition of chemotherapy resistance. Here we focus on oxaliplatin, an immunogenic chemotherapeutic agent3,4 that is effective in aggressive prostate cancer7. We show that mouse B cells modulate the response to low-dose oxaliplatin, which promotes tumour-directed CTL activation by inducing immunogenic cell death. Three different mouse prostate cancer models were refractory to oxaliplatin unless genetically or pharmacologically depleted of B cells. The crucial immunosuppressive B cells are plasmocytes that express IgA, interleukin (IL)-10 and programmed death ligand 1 (PD-L1), the appearance of which depends on TGFβ receptor signalling. Elimination of these cells, which also infiltrate human-therapy-resistant prostate cancer, allows CTL-dependent eradication of oxaliplatin-treated tumours.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout





Similar content being viewed by others
References
Chen, D. S. & Mellman, I. Oncology meets immunology: the cancer-immunity cycle. Immunity 39, 1–10 (2013)
Schietinger, A. & Greenberg, P. D. Tolerance and exhaustion: defining mechanisms of T cell dysfunction. Trends Immunol. 35, 51–60 (2014)
Zitvogel, L., Galluzzi, L., Smyth, M. J. & Kroemer, G. Mechanism of action of conventional and targeted anticancer therapies: reinstating immunosurveillance. Immunity 39, 74–88 (2013)
Kroemer, G., Galluzzi, L., Kepp, O. & Zitvogel, L. Immunogenic cell death in cancer therapy. Annu. Rev. Immunol. 31, 51–72 (2013)
Ammirante, M., Luo, J. L., Grivennikov, S., Nedospasov, S. & Karin, M. B-cell-derived lymphotoxin promotes castration-resistant prostate cancer. Nature 464, 302–305 (2010)
Ammirante, M. et al. An IKKα–E2F1–BMI1 cascade activated by infiltrating B cells controls prostate regeneration and tumor recurrence. Genes Dev. 27, 1435–1440 (2013)
Lee, J. L. et al. Gemcitabine-oxaliplatin plus prednisolone is active in patients with castration-resistant prostate cancer for whom docetaxel-based chemotherapy failed. Br. J. Cancer 110, 2472–2478 (2014)
Kaplan-Lefko, P. J. et al. Pathobiology of autochthonous prostate cancer in a pre-clinical transgenic mouse model. Prostate 55, 219–237 (2003)
Watson, P. A. et al. Context-dependent hormone-refractory progression revealed through characterization of a novel murine prostate cancer cell line. Cancer Res. 65, 11565–11571 (2005)
Ammirante, M., Shalapour, S., Kang, Y., Jamieson, C. A. & Karin, M. Tissue injury and hypoxia promote malignant progression of prostate cancer by inducing CXCL13 expression in tumor myofibroblasts. Proc. Natl Acad. Sci. USA 111, 14776–14781 (2014)
Cerutti, A. The regulation of IgA class switching. Nature Rev. Immunol. 8, 421–434 (2008)
Yoshizaki, A. et al. Regulatory B cells control T-cell autoimmunity through IL-21-dependent cognate interactions. Nature 491, 264–268 (2012)
Affara, N. I. et al. B cells regulate macrophage phenotype and response to chemotherapy in squamous carcinomas. Cancer Cell 25, 809–821 (2014)
Yu, Y. P. et al. Gene expression alterations in prostate cancer predicting tumor aggression and preceding development of malignancy. J. Clin. Oncol. 22, 2790–2799 (2004)
Luo, J. L. et al. Nuclear cytokine-activated IKKα controls prostate cancer metastasis by repressing Maspin. Nature 446, 690–694 (2007)
Doi, T. et al. IgA plasma cells express the negative regulatory co-stimulatory molecule programmed cell death 1 ligand and have a potential tolerogenic role in the intestine. Biochem. Biophys. Res. Commun. 425, 918–923 (2012)
Foster, B. A., Gingrich, J. R., Kwon, E. D., Madias, C. & Greenberg, N. M. Characterization of prostatic epithelial cell lines derived from transgenic adenocarcinoma of the mouse prostate (TRAMP) model. Cancer Res. 57, 3325–3330 (1997)
Hogquist, K. A. et al. T cell receptor antagonist peptides induce positive selection. Cell 76, 17–27 (1994)
Xiao, S. et al. Defect in regulatory B-cell function and development of systemic autoimmunity in T-cell Ig mucin 1 (Tim-1) mucin domain-mutant mice. Proc. Natl Acad. Sci. USA 109, 12105–12110 (2012)
Kang, H. S. et al. Signaling via LTβR on the lamina propria stromal cells of the gut is required for IgA production. Nature Immunol. 3, 576–582 (2002)
Feng, T., Elson, C. O. & Cong, Y. Treg cell-IgA axis in maintenance of host immune homeostasis with microbiota. Int. Immunopharmacol. 11, 589–592 (2011)
Shen, P. et al. IL-35-producing B cells are critical regulators of immunity during autoimmune and infectious diseases. Nature 507, 366–370 (2014)
Pardoll, D. M. The blockade of immune checkpoints in cancer immunotherapy. Nature Rev. Cancer 12, 252–264 (2012)
Qin, Z. et al. B cells inhibit induction of T cell-dependent tumor immunity. Nature Med. 4, 627–630 (1998)
Olkhanud, P. B. et al. Tumor-evoked regulatory B cells promote breast cancer metastasis by converting resting CD4+ T cells to T-regulatory cells. Cancer Res. 71, 3505–3515 (2011)
Fremd, C., Schuetz, F., Sohn, C., Beckhove, P. & Domschke, C. B cell-regulated immune responses in tumor models and cancer patients. OncoImmunology 2, e25443 (2013)
Shah, N. Diagnostic significance of levels of immunoglobulin A in seminal fluid of patients with prostatic disease. Urology 8, 270–272 (1976)
Schumacher, T. N., Kesmir, C. & van Buuren, M. M. Biomarkers in cancer immunotherapy. Cancer Cell 27, 12–14 (2015)
