
Abstract
Cell growth and proliferation are tightly linked to nutrient availability. The mechanistic target of rapamycin complex 1 (mTORC1) integrates the presence of growth factors, energy levels, glucose and amino acids to modulate metabolic status and cellular responses1,2,3. mTORC1 is activated at the surface of lysosomes by the RAG GTPases and the Ragulator complex through a not fully understood mechanism monitoring amino acid availability in the lysosomal lumen and involving the vacuolar H+-ATPase4,5,6,7,8. Here we describe the uncharacterized human member 9 of the solute carrier family 38 (SLC38A9) as a lysosomal membrane-resident protein competent in amino acid transport. Extensive functional proteomic analysis established SLC38A9 as an integral part of the Ragulator–RAG GTPases machinery. Gain of SLC38A9 function rendered cells resistant to amino acid withdrawal, whereas loss of SLC38A9 expression impaired amino-acid-induced mTORC1 activation. Thus SLC38A9 is a physical and functional component of the amino acid sensing machinery that controls the activation of mTOR.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others
Accession codes
Primary accessions
ArrayExpress
Data deposits
The protein-protein interactions have been submitted to the IMEx (http://www.imexconsortium.org) consortium through IntAct (http://www.ebi.ac.uk/intact/) and assigned the identifier IM-23283. The SLC network has the IntAct accession number EBI-9975668 and the RAGA-RAGC-LAMTOR network is EBI-9975664. RNA-Seq data is available in ArrayExpress (http://www.ebi.ac.uk/arrayexpress) under the accession number E-MTAB-3102.
References
Dibble, C. C. & Manning, B. D. Signal integration by mTORC1 coordinates nutrient input with biosynthetic output. Nature Cell Biol. 15, 555–564 (2013)
Laplante, M. & Sabatini, D. M. mTOR signaling in growth control and disease. Cell 149, 274–293 (2012)
Cornu, M., Albert, V. & Hall, M. N. mTOR in aging, metabolism, and cancer. Curr. Opin. Genet. Dev. 23, 53–62 (2013)
Kim, E., Goraksha-Hicks, P., Li, L., Neufeld, T. P. & Guan, K. L. Regulation of TORC1 by Rag GTPases in nutrient response. Nature Cell Biol. 10, 935–945 (2008)
Sancak, Y. et al. The Rag GTPases bind raptor and mediate amino acid signaling to mTORC1. Science 320, 1496–1501 (2008)
Sancak, Y. et al. Ragulator-Rag complex targets mTORC1 to the lysosomal surface and is necessary for its activation by amino acids. Cell 141, 290–303 (2010)
Zoncu, R. et al. mTORC1 senses lysosomal amino acids through an inside-out mechanism that requires the vacuolar H+-ATPase. Science 334, 678–683 (2011)
Bar-Peled, L., Schweitzer, L. D., Zoncu, R. & Sabatini, D. M. Ragulator is a GEF for the rag GTPases that signal amino acid levels to mTORC1. Cell 150, 1196–1208 (2012)
Jewell, J. L., Russell, R. C. & Guan, K. L. Amino acid signalling upstream of mTOR. Nature Rev. Mol. Cell Biol. 14, 133–139 (2013)
Bar-Peled, L. et al. A tumor suppressor complex with GAP activity for the Rag GTPases that signal amino acid sufficiency to mTORC1. Science 340, 1100–1106 (2013)
Tsun, Z. Y. et al. The folliculin tumor suppressor is a GAP for the RagC/D GTPases that signal amino acid levels to mTORC1. Mol. Cell 52, 495–505 (2013)
Han, J. M. et al. Leucyl-tRNA synthetase is an intracellular leucine sensor for the mTORC1-signaling pathway. Cell 149, 410–424 (2012)
Panchaud, N., Peli-Gulli, M. P. & De Virgilio, C. Amino acid deprivation inhibits TORC1 through a GTPase-activating protein complex for the Rag family GTPase Gtr1. Sci. Signal. 6, ra42 (2013)
Nicklin, P. et al. Bidirectional transport of amino acids regulates mTOR and autophagy. Cell 136, 521–534 (2009)
Hediger, M. A., Clemencon, B., Burrier, R. E. & Bruford, E. A. The ABCs of membrane transporters in health and disease (SLC series): introduction. Mol. Aspects Med. 34, 95–107 (2013)
Uhlen, M. et al. Towards a knowledge-based Human Protein Atlas. Nature Biotechnol. 28, 1248–1250 (2010)
Chapel, A. et al. An extended proteome map of the lysosomal membrane reveals novel potential transporters. Mol. Cell. Proteomics 12, 1572–1588 (2013)
Schiöth, H. B., Roshanbin, S., Hagglund, M. G. & Fredriksson, R. Evolutionary origin of amino acid transporter families SLC32, SLC36 and SLC38 and physiological, pathological and therapeutic aspects. Mol. Aspects Med. 34, 571–585 (2013)
Ögmundsdóttir, M. H. et al. Proton-assisted amino acid transporter PAT1 complexes with Rag GTPases and activates TORC1 on late endosomal and lysosomal membranes. PLoS ONE 7, e36616 (2012)
