
Abstract
Cytotoxic chemotherapy is effective in debulking tumour masses initially; however, in some patients tumours become progressively unresponsive after multiple treatment cycles. Previous studies have demonstrated that cancer stem cells (CSCs) are selectively enriched after chemotherapy through enhanced survival1,2,3. Here we reveal a new mechanism by which bladder CSCs actively contribute to therapeutic resistance via an unexpected proliferative response to repopulate residual tumours between chemotherapy cycles, using human bladder cancer xenografts. Further analyses demonstrate the recruitment of a quiescent label-retaining pool of CSCs into cell division in response to chemotherapy-induced damages, similar to mobilization of normal stem cells during wound repair4,5,6,7. While chemotherapy effectively induces apoptosis, associated prostaglandin E2 (PGE2) release paradoxically promotes neighbouring CSC repopulation. This repopulation can be abrogated by a PGE2-neutralizing antibody and celecoxib drug-mediated blockade of PGE2 signalling. In vivo administration of the cyclooxygenase-2 (COX2) inhibitor celecoxib effectively abolishes a PGE2- and COX2-mediated wound response gene signature, and attenuates progressive manifestation of chemoresistance in xenograft tumours, including primary xenografts derived from a patient who was resistant to chemotherapy. Collectively, these findings uncover a new underlying mechanism that models the progressive development of clinical chemoresistance, and implicate an adjunctive therapy to enhance chemotherapeutic response of bladder urothelial carcinomas by abrogating early tumour repopulation.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others

Accession codes
Primary accessions
BioProject
Data deposits
RNA-seq raw data have been deposited in the NCBI BioProject database under accession number PRJNA263046.
References
Visvader, J. E. & Lindeman, G. J. Cancer stem cells in solid tumours: accumulating evidence and unresolved questions. Nature Rev. Cancer 8, 755–768 (2008)
Clevers, H. The cancer stem cell: premises, promises and challenges. Nature Med. 17, 313–319 (2011)
Kreso, A. & Dick, J. E. Evolution of the cancer stem cell model. Cell Stem Cell 14, 275–291 (2014)
Beachy, P. A., Karhadkar, S. S. & Berman, D. M. Tissue repair and stem cell renewal in carcinogenesis. Nature 432, 324–331 (2004)
Arwert, E. N., Hoste, E. & Watt, F. M. Epithelial stem cells, wound healing and cancer. Nature Rev. Cancer 12, 170–180 (2012)
Ito, M. et al. Stem cells in the hair follicle bulge contribute to wound repair but not to homeostasis of the epidermis. Nature Med. 11, 1351–1354 (2005)
Mascré, G. et al. Distinct contribution of stem and progenitor cells to epidermal maintenance. Nature 489, 257–262 (2012)
Kim, J. J. & Tannock, I. F. Repopulation of cancer cells during therapy: an important cause of treatment failure. Nature Rev. Cancer 5, 516–525 (2005)
Wilson, A. et al. Hematopoietic stem cells reversibly switch from dormancy to self-renewal during homeostasis and repair. Cell 135, 1118–1129 (2008)
Chan, K. S. et al. Identification, molecular characterization, clinical prognosis, and therapeutic targeting of human bladder tumor-initiating cells. Proc. Natl Acad. Sci. USA 106, 14016–14021 (2009)
Volkmer, J. P. et al. Three differentiation states risk-stratify bladder cancer into distinct subtypes. Proc. Natl Acad. Sci. USA 109, 2078–2083 (2012)
Ho, P. L., Lay, E. J., Jian, W., Parra, D. & Chan, K. S. Stat3 activation in urothelial stem cells leads to direct progression to invasive bladder cancer. Cancer Res. 72, 3135–3142 (2012)
Ho, P. L., Kurtova, A. & Chan, K. S. Normal and neoplastic urothelial stem cells: getting to the root of the problem. Nature reviews . Urology 9, 583–594 (2012)
Shin, K. et al. Cellular origin of bladder neoplasia and tissue dynamics of its progression to invasive carcinoma. Nature Cell Biol. 16, 469–478 (2014)
Van Batavia, J. et al. Bladder cancers arise from distinct urothelial sub-populations. Nature Cell Biol. 16, 982–991 (2014)
Wang, X., Zinkel, S., Polonsky, K. & Fuchs, E. Transgenic studies with a keratin promoter-driven growth hormone transgene: prospects for gene therapy. Proc. Natl Acad. Sci. USA 94, 219–226 (1997)
Kelland, L. The resurgence of platinum-based cancer chemotherapy. Nature Rev. Cancer 7, 573–584 (2007)
Cotsarelis, G., Sun, T. T. & Lavker, R. M. Label-retaining cells reside in the bulge area of pilosebaceous unit: implications for follicular stem cells, hair cycle, and skin carcinogenesis. Cell 61, 1329–1337 (1990)