Shen, F. W. et al. Cloning of Ly-5 cDNA. Proc. Natl Acad. Sci. USA 82, 7360–7363 (1985)
Gingrich, J. R. et al. Metastatic prostate cancer in a transgenic mouse. Cancer Res. 56, 4096–4102 (1996)
Chen, J. et al. Immunoglobulin gene rearrangement in B cell deficient mice generated by targeted deletion of the JH locus. Int. Immunol. 5, 647–656 (1993)
Koh, D. R. et al. Less mortality but more relapses in experimental allergic encephalomyelitis in CD8−/− mice. Science 256, 1210–1213 (1992)
Mombaerts, P. et al. RAG-1-deficient mice have no mature B and T lymphocytes. Cell 68, 869–877 (1992)
Forrester, E. et al. Effect of conditional knockout of the type II TGF-β receptor gene in mammary epithelia on mammary gland development and polyomavirus middle T antigen induced tumor formation and metastasis. Cancer Res. 65, 2296–2302 (2005)
Harriman, G. R. et al. Targeted deletion of the IgA constant region in mice leads to IgA deficiency with alterations in expression of other Ig isotypes. J. Immunol. 162, 2521–2529 (1999)
Porichis, F. et al. High-throughput detection of miRNAs and gene-specific mRNA at the single-cell level by flow cytometry. Nature Commun. 5, 5641 (2014)
Michaud, M. et al. Autophagy-dependent anticancer immune responses induced by chemotherapeutic agents in mice. Science 334, 1573–1577 (2011)
Keren, Z. et al. B-cell depletion reactivates B lymphopoiesis in the BM and rejuvenates the B lineage in aging. Blood 117, 3104–3112 (2011)
Holzbeierlein, J. et al. Gene expression analysis of human prostate carcinoma during hormonal therapy identifies androgen-responsive genes and mechanisms of therapy resistance. Am. J. Pathol. 164, 217–227 (2004)
LaTulippe, E. et al. Comprehensive gene expression analysis of prostate cancer reveals distinct transcriptional programs associated with metastatic disease. Cancer Res. 62, 4499–4506 (2002)
Singh, D. et al. Gene expression correlates of clinical prostate cancer behavior. Cancer Cell 1, 203–209 (2002)
Arredouani, M. S. et al. Identification of the transcription factor single-minded homologue 2 as a potential biomarker and immunotherapy target in prostate cancer. Clin. Cancer Res. 15, 5794–5802 (2009)
Liu, P. et al. Sex-determining region Y box 4 is a transforming oncogene in human prostate cancer cells. Cancer Res. 66, 4011–4019 (2006)
Grasso, C. S. et al. The mutational landscape of lethal castration-resistant prostate cancer. Nature 487, 239–243 (2012)
Lapointe, J. et al. Gene expression profiling identifies clinically relevant subtypes of prostate cancer. Proc. Natl Acad. Sci. USA 101, 811–816 (2004)
Tomlins, S. A. et al. Integrative molecular concept modeling of prostate cancer progression. Nature Genet. 39, 41–51 (2007)
Welsh, J. B. et al. Analysis of gene expression identifies candidate markers and pharmacological targets in prostate cancer. Cancer Res. 61, 5974–5978 (2001)
Varambally, S. et al. Integrative genomic and proteomic analysis of prostate cancer reveals signatures of metastatic progression. Cancer Cell 8, 393–406 (2005)
Magee, J. A. et al. Expression profiling reveals hepsin overexpression in prostate cancer. Cancer Res. 61, 5692–5696 (2001)
Wallace, T. A. et al. Tumor immunobiological differences in prostate cancer between African-American and European-American men. Cancer Res. 68, 927–936 (2008)
Vanaja, D. K., Cheville, J. C., Iturria, S. J. & Young, C. Y. Transcriptional silencing of zinc finger protein 185 identified by expression profiling is associated with prostate cancer progression. Cancer Res. 63, 3877–3882 (2003)
Luo, J. H. et al. Gene expression analysis of prostate cancers. Mol. Carcinog. 33, 25–35 (2002)
Rhodes, D. R. et al. ONCOMINE: a cancer microarray database and integrated data-mining platform. Neoplasia 6, 1–6 (2004)
Di Caro, G. et al. Occurrence of tertiary lymphoid tissue is associated with T-cell infiltration and predicts better prognosis in early-stage colorectal cancers. Clin. Cancer Res. 20, 2147–2158 (2014)
Woo, J. R. et al. Tumor infiltrating B-cells are increased in prostate cancer tissue. J. Transl. Med. 12, 30 (2014)
D'Amico, A. V. et al. Biochemical outcome after radical prostatectomy, external beam radiation therapy, or interstitial radiation therapy for clinically localized prostate cancer. J. Am. Med. Assoc. 280, 969–974 (1998)
Czéh, M. et al. The immune response to sporadic colorectal cancer in a novel mouse model. Oncogene 29, 6591–6602 (2010)
Acknowledgements
We thank L. Bastian, K. Wang, A. Umemura, M. K. Kim, M. Susani, E. Gurnhofer and F. Grizzi for discussions and research materials. Antibodies and MACS-beads were gifts from eBioscience, Biolegends and Miltenyi Biotec. Anti-PD-L1 and Pdl1/2−/− mice were from I. Mellman (Genentech). Research was supported by the National Institutes of Health (NIH) (CA127923 and AI043477), DFG and German Cancer Consortium (TR36, DKTK to G.W.), the Genome Research-Austria project ‘Inflammobiota’ (FWF and P26011 to L.K.), the CureSearch Foundation (to D.D.) and postdoctoral research fellowships from the German Research Foundation (DFG, SH721/1-1 to S.S.); Irvington-CRI (to S.S. and Z.Z.); CIRM (TG2-01154 to J.F.-B.) and FIRC/AIRC (to G.D.C.). M.K. is an ACS Research Professor and holds the Ben and Wanda Hildyard Chair for Mitochondrial and Metabolic Diseases.