Hägglund, M. G. et al. Identification of SLC38A7 (SNAT7) protein as a glutamine transporter expressed in neurons. J. Biol. Chem. 286, 20500–20511 (2011)
Duran, R. V. et al. Glutaminolysis activates Rag-mTORC1 signaling. Mol. Cell 47, 349–358 (2012)
Zhang, Z., Gameiro, A. & Grewer, C. Highly conserved asparagine 82 controls the interaction of Na+ with the sodium-coupled neutral amino acid transporter SNAT2. J. Biol. Chem. 283, 12284–12292 (2008)
Oppedisano, F., Pochini, L., Broer, S. & Indiveri, C. The B degrees AT1 amino acid transporter from rat kidney reconstituted in liposomes: kinetics and inactivation by methylmercury. Biochim. Biophys. Acta 1808, 2551–2558 (2011)
Ljungdahl, P. O. Amino-acid-induced signalling via the SPS-sensing pathway in yeast. Biochem. Soc. Trans. 37, 242–247 (2009)
Goberdhan, D. C., Meredith, D., Boyd, C. A. & Wilson, C. PAT-related amino acid transporters regulate growth via a novel mechanism that does not require bulk transport of amino acids. Development 132, 2365–2375 (2005)
Settembre, C. et al. A lysosome-to-nucleus signalling mechanism senses and regulates the lysosome via mTOR and TFEB. EMBO J. 31, 1095–1108 (2012)
Giacomini, K. M. et al. Membrane transporters in drug development. Nature Rev. Drug Discov. 9, 215–236 (2010)
Wu, B. et al. Competitive intra- and extracellular nutrient sensing by the transporter homologue Ssy1p. J. Cell Biol. 173, 327–331 (2006)
Kriel, J., Haesendonckx, S., Rubio-Texeira, M., Van Zeebroeck, G. & Thevelein, J. M. From transporter to transceptor: signaling from transporters provokes re-evaluation of complex trafficking and regulatory controls. BioEssays 33, 870–879 (2011)
Taylor, P. M. Amino acid transporters: eminences grises of nutrient signalling mechanisms? Biochem. Soc. Trans. 37, 237–241 (2009)
Varjosalo, M. et al. Interlaboratory reproducibility of large-scale human protein-complex analysis by standardized AP-MS. Nature Methods 10, 307–314 (2013)
Giambruno, R. et al. Affinity purification strategies for proteomic analysis of transcription factor complexes. J. Proteome Res. 12, 4018–4027 (2013)
Pichlmair, A. et al. Viral immune modulators perturb the human molecular network by common and unique strategies. Nature 487, 486–490 (2012)
Colinge, J., Masselot, A., Giron, M., Dessingy, T. & Magnin, J. OLAV: towards high-throughput tandem mass spectrometry data identification. Proteomics 3, 1454–1463 (2003)
Bennett, K. L. et al. Proteomic analysis of human cataract aqueous humour: Comparison of one-dimensional gel LCMS with two-dimensional LCMS of unlabelled and iTRAQ(R)-labelled specimens. J. Proteomics 74, 151–166 (2011)
Choi, H. et al. SAINT: probabilistic scoring of affinity purification-mass spectrometry data. Nature Methods 8, 70–73 (2011)
Mellacheruvu, D. et al. The CRAPome: a contaminant repository for affinity purification-mass spectrometry data. Nature Methods 10, 730–736 (2013)
Snijder, B. et al. Population context determines cell-to-cell variability in endocytosis and virus infection. Nature 461, 520–523 (2009)
Galluccio, M. et al. Over-expression in E. coli and purification of the human OCTN1 transport protein. Protein Expr. Purif. 68, 215–220 (2009)
Pochini, L., Scalise, M., Galluccio, M., Amelio, L. & Indiveri, C. Reconstitution in liposomes of the functionally active human OCTN1 (SLC22A4) transporter overexpressed in Escherichia coli. Biochem. J. 439, 227–233 (2011)
Acknowledgements
We thank D. M. Sabatini, S. Wang and Z. Tsun for discussing results before publication and generously providing Flag–SLC38A9 and Flag–METAP2 stably expressing cells, all members of the Superti-Furga laboratory for discussions, the Bennett laboratory for the proteomic analyses, F. Pauler and the Barlow laboratory for the RNA-seq analysis and M. Gstaiger for providing expression vectors. This work was supported by the Austrian Academy of Sciences, ERC grant to G.S.-F. (i-FIVE 250179), EMBO long-term and Marie Curie fellowships to M.R. (ALTF 1346-2011, IEF 301663), EMBO long-term fellowship to R.K.K. (ALTF 314-2012), Swiss NSF fellowship (P300P3_147897) to B.S., Vienna Science and Technology Fund (WWTF VRG10-001) and the Austrian Science Fund (FWF P 25522-B20) to C.K., the Italian Ministry of Instruction University and Research, PON-ricerca e competitività 2007-2013 (no. PON01_00937) to C.I., the Austrian Federal Ministry for Science and Research (GenAu projects, APP-III and BIN-III) to L.A.H., K.L.B. and G.S.-F., the Austrian Science Fund MCBO/SFB021 to L.A.H.
