
Abstract
Hepatitis A virus (HAV) remains enigmatic, despite 1.4 million cases worldwide annually1. It differs radically from other picornaviruses, existing in an enveloped form2 and being unusually stable, both genetically and physically3, but has proved difficult to study. Here we report high-resolution X-ray structures for the mature virus and the empty particle. The structures of the two particles are indistinguishable, apart from some disorder on the inside of the empty particle. The full virus contains the small viral protein VP4, whereas the empty particle harbours only the uncleaved precursor, VP0. The smooth particle surface is devoid of depressions that might correspond to receptor-binding sites. Peptide scanning data extend the previously reported VP3 antigenic site4, while structure-based predictions5 suggest further epitopes. HAV contains no pocket factor and can withstand remarkably high temperature and low pH, and empty particles are even more robust than full particles. The virus probably uncoats via a novel mechanism, being assembled differently to other picornaviruses. It utilizes a VP2 ‘domain swap’ characteristic of insect picorna-like viruses6,7, and structure-based phylogenetic analysis places HAV between typical picornaviruses and the insect viruses. The enigmatic properties of HAV may reflect its position as a link between ‘modern’ picornaviruses and the more ‘primitive’ precursor insect viruses; for instance, HAV retains the ability to move from cell-to-cell by transcytosis8,9.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others


References
WHO. Hepatitis A. Fact Sheet No. 328 http://www.who.int/mediacentre/factsheets/fs328/en/ (2013)
Feng, Z. et al. A pathogenic picornavirus acquires an envelope by hijacking cellular membranes. Nature 496, 367–371 (2013)
Siegl, G., Weitz, M. & Kronauer, G. Stability of hepatitis A virus. Intervirology 22, 218–226 (1984)
Ping, L. H. & Lemon, S. M. Antigenic structure of human hepatitis A virus defined by analysis of escape mutants selected against murine monoclonal antibodies. J. Virol. 66, 2208–2216 (1992)
Borley, D. W. et al. Evaluation and use of in-silico structure-based epitope prediction with foot-and-mouth disease virus. PLoS ONE 8, e61122 (2013)
Tate, J. et al. The crystal structure of cricket paralysis virus: the first view of a new virus family. Nature Struct. Biol. 6, 765–774 (1999)
Squires, G. et al. Structure of the Triatoma virus capsid. Acta Crystallogr. D. 69, 1026–1037 (2013)
Dotzauer, A., Brenner, M., Gebhardt, U. & Vallbracht, A. IgA-coated particles of Hepatitis A virus are translocalized antivectorially from the apical to the basolateral site of polarized epithelial cells via the polymeric immunoglobulin receptor. J. Gen. Virol. 86, 2747–2751 (2005)
Dotzauer, A. et al. Hepatitis A virus-specific immunoglobulin A mediates infection of hepatocytes with hepatitis A virus via the asialoglycoprotein receptor. J. Virol. 74, 10950–10957 (2000)
Deinhardt, F. Prevention of viral hepatitis A: past, present and future. Vaccine 10 (suppl. 1). 10–14 (1992)
Lemon, S. M. & Binn, L. N. Antigenic relatedness of two strains of hepatitis A virus determined by cross-neutralization. Infect. Immun. 42, 418–420 (1983)
Graff, J. et al. Hepatitis A virus capsid protein VP1 has a heterogeneous C terminus. J. Virol. 73, 6015–6023 (1999)
Tesar, M., Jia, X. Y., Summers, D. F. & Ehrenfeld, E. Analysis of a potential myristoylation site in hepatitis A virus capsid protein VP4. Virology 194, 616–626 (1993)
Martin, A. & Lemon, S. M. Hepatitis A virus: from discovery to vaccines. Hepatology 43, S164–S172 (2006)
Kaplan, G. et al. Identification of a surface glycoprotein on African green monkey kidney cells as a receptor for hepatitis A virus. EMBO J. 15, 4282–4296 (1996)
Bishop, N. E. & Anderson, D. A. RNA-dependent cleavage of VP0 capsid protein in provirions of hepatitis A virus. Virology 197, 616–623 (1993)
Wang, X. et al. A sensor-adaptor mechanism for enterovirus uncoating from structures of EV71. Nature Struct. Mol. Biol. 19, 424–429 (2012)