Chen, J. et al. A restricted cell population propagates glioblastoma growth after chemotherapy. Nature 488, 522–526 (2012)
Green, D. R. The end and after: how dying cells impact the living organism. Immunity 35, 441–444 (2011)
Goessling, W. et al. Genetic interaction of PGE2 and Wnt signaling regulates developmental specification of stem cells and regeneration. Cell 136, 1136–1147 (2009)
Hoggatt, J. et al. Differential stem- and progenitor-cell trafficking by prostaglandin E2. Nature 495, 365–369 (2013)
Lukacs, R. U., Goldstein, A. S., Lawson, D. A., Cheng, D. & Witte, O. N. Isolation, cultivation and characterization of adult murine prostate stem cells. Nature Protocols 5, 702–713 (2010)
Meeks, J. J. et al. A systematic review of neoadjuvant and adjuvant chemotherapy for muscle-invasive bladder cancer. Eur. Urol. 62, 523–533 (2012)
Sternberg, C. N. et al. ICUD-EAU International Consultation on Bladder Cancer 2012: chemotherapy for urothelial carcinoma-neoadjuvant and adjuvant settings. Eur. Urol. 63, 58–66 (2013)
Niegisch, G. et al. Neoadjuvant chemotherapy in patients with muscle-invasive bladder cancer: which patients benefit? Eur. Urol. 64, 355–357 (2013)
Coxib and traditional NSAID Trialists’ (CNT) Collaboration. Vascular and upper gastrointestinal effects of non-steroidal anti-inflammatory drugs: meta-analyses of individual participant data from randomised trials. Lancet 382, 769–779 (2013)
Wu, X. R. Urothelial tumorigenesis: a tale of divergent pathways. Nature Rev. Cancer 5, 713–725 (2005)
Flaig, T. W. & Theodorescu, D. Bladder cancer in 2011: the dawn of personalized medicine. Nature reviews . Urology 9, 65–66 (2012)
Knowles, M. A. & Hurst, C. D. Molecular biology of bladder cancer: new insights into pathogenesis and clinical diversity. Nature Rev. Cancer http://dx.doi.org/10.1038/nrc3817 (in the press)
Acknowledgements
We would like to thank S. Tsai, J. Rosen, L. Donehower, M. Brenner and B. Deneen for their editorial suggestions, and grant funding from the National Cancer Institute CA129640, CA175397, V Scholar Award, Dan L Duncan Career Award, and Bladder Cancer Partnership (K.S.C.), CPRIT pre-doctoral fellowship RP101499 (A.V.K.), AUA Research Scholar Award (P.L.H.) and CPRIT training grant RP140102 for SMART program (T.T.R.). This work is supported by the Genomic & RNA Profiling Core (L. White), Cytometry & Cell Sorting Core (NIAID AI036211, NCI CA125123 and RR024574; J. Sederstorm), and Human Tissue Acquisition & Pathology Core (NCI CA125123; M. Ittmann) at Baylor College of Medicine, the Texas Advanced Computing Center at UT Austin and Rice University for providing High Performance Computing resources. We dedicate this work to the memory of J. Pride (a patient with bladder cancer and friend of I. Weissman), who funded the original work of K.S.C., which served as an important foundation leading to the current study.
Author information
Authors and Affiliations
Contributions
K.S.C. and A.V.K. conceived, designed and performed data analysis. A.V.K., J.X. and S.P. performed the functional xenograft tumour experiments. A.V.K., J.X., E.L. and P.L.H. performed molecular analyses experiments. A.V.K. and J.X. performed, and Q.M. and F.C. analysed, the RNA-seq studies. S.P.L. provided human samples. R.K. coordinated human sample acquisition, R.K., P.L.H. and T.T.R. performed immunohistochemical and pathological analysis. K.S.C. and A.V.K. wrote the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Extended data figures and tables
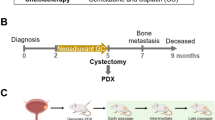
Extended Data Figure 1 Human bladder urothelial carcinomas and their CK14 status pre- and post-neoadjuvant chemotherapy.
a, Table summarizing the clinical information of 15 patients with bladder urothelial carcinomas, with paired pre- and post-chemotherapy tissues (n = 15). PDX-3 was derived from cancer tissues from patient 3 (bold). b, Patient subgroups showing an enrichment (increase) or persistence (maintain high expression) (left) or a resolution (absence) (right) of CK14 staining after chemotherapy treatment. c, d, Representative images of CK14 immunohistochemistry staining (original magnification, ×20), sub-classified as enrichment/persistence (c) and resolution (d) of CK14 staining in matching bladder urothelial carcinoma tissues obtained pre- and post-neoadjuvant chemotherapy. Representative staining patterns of CK14 high-infiltrative staining (>25%), CK14 low-infiltrative staining (<25%), CK14 basal-restricted staining, and CK14 resolution are individually indicated.