Author information
Authors and Affiliations
Contributions
M.R. and G.S.-F. conceived the study. L.P., M.G. and C.I. designed and performed transport assays. M.R., T.S., M.E.G.d.A., E.L.R., M.B., K.L.B., L.A.H. and G.S.-F. designed and performed TAP-mass spectrometry experiments. M.R., M.E.G.d.A., B.S., A.F., M.B., S.S. and P.A.F. performed the other experiments. M.R., L.A.H. and G.S.-F. designed the other experiments. R.K.K. and B.S. performed bioinformatic data and image analysis. K.V.M.H., J.W.B., L.X.H., C.K. generated reagents and provided scientific insight. M.R. and G.S.-F. wrote the manuscript. All authors contributed to the discussion of results and participated in manuscript preparation.
Corresponding author
Ethics declarations
Competing interests
A patent has been filed with data generated in this manuscript where M.R. and G.S.-F. are listed as inventors.
Extended data figures and tables
Extended Data Figure 1 Expression of SLC members of amino acid transporter families.
a, Table of SLCs belonging to amino acid transporter families robustly expressed in HEK293 and K562 cells as monitored by RNA-seq. SLC members of amino acid transporter-containing families16 (SLC1, 6, 7, 16, 17, 18, 32, 36, 38 and 43 families) expressed (FPKM >0.5) in both cell lines were ranked according to their expression level, top ten are shown. The number of PubMed entries was obtained by querying the GeneSymbol (24 October 2013). b, Expression of members of the SLC32, SLC36 and SLC38 families in HEK293 and K562 cells.
Extended Data Figure 2 Biochemical and functional characterization of SLC38A9.
a, b, Where indicated, HEK293T cells were transfected with tagged SLC38A9 constructs (+) or empty vector (−). Cell lysates were left untreated (Untr.) or incubated 1 h at 37 °C in presence or absence of PNGase and analysed by immunoblot. Results are representative of two independent experiments (n = 2). c, Cell size measurements of HEK293T cells after short hairpin (shRNA)-mediated knockdown against GFP (control, dashed black line) or SLC38A9 (grey line), measured by automated microscopy and image analysis. Sparse and interphase cells were selected using image analysis and machine learning, and nucleus diameter was used as robust proxy for cell size38. Smoothed distributions of 2,400 and 4,165 cells, respectively, are shown. d, Cell proliferation measurement of HEK293T cells transduced with lentivirus-encoded shRNA against SLC38A9 or GFP. 105 cells were seeded and counted every 24 h. Mean values ± s.d. from triplicates. Results are representative of two independent experiments (n = 2). e, f, Where indicated, HEK293T cells were transfected with tagged SLC38A9. Cell lysates were prepared and left untreated (Untr.) or incubated 1 h at 37 °C with PNGase and analysed by immunoblot, Where indicated, cell lysates were boiled for 5 min at 95 °C after PNGase treatment. g, h, Lysates from murine NIH/3T3 (g) or Raw 264.7 (h) cells were subjected to immunoprecipitation with the indicated antibodies, treated with PNGase and analysed by immunoblot. Results are representative of two independent experiments (n = 2). <, SLC38A9; *, non-specific band.