Author information
Authors and Affiliations
Contributions
M.K. and S.S. conceived and designed the project. S.S. performed experiments. S.S. and M.K. analysed data. J.F.B., Z.Z., D.D., M.A., G.W. and A.S. assisted with experiments and analysis. S.S., G.D.C., E.S.-L. and D.E.H. performed immunohistochemical analyses of human samples. G.W. performed Tag-specific ELISA. D.E.H., C.J., P.B., C.J.K., T.K. and L.K. collected and provided human specimens. M.K. and S.S. wrote the manuscript, with all authors contributing to writing and providing feedback.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Extended data figures and tables
Extended Data Figure 1 Treatment schemes and characterization of tumours and mouse survival before and after treatment.
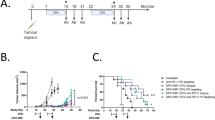
a, Early and late treatment schemes for TRAMP mice. b, TRAMP mice (n = 3–6 per group) were subjected to early oxaliplatin treatment as described in a and prostate weights were determined at 14 weeks, 1 week after completion of four treatment cycles. Dashed red line indicates prostate weight of tumour-free controls (n = 33 in total). Prostate weight in b is shown in a log2 scale. c, d, Histopathology of TRAMP tumours. c, Representative images of haematoxylin-and-eosin-stained prostate sections from TRAMP mice are shown. Scale bars, 100 μm. PD, poorly differentiated adenocarcinoma; PIN, prostatic intraepithelial neoplasia; WD, well differentiated adenocarcinoma. d, Histopathological assessment of early and late treated TRAMP tumours in wild-type and Jh−/− mice without or with oxaliplatin treatment. The percentages of the different histotypes shown in c are depicted (n = 3–7 per group). Fisher’s chi-square analysis was used to calculate statistical significance. e, Early and late treatment schemes for mice bearing subcutaneous (s.c.) MC tumours. f, MC cells were subcutaneously transplanted into wild-type and Jh−/− mice (n = 3–7 per group) that were subjected to early oxaliplatin treatment when tumour volume was 100 mm3. Forty-eight hours after completion of three treatment cycles, mice were euthanized and tumour volumes (mm3) were measured (n = 19 in total). g, MC tumours from indicated mice were stained for CD45 (green) and cleaved caspase 3 (CC3; red) (n = 4–6 per group). h, i, MC tumours (n = 3–5 per group) grown in wild-type, Jh−/− and Cd8a−/− mice were stained for CD45 (green) and γH2AX (red), and the γH2AX+ foci in CD45− cells were enumerated (i). Scale bars, 100 μm. All results are mean ± s.e.m. j, Representative images of subcutaneous MC tumours (n = 5–6 per group) from wild-type and Jh−/− mice, with or without oxaliplatin treatment stained for αSMA (red) and CD31 (green). k, l, Frequency of αSMA- (k) and CD31- (l) positive cells within tumours from j. Shown are median values ± s.e.m. *P < 0.05; **P < 0.01; ***P < 0.001 (Mann–Whitney and t-tests). m, TRAMP mice (wild-type, Cd8a−/− or Jh−/−; n = 6–14 per group) were treated weekly with low-dose oxaliplatin. Moribund mice were euthanized, and survival was compared by Kaplan–Meyer analysis and significance was determined (wild-type: P > 0.05; Cd8a−/−: P > 0.05; Jh−/−: **P < 0.002). n, Survival curves for the different TRAMP groups before and after oxaliplatin treatment. Significant differences in survival times are indicated on the right. No statistically significant differences in survival were found between wild-type and Jh−/− or Jh−/− and Cd8a−/− mice without treatment. Significant differences in survival times were observed between all three oxaliplatin-treated groups (wild-type, Cd8a−/− or Jh−/−; indicated on the right). *P < 0.05; **P < 0.01; ***P < 0.001 (Kaplan–Meyer analysis). The number of mice used in each experiment and the number of experiments (replications) are shown also in Supplementary Table 1.