Extended Data Figure 2 Cytotoxic chemotherapy induces CK14+ CSC proliferation despite reducing tumour size.
a, Relative change in xenograft tumour volume from multiple xenograft tumour lines in response to GC chemotherapy or vehicle treatment (n = 6 per group). Xenograft tumours were derived from primary urothelial carcinoma patients (PDX-1 and PDX-2). b, Quantification of the percentage of CK14+ cancer cells in chemotherapy-treated and vehicle-treated xenograft tumours. c, Representative images demonstrating immunofluorescence staining of CK14+ cancer cells in chemotherapy-treated and vehicle-treated xenograft tumours. d, Representative images demonstrating immunofluorescence staining of pHH3 (red) and CK14 (green) in chemotherapy-treated and vehicle-treated xenograft tumours. Yellow arrows indicate CK14−pHH3+ cells, white arrows indicate CK14+pHH3+ cancer cells. e, Graph quantifying the change in phospho-histone H3 positive (pHH3+) proliferating cells within CK14+ cancer cells (n = 6 per group). All data represent mean ± s.e.m. Box plots in b show twenty-fifth to seventy-fifth percentiles, with line indicating the median and whiskers indicating the smallest and the largest values. *P < 0.05; **P < 0.01; ***P < 0.001 (two-tailed Student’s t-test). Scale bar, 100 μm.
Extended Data Figure 3 Methodology to purify CK14+ cells for functional evaluation of sphere-forming and tumorigenic properties.
a, Generation of a reporter construct to isolate viable CK14+ bladder cancer cells by FACS. A previously reported and validated human KRT14 gene promoter fragment was subcloned into a promoterless lentiviral construct that encodes a red fluorescent protein, DD-tdTomato. b–d, T24 high-grade urothelial carcinoma cells were stably transduced with the KRT14 reporter construct. FACS analysis validated that Tm+ CK14+ cells (red) represent a subpopulation of previously reported CD44+CD49f+ tumorigenic cells. e, Purified Tm+ CK14+ cancer cells demonstrated greater sphere-forming ability than Tm− CK14−cancer cells in vitro (biological duplicates). f, Tm+ CK14+ bladder cancer cells are approximately 60-fold enriched for tumorigenic cells when engrafted in immunocompromised mice in vivo. Summary of tumour engraftment efficiency and image demonstrating tumour size after transplantation of 10, 50, 500 and 5,000 Tm+ CK14+ or Tm− CK14− cancer cells as purified by FACS. Data represent mean and range (e) and mean ± s.e.m (f). ***P < 0.001 (two-tailed Student’s t-test).
Extended Data Figure 4 Cell viability of purified Tm+ CK14+ and Tm− CK14− cancer cells after GC chemotherapy treatment in vitro (raw FACS data for Fig. 1h).
Dot plots depict FACS analyses showing cell viability of Tm+ CK14+ (red) and Tm− CK14−(black) cancer cells following 11 consecutive days of chemotherapy treatment in vitro. The percentage of viable cells defined as annexin V− PI− is shown in the bottom left quadrant of each plot. Experiments were performed in biological duplicates.
Extended Data Figure 5 Cell cycle profiles of purified Tm+ CK14+ and Tm− CK14− cancer cells after GC chemotherapy in vitro (raw FACS data for Fig. 1i).
Histogram plots depict original FACS analyses of cell cycle profiles from Tm+ CK14+ (red) and Tm− CK14− (black) cancer cells after 11 consecutive days of chemotherapy treatment in vitro. Experiment was performed in biological duplicates. PI, propidium iodide.
Extended Data Figure 6 LRCCs are mutually exclusive to active proliferative cancer cells.
a, b, Immunofluorescence staining to locate LRCCs (green, IdU+) at 0, 4 and 8 weeks of chase periods in patient-derived urothelial carcinoma xenograft (PDX-1) (a) and xenograft established from T24 high-grade urothelial carcinoma cells (b). c, d, Bar graph quantifying the percentage of LRCCs in patient-derived xenograft (PDX-1) (c) and immortalized cancer xenograft (T24) (d) at various chase periods (n = 4). e–g, Immunofluorescence staining to evaluate the localization of LRCCs (green, IdU+) and proliferating cells (red) using CldU (e), proliferating cell nuclear antigen (PCNA; f) or phospho-histone H3 (pHH3; g) in high-grade urothelial carcinoma (T24) at steady state. h, Immunofluorescence co-staining to locate IdU+ LRCCs, CldU+ proliferating and CK14+ cancer cells at various time points after GC chemotherapy. Data shown in c and d represent mean ± s.e.m. Scale bars, 100 μm.