Extended Data Figure 3 SLC38A9 proteomic analysis: bait localization and results.
a, Single-channel and merged confocal microscopy images of DAPI stained nuclei and indirect immunofluorescence against HA-tagged SLC38A9 and endogenous lysosomal markers LAMP1 (top panel) and LAMP2 (middle panel) and the non-induced and secondary antibody only control (bottom panel) in HEK293 Flp-In TREx cells. Scale bar, 10 μm. Intensity profiles for SLC38A9 (green) and LAMP1, LAMP2 or secondary antibody control (red) along the cross-section lines indicated in the respective merged channel images are shown. b, Quantification of HA-SLC38A9 signal above background (dashed lines in a) that colocalizes with LAMP1, LAMP2 or secondary antibody only positive areas. Average and s.d. of at least two images is shown, analysing colocalization in 22, 34 and 27 cells respectively. c, HEK293 Flp-In TREx cells inducibly expressing SLC38A9 were treated or not with doxycycline (Dox) for 24 h. Where indicated, cell lysates were treated with PNGase and analysed by immunoblot. d, Tabular view summarizing the proteomic analysis of SLC38A9, SLC38A1, SLC38A2 and SLC36A1. Comparison of the SLC38A9 interactors identified by TAP–LC–MS/MS to the same analysis performed with the other transporters. Spectral counts (Sp. c., average of biological replicates) and sequence coverage (Sq. c., percentage, average of biological replicates) are indicated. Data shown are based on two independent experiments for each condition (n = 2), each analysed in two technical replicates.
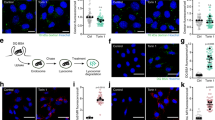
Extended Data Figure 4 SLC38A9 localizes to the late endosome/lysosome compartment.
a–h, HeLa cells were transfected with the indicated ST-HA tagged SLC38A9 construct. Merged and single-channel confocal microscopy images of indirect immunofluorescence of HA-tagged SLC38A9 (red) and endogenous lysosomal marker LAMP1 (green) are shown. Representative cells are shown. Scale bar, 10 μm.
Extended Data Figure 5 SLC38A9 is an integral component of the Ragulator–RAG GTPase complex.
a, Tabular view of spectral counts (Spec. count, average of biological replicates) and sequence coverage (Seq. cov., percentage, average of biological replicates) of the core Ragulator–RAG GTPase network and published interactors detected. Data shown are based on two independent experiments for each condition (n = 2), and analysed in two technical replicates. b, c, SLC38A9 peptides detected in LAMTOR1, 3, 4 and 5 (b) or in SLC38A9 (c) TAP–LC–MS/MS analysis are mapped on SLC38A9 sequence and highlighted in bold. Transmembrane helices are highlighted in light brown. Potential tryptic cleavage sites are in red.
Extended Data Figure 6 The cytoplasmic N-terminal region of SLC38A9 binds the Ragulator–RAG GTPase complex through evolutionary conserved motifs.
a, Sequence alignment of the N-terminal cytoplasmic region (amino acids 1–112) of human, mouse, rat, Xenopus and zebrafish SLC38A9. Amino acids selected for deletion and motifs substituted to alanine are highlighted. Black and grey shading indicates >60% amino acid sequence identity and similarity, respectively. b, c, HEK293T cells were transfected with the indicated tagged SLC38A9 constructs. Immunoprecipitates and cell extracts were analysed by immunoblot. SLC38A9 mutant constructs are labelled with the number of the encoded amino acids (b) or with the amino acid motif substituted to alanine (c). Results are representative of two independent experiments (n = 2).
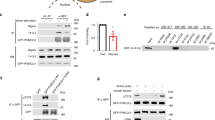
Extended Data Figure 7 Characterization of SLC38A9-mediated amino acid transport in proteoliposomes.
a, Purification of SLC38A9. Lanes represent empty vector control and SLC38A9 expressed in E. coli and purified by Ni-chelating chromatography. Immunoblot of the same fractions using anti-His or anti-SLC38A9 antibody are shown. b, Orientation of SLC38A9 in proteoliposomes. Purified His-SLC38A9 protein or proteoliposomes reconstituted with SLC38A9 were incubated overnight at 37 °C in presence or in absence of 1 U thrombin. Proteoliposomes were then solubilized with SDS and analysed by immunoblot. Results are representative of two independent experiments (n = 2). c, Time course of glutamine uptake by SLC38A9 in proteoliposomes reconstituted with the purified protein fraction. The uptake of 10 μM [3H]glutamine was measured at different time intervals in the presence of the indicated intraliposomal sodium concentrations. Transport was calculated by subtracting the radioactivity associated to proteoliposomes reconstituted with the empty vector fraction. Values represent means of specific transport ± s.d. from three independent experiments (n = 3). d, Time course of glutamine uptake in proteoliposomes reconstituted with purified SLC38A9 wild-type or N128A mutant protein. Values represent means of specific transport ± s.d. from 3 independent experiments (n = 3). Significance was estimated by Student’s t-test (*P < 0.01). Immunoblot analysis of purified protein reconstituted in the proteoliposomes. e, Effect of pH on the reconstituted SLC38A9. Reconstitution and transport assay were performed at the indicated pH. Results are means of specific transport rate ± s.d. from three different experiments (n = 3). f, Inhibition of the [3H]glutamine uptake in proteoliposomes. 1 mM MeAIB (α-(methylamino)isobutyric acid) was added together with 10 μM [3H]glutamine. Transport was measured at 60 min. Values represent means of percent residual activity with respect to control (without added inhibitor) ± s.d. from three independent experiments (n = 3).