Extended Data Figure 2 B cells attenuate oxaliplatin-triggered CTL activation.
a, Flow cytometry of CD8+ T lymphocytes in prostates of 20-week-old TRAMP mice after four cycles of oxaliplatin treatment (n = 4–6 per group) normalized to prostate weights. b, Late subcutaneous MC tumours from wild-type and Jh−/− mice were stained for CD8 and analysed by immunofluorescent microscopy. In the top left areas (white square), single CD8 staining (green) without DAPI counterstain is shown. Tumoral CD8+ cells were counted in 3–4 HMF (200×) per tumour (n = 4–5 tumours per group). Scale bars, 100 μm. c, d, Late subcutaneous MC tumours were analysed by flow cytometry for CD4+ lymphocytes in spleens (c) and tumours (d) after three oxaliplatin treatment cycles (n = 4–7 per group). The results show percentages of CD4+ cells in the CD45+ population. e, Flow cytometric analysis of TNF and IFNγ expression by CD8+ cells in MC tumours from wild-type and Jh−/− mice treated as above (n = 6–8) and re-stimulated in vitro with tumour cell lysate. f, Flow cytometry of STAT1 phosphorylation in CD8+ cells from MC tumours of treated and untreated wild-type and Jh−/− mice (for isotype controls, see Extended Data Fig. 10u). The results are summarized in the right panel (n = 3 mice per group). g, Expression of GrzB and Ki67 in CD8+ T effector cells (CD8+CD44+) from spleens of MC inoculated mice after oxaliplatin treatment. h, i, Flow cytometry of PD-1 and Tim-3 expression by CD8+ T effector cells (CD8+CD44+ cells) in spleen (h) and MC tumours (i) as indicated with or without oxaliplatin treatment. Shown are percentages of the corresponding CD8+ T cells in the CD8+CD44+ population (n = 3–5 per group). j, The experimental scheme for B cell immunodepletion in tumour-bearing mice. MC tumours were raised in wild-type or Cd8a−/− mice, 16 days after subcutaneous tumour cell inoculation. B cells were depleted by twice weekly administration of antibodies directed against CD19, CD20, CD22 and B220. Four days after first antibody treatment, mice were treated with oxaliplatin (n = 4–7 mice per group, total: 44). After three weekly chemotherapy cycles, mice were euthanized. k, Flow cytometry analysis of tumour-infiltrating CD45+CD8+ T cells stained for IFNγ (left) or IFNγ and TNF (right) after in vitro restimulation with PMA and ionomycin (n = 4–6 mice per group). l–n, Flow cytometry analysis of CD19+ (l, m) and IgA+ (n) cells in spleens and tumours isolated from the wild-type mice described above, confirming depletion of CD19+ B cells and oxaliplatin-induced IgA+ cells in spleen and tumours. o, Serum IgA concentrations in the mice described in j (n = 3– 5 per group). p, Flow cytometry analyses of CD19+ B cells in tumours isolated from Cd8a−/− mice subjected to B cell depletion or not, confirmed the efficient depletion of tumoral CD19+ B cells. All results are mean ± s.e.m. *P < 0.05; **P < 0.01; ***P < 0.001 (Mann–Whitney and t-tests).
Extended Data Figure 3 Immunogenic chemotherapy induces tumour infiltration by immunosuppressive CD19+CD20lowB220lowIgA+ B cells.
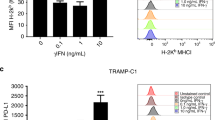
a, b, MC tumours (n = 4–9 per group) raised in wild-type mice without or with oxaliplatin treatment were stained for B220 (a), and tumour-infiltrating B220+ cells per HMF were enumerated (b). In a, single B220 staining (above) and combined staining B220/DAPI (below) are shown. Scale bars, 100 μm. c, d, The flow cytometry plots and gating strategy for analysis of splenic B cell populations using CD19, IgA, B220, CD138 and CD20 antibodies. Results from wild-type mice bearing MC tumours are shown in c and from oxaliplatin-treated mice in d (n = 8 mice per group). Oxaliplatin treatment modestly increased the amount of splenic IgA+ cells. Splenic IgA+ cells expressed CD138 as expected and showed lower levels of B220 and CD20, in either control or oxaliplatin-treated mice. e, The gating strategies for analysis of tumoral B cells using CD19, IgA, B220 and CD138 antibodies. Results from MC tumours in two representative oxaliplatin-treated wild-type mice are shown (n = 8 mice per group), demonstrating the presence of IgA+ cells in oxaliplatin-treated tumours with a typical CD138+B220low phenotype. f–i, Flow cytometry plots and gating strategies for analysis of tumoral B cell populations using CD19, B220, CD138, IgA and PD-L1 antibodies. Results from wild-type mice bearing MC tumours without (f) or with (g) oxaliplatin treatment (n = 6 mice per group) and Iga−/− mice bearing MC tumours without (h) or with (i) oxaliplatin treatment (n = 6 mice per group) are shown. Oxaliplatin treatment increased the amount of tumoral IgA+CD138+B220lowPD-L1+cells in wild-type mice. j, Flow cytometric analysis of PD-L1 and IL-10 expression in IgA+B220−/low and B220+IgA− B cells from oxaliplatin-treated TRAMP tumours (n = 4). k, Flow cytometric analysis of IgA and CD138 expression by TRAMP tumour-infiltrating B cells. Shown are percentages of IgA+ cells among all tumour-infiltrating CD19+CD138+ cells. l, Wild-type mice bearing MC tumours were treated with oxaliplatin as above. Two days after the first or last oxaliplatin cycle, mice were euthanized, tumours were isolated and analysed by flow cytometry as indicated (n = 6 per group). After dead-cell exclusion, tumour-infiltrating B cells were stained with CD19, CD20, B220, IgA and IgM antibodies. Shown are the results for control (left), one cycle (middle), and three cycles (right) of oxaliplatin treatment, demonstrating the presence of tumoral IgA+ cells with a CD19+CD20lowB220low IgA+ cell phenotype within 48 h after oxaliplatin treatment.