Rossmann, M. G. et al. Structure of a human common cold virus and functional relationship to other picornaviruses. Nature 317, 145–153 (1985)
Siegl, G., Frosner, G. G., Gauss-Muller, V., Tratschin, J. D. & Deinhardt, F. The physicochemical properties of infectious hepatitis A virions. J. Gen. Virol. 57, 331–341 (1981)
De Colibus, L. et al. More-powerful virus inhibitors from structure-based analysis of HEV71 capsid-binding molecules. Nature Struct. Mol. Biol. 21, 282–288 (2014)
Acharya, R. et al. The three-dimensional structure of foot-and-mouth disease virus at 2.9 Å resolution. Nature 337, 709–716 (1989)
Liu, Y. & Eisenberg, D. 3D domain swapping: as domains continue to swap. Protein Sci. 11, 1285–1299 (2002)
Walter, T. S. et al. A plate-based high-throughput assay for virus stability and vaccine formulation. J. Virol. Methods 185, 166–170 (2012)
Porta, C. et al. Rational engineering of recombinant picornavirus capsids to produce safe, protective vaccine antigen. PLoS Pathog. 9, e1003255 (2013)
Filman, D. J. et al. Structural factors that control conformational transitions and serotype specificity in type 3 poliovirus. EMBO J. 8, 1567–1579 (1989)
Warwicker, J. Model for the differential stabilities of rhinovirus and poliovirus to mild acidic pH, based on electrostatics calculations. J. Mol. Biol. 223, 247–257 (1992)
Garriga, D. et al. Insights into minor group rhinovirus uncoating: the X-ray structure of the HRV2 empty capsid. PLoS Pathog. 8, e1002473 (2012)
Ren, J. et al. Picornavirus uncoating intermediate captured in atomic detail. Nature Commun. 4, 1929 (2013)
Butan, C., Filman, D. J. & Hogle, J. M. Cryo-electron microscopy reconstruction shows poliovirus 135S particles poised for membrane interaction and RNA release. J. Virol. 88, 1758–1770 (2014)
Riffel, N. et al. Atomic resolution structure of Moloney murine leukemia virus matrix protein and its relationship to other retroviral matrix proteins. Structure 10, 1627–1636 (2002)
Walter, T. S. et al. A procedure for setting up high-throughput nanolitre crystallization experiments. Crystallization workflow for initial screening, automated storage, imaging and optimization. Acta Crystallogr. D 61, 651–657 (2005)
Minor, W., Cymborowski, M., Otwinowski, Z. & Chruszcz, M. HKL-3000: the integration of data reduction and structure solution—from diffraction images to an initial model in minutes. Acta Crystallogr. D 62, 859–866 (2006)
Brünger, A. T. et al. Crystallography & NMR system: A new software suite for macromolecular structure determination. Acta Crystallogr. D 54, 905–921 (1998)
Emsley, P., Lohkamp, B., Scott, W. G. & Cowtan, K. Features and development of Coot. Acta Crystallogr. D 66, 486–501 (2010)
Stuart, D. I., Levine, M., Muirhead, H. & Stammers, D. K. Crystal structure of cat muscle pyruvate kinase at a resolution of 2.6 A. J. Mol. Biol. 134, 109–142 (1979)
PyMOL. The PyMOL Molecular Graphics System, Version 1.5.0.4 Schrödinger, LLC. (2002)
Schuck, P. Size-distribution analysis of macromolecules by sedimentation velocity ultracentrifugation and lamm equation modeling. Biophys. J. 78, 1606–1619 (2000)
Lawrence, M. C. & Colman, P. M. Shape complementarity at protein/protein interfaces. J. Mol. Biol. 234, 946–950 (1993)
Chapman, M. S. & Rossmann, M. G. Comparison of surface properties of picornaviruses: strategies for hiding the receptor site from immune surveillance. Virology 195, 745–756 (1993)
Acknowledgements
This work was supported by the Strategic Priority Research Program of the Chinese Academy of Sciences, grant no. XDB08020200, the Ministry of Science and Technology 973 Project (grant no. 2014CB542800), the National Major Project of Infectious Disease (2012ZX10004701) and National Science Foundation grant 81330036. D.I.S. and E.E.F. are supported by the UK Medical Research Council (grant G1000099) and J.R. by the Wellcome Trust. This work is a contribution from the Instruct Centre, Oxford. Administrative support was provided by the Wellcome Trust (075491/Z/04). We thank J. Johnson and A. Routh for supplying CrPV, and S. Lemon and K. McKnight for discussions.