Extended Data Figure 8 Nucleotide-loading/conformation dependent interaction of RAGB/RAGC heterodimers with SLC38A9.
HEK293T cells were transfected with the indicated combination of tagged RAG GTPases mutant constructs or empty vector (−). Anti-HA immunoprecipitates and cell extracts were treated with PNGase and analysed by immunoblot. W, wild type; 75, S75N; 120, Q120L; 99, Q99L; 54, T54N. Results are representative of two independent experiments (n = 2).
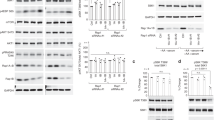
Extended Data Figure 9 Stable expression of SLC38A9 mediates sustained mTORC1 activation upon amino acid starvation.
a, SLC38A9 or METAP2 stably expressing HEK293T cells were starved for the indicated time in medium without amino acids and serum. Cell lysates were analysed by immunoblot. Results are representative of two independent experiments (n = 2). b, Representative images in the GFP channels of HEK293T cells stably expressing EGFP–LC3B and SLC38A9 or METAP2 starved for 120 min (related to Fig. 4b). Scale bar, 40 μm. c, HEK293T cells stably expressing TFEB–STHA and SLC38A9 or METAP2 were starved for the indicated time. Cytoplasmic and nuclear fraction were analysed by immunoblot. Results are representative of two independent experiments (n = 2). d, Immunoblot analysis of HEK293T cells stably expressing the indicated SLC38A9 constructs. e, SLC38A9 (S) or METAP2 (M) stably expressing HEK293T were starved for 50 min and then stimulated with amino acids for 20 min. Where indicated, cells were treated with concanamycin A (5 µM) or DMSO during both incubation times. Cell lysates were analysed by immunoblot with the indicated antibodies. Results are representative of two independent experiments (n = 2). f, SLC38A9 (S) or METAP2 (M) stably expressing HEK293T were treated for 30 min with DMSO (D), concanamycin A (C, 5 µM) or Torin 1 (T, 250 nM) and then starved for the indicated times in presence of the inhibitors. Cell lysates were analysed by immunoblot. Results are representative of two independent experiments (n = 2).
Extended Data Figure 10 Expression of SLC38A9 is required for amino acid-induced mTORC1 activation and is not affected by starvation.
a, HeLa cells were transfected with siRNA targeting SLC38A9 (SLC), LAMTOR1 (LT1) or non-targeting control (Cntr). After 72h, cells were starved for 50 min in medium without amino acids and serum and then stimulated with amino acids in presence of insulin (1 µM). Cell lysates were analysed by immunoblot. Results are representative of three independent experiments (n = 3). b, c, HEK293T cells were starved for the indicated times. SLC38A9 expression was analysed by quantitative PCR (b) and immunoblot (c). In b, mean values ± s.d. from technical triplicates are shown. Results are representative of two independent experiments (n = 2). <, SLC38A9; *, non-specific band.
Rights and permissions
About this article

Cite this article
Rebsamen, M., Pochini, L., Stasyk, T. et al. SLC38A9 is a component of the lysosomal amino acid sensing machinery that controls mTORC1. Nature 519, 477–481 (2015). https://doi.org/10.1038/nature14107
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature14107
This article is cited by
-
Glutamine addiction in tumor cell: oncogene regulation and clinical treatment
Cell Communication and Signaling (2024)
-
Lysosomes as coordinators of cellular catabolism, metabolic signalling and organ physiology
Nature Reviews Molecular Cell Biology (2024)
-
Mobilization of cholesterol induces the transition from quiescence to growth in Caenorhabditis elegans through steroid hormone and mTOR signaling
Communications Biology (2024)
-
Bacterial over-production of the functionally active human SLC38A2 (SNAT2) exploiting the mistic tag: a cheap and fast tool for testing ligands
Molecular Biology Reports (2024)
-
Amino acid metabolism in immune cells: essential regulators of the effector functions, and promising opportunities to enhance cancer immunotherapy
Journal of Hematology & Oncology (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