Extended Data Figure 4 Immunogenic chemotherapy induces tumoral and systemic IgA production through CSR.
a, Ex vivo analysis of IgA released by tumour single cell suspension isolated from oxaliplatin-treated TRAMP tumours. Single cell suspension from non-treated tumours and culture medium without cells were used as controls. b, c, Serum IgA (b) and IgG (c) in treated and untreated TRAMP mice and age-matched naive FVB controls (n = 7–14 per group). d, Serum IgA amounts in control or oxaliplatin-treated mice bearing MC tumours (n = 5–7 per group) were determined and compared to age-matched naive FVB controls (n = 7). e, Serum IgG amounts in control or oxaliplatin-treated mice bearing MC tumours (n = 5–7 mice per group) were determined and compared to age-matched naive FVB controls. a–e, Results are mean ± s.e.m. Mann–Whitney and t-tests were used to calculate statistical significance. f, g, Immunofluorescence analysis of activation-induced cytidine deaminase (AID, green) and IgA (red) expression in spleen (f, used as a positive control) and MC tumours from oxaliplatin-treated wild-type mice (g). Scale bars: 10 μm (f, left and g) and 100 μm (f, right). Arrows point to IgA+AID+ cells. Shown are representative results of spleens and tumours isolated from four mice per group. h, qRT–PCR analysis of Tgfb1 mRNA in MC tumours raised in wild-type or Jh−/− mice without or with oxaliplatin treatment (n = 3–7 mice per group). Results are mean ± s.e.m. i, Flow cytometry of SMAD2/3 phosphorylation in MC tumour-infiltrating B cells from wild-type mice before and after oxaliplatin treatment (n = 4 per group). Shown are the MFI and percentages (see Fig. 3e). j, Flow cytometry of SMAD2/3 phosphorylation and PD-L1 in MC tumour-infiltrating B cells from wild-type mice before and after oxaliplatin treatment (n = 4 per group). Shown are the percentages of PD-L1+p-SMAD2/3+ cells within CD45+CD19+ cells. k, qRT–PCR analysis of Il21 mRNA in MC tumours raised in wild-type or Jh−/− mice without or with oxaliplatin treatment (n = 4–5 mice per group). Chemotherapy-induced Il21 mRNA mainly in wild-type mice. l, m, Flow cytometry of tumour-infiltrating B cells stained for p-STAT3 and IL-10 (n = 5–6 per group) before and after oxaliplatin treatment. n, Flow cytometry analysis of Actb (β-actin) mRNA, IL-10 protein and Il10 mRNA in MC tumour-infiltrating IgA+ cells using PrimeFlow RNA technology (pooled data of 4 mice per group, after oxaliplatin treatment). Left, Actb mRNA gated on CD45+ cells; middle, Il10 mRNA and IL-10 protein expression after 1 h stimulation with PMA plus ionomycin and LPS gated on IgA+ cells, right panel: Il10 mRNA and IL-10 protein expression after 5 h stimulation with PMA plus ionomycin and LPS, gated on IgA+ cells. o, p, Flow cytometric analysis of tumour-infiltrating B cells in TRAMP mice (n = 4–5 per group) stained for CD19, B220, IL-10 and LTαβ (o). The percentage of tumour-infiltrating LTαβ+ cells amongst all tumour-infiltrating B cells was determined (p). q, Flow cytometric analyses of CD5 expression by B cells from spleen and MC-tumour of wild-type mice after oxaliplatin treatment (n = 4–5 per group). Shown are mean ± s.e.m. *P < 0.05; **P < 0.01; ***P < 0.001 (Mann–Whitney and t-tests).
Extended Data Figure 5 Immunogenic chemotherapy or B cell deficiency has marginal effects on Treg, natural killer and myeloid cells.
a, b, qRT–PCR analyses of Nos2 (a) and Arg1(b) mRNA content of MC tumours (n = 4–7 mice per group). Chemotherapy induced Nos2 and Arg1 expression in wild-type and Jh−/− mice and no significant and consistent differences were found between both groups. c, d, qRT–PCR analyses of Il12p40 (c), Il12p35 (d) mRNA in MC tumours grown in wild-type and Jh−/− mice (n = 4–6 mice per group). e–i, Flow cytometry analyses of tumour-infiltrating or splenic lymphocytes and monocytes: tumoral Nk1.1+ cells (e), tumoral CD11b+CD11c+MHCII+ cells (f), tumoral CD11b+GR-1+ cells (g), CD4+FoxP3+ cells (splenic, h; tumoral, i). Cells in e–i are from tumour-bearing mice subjected to oxaliplatin treatment and/or B cell depletion as indicated (B cell depletion plus oxaliplatin; n = 4–6 mice per group). Results are mean ± s.e.m. Mann–Whitney and t-tests were used to calculate statistical significance.
Extended Data Figure 6 Analyses of B and T cells in human prostate cancer specimens.