Author information
Authors and Affiliations
Contributions
Q.G., J.W., Z.H. and W.Y. prepared samples. X.W., J.R., Q.G., X.L., Y.S., E.E.F. and D.I.S. performed research, J.R., X.W., E.E.F. and D.I.S. analysed data and with D.J.R. and Z.R. wrote the manuscript, in discussion with J.W. and Z.H.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Extended data figures and tables
Extended Data Figure 1 HAV purification and characterization.
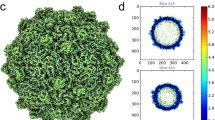
a, Zonal ultracentrifugation of a 15% to 45% (w/v) sucrose density gradient at 103,614 g for 3.5 h was used to purify HAV, as described in Methods. Two predominant particle types were separated; one was located at ∼27% sucrose, the other at ∼32% sucrose. The absorbance ratios (λ260/λ280) were 0.76 for the top band and 1.66 for the bottom band, indicating that the former contained mainly empty particles without the RNA genome and the latter mainly full particles with the RNA genome. The top band was much broader than the bottom one. Three and two fractions were collected around the top and the bottom bands, respectively, for further purification. b, SDS–PAGE for protein composition analysis using A NuPAGE 4–12% Bis-Tris Gel (Invitrogen). Each lane was loaded with 5–10 μg sample (lane 1, full particles; lane 2, empty particles; lane 3, markers). The calculated molecular masses of VP0, VP1, VP2, VP3 and VP4 were 27.26 kDa, 30.73 kDa, 24.80 kDa, 27.86 kDa and 2.50 kDa, respectively. The results appear to show incomplete cleavage of VP0 in the full particles and no VP0 cleavage in the empty particles. c, d, Sedimentation velocity experiments were performed on a Beckman XL-I analytical ultracentrifuge at 20 °C. Samples were loaded into a conventional double-sector quartz cell and mounted in a Beckman four-hole An-60 Ti rotor. Data were collected at 15,000 r.p.m. at a wavelength of 287 nm. Interference sedimentation coefficient distributions were calculated from the sedimentation velocity data using SEDFIT37. The full particle has a sedimentation coefficient of 144S and the empty particle 82S. e, f, Negative stain electron microscopy of HAV particles. Particles from the top band in panel a, shown in panel f, suggest that light sedimentation fractions were mainly composed of empty particles. Note that some particles appear to have external features. Panel e shows the heavy particles, which appear to contain viral RNA.
Extended Data Figure 2 HAV capsid protein structure.
a–c, Stereo diagrams showing the structures of the capsid proteins VP1, VP2 and VP3, respectively. The Cα backbone is shown as a thin line, the N and C termini are labelled, every 10th residue is marked with a small sphere and every 20th is numbered. d, VP1 is initially produced with an 8-kDa C-terminal extension (known as PX) unique to HAV among picornaviruses. PX is cleaved from the full particle by an unknown host protease. According to the data shown in Extended Data Fig. 1 and the electron density maps, the particles we have analysed do not contain PX, but if it were present we would expect its course to start at the purple positions (C termini of VP1) on the surface of the virus.
Extended Data Figure 3 Antigenicity of full and empty particles.
The reactivity of HAV full and empty particles against a panel of six purified HAV neutralizing monoclonal antibodies was measured by ELISA. The bar charts represent the average OD450 reading for the six monoclonal antibodies at each dilution with the standard deviation shown as an error bar.
Extended Data Figure 4 PaSTRy assays.