Extended Data Figure 7 Celecoxib abrogates CK14+ cancer cell enrichment after GC chemotherapy in T24 and PDX-3 xenografts.
a, In vivo treatment protocol recapitulating clinical regimen of one chemotherapy cycle in the presence or absence of celecoxib treatment. b, Immunofluorescence staining examining the percentage of CK14+ cancer cells in representative T24 xenograft tumours from various treatment groups. Scale bars, 1,000 μm. c, Immunohistochemical staining examining the percentage of CK14+ cancer cells in representative PDX-3 xenograft tumours from various treatment groups. Scale bars, 1,000 μm. Images in b and c are representative of n = 6 tumours analysed for each treatment group.
Extended Data Figure 8 Co-administration of aspirin diminishes thrombosis without impairing adjuvant effect of celecoxib.
a, Antithrombotic effect of aspirin measurable by tail bleeding time. b, Temporal percentage change in tumour size after two cycles of celecoxib plus GC combination chemotherapy in the presence or absence of aspirin (n = 12 per group). c, Dynamics of COX1 and COX2 expression after GC chemotherapy, data shown for T24 cancer cells (displayed are representative blots from n = 3 experiments). All data represent mean ± s.e.m., tumour volume measurements shown in b are relative to mean tumour volume at day 0. *P < 0.05; **P < 0.01; ***P < 0.001 (one-way ANOVA followed by Dunnett’s test for multiple comparisons); ns, not significant.
Extended Data Figure 9 Enrichment of the ‘wound-response gene signature’ in chemoresistant bladder urothelial carcinomas.
a, GSEA validated an enrichment of the ‘wound-response gene signature’ (GO:0009611) in a panel of non-responding (or chemoresistant) human urothelial bladder carcinomas (n = 20; GSE48277), by comparing post-chemotherapy to pre-chemotherapy cancer tissues. Heat map demonstrates part of the genes within leading edge, including the COX2 gene PTGS2. b, Enlarged heat map for Fig. 4g. c, Enlarged heat map for Fig. 4h. d, Bioinformatics analysis of chemoresistant cancers in a panel of non-responding (or chemoresistant) human urothelial bladder carcinomas (n = 20; GSE48277) validated a significant increase of PTGS2 in post-chemotherapy tissues in comparison to matching pre-chemotherapy tissues. e, Dot plots representing the scoring of COX2 staining in bladder urothelial carcinoma tissues obtained pre-neoadjuvant chemotherapy in two subgroups of patients with different response to neoadjuvant chemotherapy (subgroups described in Extended Data Fig. 1a–c).
Extended Data Figure 10 Schematic model: recurrent CSC repopulation and its manifestation of chemoresistance.
Cytotoxic chemotherapy effectively induces apoptosis but paradoxically elicits a wound response of bladder cancer stem cells to proliferate and repopulate residual tumours. Release of PGE2 from neighbouring apoptotic cancer cells is sufficient to promote this CSC repopulation. In vivo administration of celecoxib effectively abolishes this PGE2/COX2-mediated wound response gene signature, and attenuates progressive manifestation of chemoresistance in preclinical models of human urothelial carcinomas.
Rights and permissions
About this article

Cite this article
Kurtova, A., Xiao, J., Mo, Q. et al. Blocking PGE2-induced tumour repopulation abrogates bladder cancer chemoresistance. Nature 517, 209–213 (2015). https://doi.org/10.1038/nature14034
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature14034
This article is cited by
-
LASS2 enhances chemosensitivity to cisplatin by inhibiting PP2A-mediated β-catenin dephosphorylation in a subset of stem-like bladder cancer cells
BMC Medicine (2024)
-
Lipids as mediators of cancer progression and metastasis
Nature Cancer (2024)
-
Cancer stem cell fate determination: mito-nuclear communication
Cell Communication and Signaling (2023)
-
Regulation and signaling pathways in cancer stem cells: implications for targeted therapy for cancer
Molecular Cancer (2023)
-
Eta polycaprolactone (ε-PCL) implants appear to cause a partial differentiation of breast cancer lung metastasis in a murine model
BMC Cancer (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