a–g, Tissue microarrays of tumour and non-tumour tissue from 110 PC patients were stained for CD8 and CD20 (5–6 spots per patient = 3–4 tumour tissue and 2 non-tumour tissue). a, b, Representative examples of CD8 (a) and CD20 (b) IHC of PC tissue microarrays (left). Right, computer assisted image analysis with ad hoc developed image software. Tumour tissue is represented in yellow and CD8+ and CD20+ cells are represented in red. The percentages of immune reactive area (IRA) occupied by CD8+ or CD20+ cells are shown. Scale bars, 200 μm. c, d, Comparison of CD8 and CD20 IRAs in matched non-tumour and tumour tissues from each E-PC patient (n = 87). e–g, Patients were divided into three subgroups: E-PC (n = 86), TR-PC (n = 9) and M-PC (n = 15). e, CD8+ cell infiltration into tumour tissues of the different groups. f, CD20+ cell infiltration into tumour tissues of the different groups. g, The CD8+/CD20+ ratio for the different groups. Each dot represents one patient. Line indicates the median value. Mann–Whitney test was used to calculate statistical significance between the two groups. Kruskal–Wallis test was used to calculate statistical significance between the three groups. h, IHC analysis of low-risk (n = 5) and high-risk (n = 5) human PC specimens using IgA (red) and αSMA (black). Nuclei were counterstained with haematoxylin. Scale bar, 100 μm i, Immunofluorescence analysis of human PC showing IL-10-expressing (red) IgA+ (green) CD138+ (turquoise) plasma cells (n = 6). Representative images are shown. White arrow indicates IL-10-expressing IgA+ cells. Scale bars, 50 μm. j, Human normal prostate (n = 3–5) and human PC (n = 5) were stained for IgA and CD8. Typical images are shown. Red and green arrows indicate IgA+ and CD8+ cells, respectively. Scale bar, 100 μm. k, Human normal prostate (n = 3) and human PC (n = 5) were stained for IgA (red arrow) and CD20 (green arrow). Scale bar, 100 μm. l, Flow cytometric analysis of human prostate tumour-infiltrating CD19+ cells and IgA+ cells. The percentages of IL-10-expressing B cells in CD19+IgA+ (two different samples) and CD19+IgA− B cells are shown. m, Summary of results obtained from human blood samples taken from healthy donors (n = 3) and patients with PC (n = 5) and prostate tissue specimens (benign, malignant; n = 4) analysed by flow cytometry for IL-10 expression in CD19+IgA− and CD19+IgA+ B cells. n, o, Tissue microarrays from 110 patients with PC (described above) were stained for IgA and CD138. Patients were divided into three subgroups: E-PC (n = 86), TR-PC (n = 9) and M-PC (n = 15). n, Representative images of IgA (immunoperoxidase) and CD138 (alkaline phosphatase) double staining of tumour tissues from each group. CD138+ and IgA+ double-positive cells in the PC stroma are indicated by the white arrows (haematoxylin counterstain). Scale bar, 100 μm. o, Frequencies of IgA+ and CD138+ double-positive cells in the tumour stroma of the different PC patient groups. p, PC patient specimens were divided into two groups: IgA−/low (n = 64) and IgA+/hi (n = 46). Shown is the CD8+/CD20+ ratio for each group. Each dot represents one patient. Line indicates the median value. q, IgA mRNA expression (IGHA1) is significantly increased in human PC tissue relative to healthy or benign prostate tissue in 5 out of 15 studies evaluated via Oncomine. Results from one significant study14 are presented. *P < 0.05; **P < 0.01; ***P < 0.001 (chi-square test and Fisher’s exact test).
Extended Data Figure 7 Effects of TGFβR2, IgA, PD-L1 and IL-10 ablations on tumour-infiltrating lymphocytes.
MC tumours were raised in wild-type, Tgfbr2ΔB or Iga−/− mice (n = 5–11 per group). Mice were subjected to three cycles of late oxaliplatin treatment after which splenic and tumoral B cells were analysed. After dead cell exclusion, splenic (Spl; a, b) and tumoral (Tu; c, d) B cells were stained with CD19, B220, IgA, IgG2a and IgG1 antibodies and analysed by flow cytometry. e, Serum IgG concentrations in control or oxaliplatin-treated wild-type, Tgfbr2ΔB or Iga−/− mice bearing MC tumours (n = 5–9 per group). f, Flow cytometry of tumour-infiltrating CD19+ B cells from wild-type, Tgfbr2ΔBor Iga−/− MC tumour-bearing mice (n = 4–7 per group) analysed for PD-L1 expression, revealing lower PD-L1 surface expression on Tgfbr2Δ and Iga−/− B cells after oxaliplatin treatment. g, Flow cytometry of tumour-infiltrating B220hi B cells (left) and IgA+B220low B cells (right) from wild-type, Tgfbr2ΔBor Iga−/− MC tumour-bearing mice (n = 4–7 per group) analysed for IL-10 expression, revealing no difference in IL-10 expression by B220hiIgA− B cells in the corresponding groups, and lower IL-10 expression by Tgfbr2Δ B cells after oxaliplatin treatment compared to wild-type mice. Results are mean ± s.e.m. Mann–Whitney and t-tests were used to calculate statistical significance. h, The experimental scheme. Wild-type mice bearing MC tumours were divided into four treatment groups (n = 7–8 per group): (1) isotype control (IgG2a), (2) oxaliplatin (weekly), (3) anti-PD-L1 (twice weekly), and (4) oxaliplatin plus anti-PD-L1 (weekly and twice weekly, respectively). After three treatment cycles, mice were euthanized and analysed. i, Tumour growth curves of tumour-bearing mice and gross appearance of untreated and treated mice. Significance was determined by Mann–Whitney and t-tests. j, Flow cytometric analysis for GrzB expression by tumour-infiltrating CD8+ T effector cells (CD8+CD44+) from MC tumour-bearing mice treated as described above. Results are shown either as percentages of GrzB+ cells amongst CD8+ T cells (black), or percentages of GrzB+CD8+CD44+ T cells amongst tumoral CD45+ cells (red). k, Flow cytometry of PD-L1 expression on tumour-infiltrating IgA+ CD19+ B cells in the different treatment groups. l, m, Serum IgA (l) and IgG (m) concentrations in the different treatment groups described in h. n, The experimental scheme for the experiment whose results are shown in Fig. 4g. B cells were isolated from wild-type, Pdl1/2−/− and Il10−/− mice and 5 × 106 cells (purity 98%) were intraperitoneally transferred into MC tumour-bearing Jh−/− mice (16 days after MC cell inoculation). After 2 days (day 18), the mice were given three oxaliplatin treatment cycles and analysed. o, Flow cytometric analysis of splenocytes after staining with CD45 and CD19 antibodies, confirming presence of B cells in the ABCT groups. Shown are percentages and absolute B cell numbers in spleen. p, Tumour infiltrating CD8+ cells from MC tumour-bearing Jh−/− mice transplanted with B cells and treated as above were re-stimulated for 4 h with PMA and ionomycin before flow cytometry (n = 4–6 mice per group). Results are mean ± s.e.m. *P < 0.05; **P < 0.01; ***P < 0.001 (Mann–Whitney and t-tests).