To characterize the stability of HAV full and empty particles compared to CrPv across the pH range from 2.0 to 10.0, differential scanning fluorimetry assays were performed with dyes SYTO9 (to detect RNA exposure) and SYPRO red (to detect protein melting)23. a, The raw fluorescence traces of HAV full particles incubated with SYTO9. b, The raw fluorescence traces of HAV full particles incubated with SYPRO red. c, The raw fluorescence traces of HAV empty particles incubated with SYPRO red. d, The raw fluorescence traces of CrPV full particles incubated with SYTO9 across the same pH range. The colour scheme is dark red (pH 2.0), red (pH 3.0), orange (pH 4.0), yellow (pH 5.0), green (pH 6.0), sky blue (pH 7.0), blue (pH 8.0), dark blue (pH 9.0) and purple (pH 10.0). Since the SYTO9 dye didn’t function well below pH 4.0 the fluorescence traces for pH 2.0 and pH 3.0 are omitted. These results indicate that HAV full virions are most stable at pH 5.0 and RNA genome release occurs at about 76 °C and protein melting at 77 °C; that is, there is no notable transition between RNA release and particle loss. HAV empty particles show a similar trend but appear to withstand temperatures up to 81 °C in low pH buffer. In contrast CrPV is most stable at pH 4.0 and RNA genome release occurs at about 54 °C.
Extended Data Figure 5 Interactions between α1 helices of VP2 at the icosahedral two-fold.
a, The close-packing of the two helices is shown, in particular the packing of Tyr 100 against the adjacent helix. The contact area between these helices and the surface complementarity38 suggests a well-fitting interface for HAV (contact area 83 Å2, surface complimentarity 0.755; EV71, 111/0.550; Polio-1, 124/0.785; CrPV, 72/0.707; FMDV, 50/0.593). The interface between protomers forming the pentameric units is similar for all of these viruses (HAV, 4,267 Å2; Polio-1, 4369 Å2; CrPV, 4,647 Å2; EV71, 4,131 Å2; FMDV-A22, 3,432 Å2). b, Helical wheel diagrams for helix α1 of several picornaviruses showing the unusually small side chains at the helix interface in HAV.
Extended Data Figure 6 Antigenicity and YPX3L ALIX-interacting motifs.
a, Previously indicated antigenic sites of HAV are mapped onto the structure (pink spheres). Late domain YPX3L motifs are shown as orange spheres. b–d, Surface maps of HAV generated using RIVEM39 such that area for each residue as drawn corresponds to its accessible area and the coloration is according to the radius from the virus centre (blue, deepest; red, most exposed). On these the antigenic sites of HAV are depicted in purple. b, Previously reported antigenic sites; c, predicted sites5; d, new sites determined by peptide mapping (indigo) together with previously identified sites (purple).
Extended Data Figure 7 Monoclonal antibody neutralization assays and peptide epitope mapping.
a, In vitro neutralization assays of monoclonal antibodies against HAV (TZ84). b, Peptides used for epitope mapping. c, Reactivity of the neutralizing mAb 11 against synthetic peptides measured by peptide-ELISA.
Supplementary information
In-situ crystal analysis: movement on exposure to X-rays.
A trapezoidal plate-shaped crystal of HAV (100×100×5 μm, thin axis parallel to the X-ray beam), in mother liquor containing 30% MPD, was exposed for 0.1 s to full beam (∼1012 photons s-1) at I24, Diamond at room temperature. The real-time video (viewed co-axial to the X-ray beam) shows the crystal darting upwards and left away from the beam (marked by white cross-hairs) once the exposure begins (at 3 s) before returning to its starting position. The pale grey scale bar bottom right shows 50 μm horizontally. (MOV 8553 kb)
Rights and permissions
About this article

Cite this article
Wang, X., Ren, J., Gao, Q. et al. Hepatitis A virus and the origins of picornaviruses. Nature 517, 85–88 (2015). https://doi.org/10.1038/nature13806
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature13806
This article is cited by
-
Cell entry and release of quasi-enveloped human hepatitis viruses
Nature Reviews Microbiology (2023)
-
Hepatitis A virus infection
Nature Reviews Disease Primers (2023)
-
Metagenomic analysis of viral diversity and a novel astroviruse of forest rodent
Virology Journal (2022)
-
Oligomers of hepatitis A virus (HAV) capsid protein VP1 generated in a heterologous expression system
Microbial Cell Factories (2022)
-
Structural basis for neutralization of an anicteric hepatitis associated echovirus by a potent neutralizing antibody
Cell Discovery (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