Extended Data Figure 8 Low-dose cisplatin treatment is devoid of immunogenic activity and low-dose oxaliplatin does not affect gut barrier function.
a, Flow cytometry of MC cells stained with annexin V and propidium iodide 24 h after treatment with either oxaliplatin or cisplatin (both at 20 μM). b, Flow cytometry analysis of MC cells treated as above and stained with antibody to the autophagy marker LC3A. c–e, MC tumours were raised in wild-type and Jh−/− mice until 400 mm3 in size, after which the mice were treated with either cisplatin or oxaliplatin at 6 mg kg−1 (n = 4–5 per group). After three weekly chemotherapy cycles, mice were euthanized. c, Tumour weights; left: wild-type mice; right: Jh−/− mice. d, e, Flow cytometry of tumour-infiltrating CD8 (d) and CD4 (e) cells. Left: wild-type mice, right: Jh−/− mice. f, Gut permeability was measured in wild-type mice before and after low- (LD) and high- (HD) dose oxaliplatin treatment using orally administered fluorescein isothiocyanate (FITC)-dextran. Shown are FITC-dextran concentrations in serum (μg ml−1) (n = 5 mice per group). g, Serum IgA concentrations in naive wild-type (FVB) and Tgfbr2ΔB mice before and after oxaliplatin treatment (n = 3–6 per group). h, IgA staining of colon sections of untreated or low-dose oxaliplatin-treated wild-type mice. Scale bars, 100 μm. i–k, Flow cytometry of CD8+ (i), CD4+ (j) and Nk1.1+ (k) cells in spleens of naive wild-type and Tgfbr2Δ mice without or with oxaliplatin treatment. All results are mean ± s.e.m. *P < 0.05; **P < 0.01; ***P < 0.001 (Mann–Whitney and t-tests).
Extended Data Figure 9 Immunogenic chemotherapy supports adoptive T cell transfer only in the absence of B cells.
a, The experimental scheme. Immunogenic TRAMP-C2 cells were subcutaneously inoculated into wild-type or Tcrb−/− mice. After 30 days, the mice were divided into four groups (n = 4–5 per group): (1) control, (2) oxaliplatin (weekly), (3) ATCT, and (4) ATCT plus oxaliplatin (weekly). The first oxaliplatin cycle was given at day 31. Two days after the second cycle, CD8+ T cells from CD45.1 × CD45.2 wild-type mice (3 × 106 cells) were transferred into tumour-bearing mice and this was followed by two more oxaliplatin cycles after which mice were euthanized for analysis on day 59. b, Tumour volumes (mm3) c, d, Flow cytometric analysis of spleen (c) and tumour (d) cells after staining with CD45.1, CD45.2, CD8 and TCRαβ antibodies, confirming expansion of adoptively transferred T cells. e, Tumour growth curves. f, The experimental scheme. Immunogenic TRAMP-C2 cells were subcutaneously inoculated into wild-type or Rag1−/−;OT-1 mice (no B cells), that contain CD8+ T cells specific for chicken ovalbumin which is not expressed by TRAMP-C2 cells. After 30 days, tumour-bearing Rag1−/−;OT-1 mice were divided into four groups (n = 3–4 mice per group): (1) control, (2) oxaliplatin treatment, (3) ATCT, and (4) oxaliplatin treatment plus ATCT. The first oxaliplatin cycle was given at day 31. Two days after the second oxaliplatin cycle, CD8+ T cells (3 × 106) from CD45.1 × CD45.2 mice were adoptively transferred into tumour-bearing mice, which were euthanized on day 59 and analysed. g, Flow cytometric analysis of tumour-infiltrating cells stained with CD45.1, CD45.2, CD8 and TCRαβ antibodies, confirming infiltration of adoptively transferred T cells. h, Flow cytometric analysis of GrzB expression in adoptively transferred, tumour-infiltrating, CD8+ T effector cells (CD45.1+CD8+CD44+) from tumour-bearing mice treated as above. i, Tumour volumes (mm3). j, Tumour growth curves. k, The experimental scheme for Fig. 5a–f. Sixteen-week-old TRAMP;Rag1−/− mice (no B and T cells) were treated with oxaliplatin (weekly). One day after the first treatment cycle, CFSE-labelled splenocytes from either wild-type (B and T cells, SP-WT) or Jh−/− (T but no B cells, SP-Jh−/−) mice were transferred into the tumour-bearing mice (5 × 106 T cells per mouse; 4–5 mice per group). l, m, After 6 days, one mouse from each group was euthanized , and the proliferation of CD8+ (l) and CD4+ (m) cells in bone marrow (BM), spleen and prostates was analysed by CFSE staining and flow cytometry. n–r, After three more oxaliplatin cycles (4 weeks in total), the mice were euthanized and analysed. n, Frequency of adoptively transferred CD19+ cells amongst CD45+ cells in spleens and prostates 30 days after ACT. o, Flow cytometric analyses of CD19+ B lymphocytes for TIM-1 expression in spleens (left) and prostates (right) of above mice. p–r, Flow cytometric analyses of T cells. Percentages of CD8+ and CD4+ cells in lymph nodes (LN; p); spleens (q); prostates (r) of above TRAMP;Rag1−/− mice. Red: splenocytes from wild-type mice (T and B cell transfer), blue: splenocytes from Jh−/− mice (T cell transfer). Results are mean ± s.e.m. *P < 0.05; **P < 0.01; ***P < 0.001 (Mann–Whitney and t-tests).
Extended Data Figure 10 Immunogenic chemotherapy supports adoptive T cell transfer only in the absence of B cells and analysis of lymphocytes and monocytes in tumour-free mice.
a, The experimental scheme for Fig. 5g–i. MC tumour-bearing Rag1−/− mice (no B and T cells) were treated with oxaliplatin (weekly). One day after first oxaliplatin treatment, 5 × 106 T cells (negative selection) from wild-type mice immunized with MC cell lysate37 were adoptively transferred into tumour-bearing mice (4–5 mice per group), alone or in combination with 5 × 106 B cells from wild-type or Tgfbr2ΔB mice (purity 98%). After two more oxaliplatin cycles (3 weeks total), the mice were euthanized and analysed. b, Serum IgG analysis of above mice. c, Flow cytometric analysis of splenocytes after staining with CD45 and CD19 antibodies. d–p, Wild-type, Jh−/−, Iga−/− and Tgfbr2ΔB mice in the FVB background and wild-type, Pdl1/2−/−, Il10−/− and Iga−/− in the C57BL/6 background were analysed for the distribution of immune markers. d, Spleen weights of wild-type, Jh−/− and Tgfbr2ΔB mice in the FVB background. e, Flow cytometry of splenocytes for the following markers: CD3 (left), CD8 (middle) and CD4 (right), gated on the splenic CD45+ population. f, Absolute cell numbers of splenic CD3+ (left), CD8+ (middle) and CD4+ (right) cells are shown (percentage × cell count of whole spleen). g, h, Flow cytometry for TNF and IFNγ in CD8+ cells from tumour-free wild-type, Jh−/−, Tgfbr2ΔB and Iga−/− mice (n = 6–8) that were re-stimulated in vitro with PMA and ionomycin and the representative flow cytometry panels (h). i, j, Flow cytometry of splenocytes from wild-type and Tgfbr2ΔB for: CD19+IgM+ cells (i) and IgA (j) gated on the splenic CD45+ population. k–n, Flow cytometry of splenocytes from wild-type, Pdl1/2−/−, Il10−/− and Iga−/− mice for: CD45+CD19+IgM+ cells (k), CD45+IgA+ cells (l), PD-L1 expression by CD19+IgM+ cells (m), and IL-10 expression by CD19+ cells (n), as indicated. o, p, Serum IgA and IgG concentrations were analysed in wild-type, Pdl1/2−/− and Il10−/− mice (n = 4–5 mice per group). q–u, The different gating strategies and staining controls are shown. q, Gating strategies for tumour-infiltrating lymphocytes: lymphocyte gate, dead cell exclusion, doublets exclusion, and gating on the CD45+ population. r, Flow cytometric analysis of IL-10 and IgA expression, gated on the CD45+ population: (1) isotype control (no staining); (2) non-stimulated splenocytes: showing IgA staining, but not IL-10; (3) stimulated splenocytes from Il10−/− mice showing IgA staining, but not IL-10; and (4) stimulated splenocytes from wild-type mice showing IgA and IL-10 staining. s, Flow cytometric analysis of IL-10 and CD19 expression, gated on the CD19+B220+ population. Left, stimulated cells from Il10−/− mice, showing B cell staining, but not IL-10; right: stimulated cells from wild-type mice showing B cell staining and IL-10 staining. t, Flow cytometric analysis of IL-10 and IgA expression, gated on the IgA+ population: left: stimulated cells from Il10−/− mice, showing IgA cell staining, but not IL-10. Right, stimulated cells from wild-type mice showing IgA and IL-10 staining. These results confirm IL-10 production by IgA+ cells. u, Flow cytometric analysis of p-STAT1 staining with corresponding isotype control. All results are mean ± s.e.m. *P < 0.05; **P < 0.01; ***P < 0.001 (Mann–Whitney and t-tests).
Supplementary information
Supplementary Information
This file contains Supplementary Tables 1-2. (PDF 211 kb)
Rights and permissions
About this article

Cite this article
Shalapour, S., Font-Burgada, J., Di Caro, G. et al. Immunosuppressive plasma cells impede T-cell-dependent immunogenic chemotherapy. Nature 521, 94–98 (2015). https://doi.org/10.1038/nature14395
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature14395
This article is cited by
-
B cells in head and neck squamous cell carcinoma: current opinion and novel therapy
Cancer Cell International (2024)
-
The role of regulatory B cells in immune regulation and childhood allergic asthma
Molecular and Cellular Pediatrics (2024)
-
Innate and adaptive immune cell interaction drives inflammasome activation and hepatocyte apoptosis in murine liver injury from immune checkpoint inhibitors
Cell Death & Disease (2024)
-
PPARδ inhibition blocks the induction and function of tumor-induced IL-10+ regulatory B cells and enhances cancer immunotherapy
Cell Discovery (2023)
-
Immuno-regulatory malignant B cells contribute to Chronic Lymphocytic Leukemia progression
Cancer Gene Therapy (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
