
Abstract
Blood vessel growth in the skeletal system and osteogenesis seem to be coupled, suggesting the existence of molecular crosstalk between endothelial and osteoblastic cells1,2. Understanding the nature of the mechanisms linking angiogenesis and bone formation should be of great relevance for improved fracture healing or prevention of bone mass loss. Here we show that vascular growth in bone involves a specialized, tissue-specific form of angiogenesis. Notch signalling promotes endothelial cell proliferation and vessel growth in postnatal long bone, which is the opposite of the well-established function of Notch and its ligand Dll4 in the endothelium of other organs and tumours3,4. Endothelial-cell-specific and inducible genetic disruption of Notch signalling in mice not only impaired bone vessel morphology and growth, but also led to reduced osteogenesis, shortening of long bones, chondrocyte defects, loss of trabeculae and decreased bone mass. On the basis of a series of genetic experiments, we conclude that skeletal defects in these mutants involved defective angiocrine release of Noggin from endothelial cells, which is positively regulated by Notch. Administration of recombinant Noggin, a secreted antagonist of bone morphogenetic proteins, restored bone growth and mineralization, chondrocyte maturation, the formation of trabeculae and osteoprogenitor numbers in endothelial-cell-specific Notch pathway mutants. These findings establish a molecular framework coupling angiogenesis, angiocrine signals and osteogenesis, which may prove significant for the development of future therapeutic applications.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others

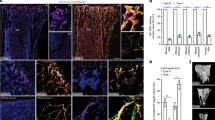
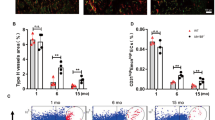
Change history
19 March 2014
Minor edits were made to panel labelling in the Fig. 3 legend.
References
Eshkar-Oren, I. et al. The forming limb skeleton serves as a signaling center for limb vasculature patterning via regulation of Vegf. Development 136, 1263–1272 (2009)
Maes, C. et al. Osteoblast precursors, but not mature osteoblasts, move into developing and fractured bones along with invading blood vessels. Dev. Cell 19, 329–344 (2010)
Benedito, R. et al. The notch ligands Dll4 and Jagged1 have opposing effects on angiogenesis. Cell 137, 1124–1135 (2009)
Noguera-Troise, I. et al. Blockade of Dll4 inhibits tumour growth by promoting non-productive angiogenesis. Nature 444, 1032–1037 (2006)
Skawina, A., Litwin, J. A., Gorczyca, J. & Miodonski, A. J. The vascular system of human fetal long bones: a scanning electron microscope study of corrosion casts. J. Anat. 185, 369–376 (1994)
Trueta, J. & Morgan, J. D. The vascular contribution to osteogenesis. I. Studies by the injection method. J. Bone Joint Surg. Br. 42-B, 97–109 (1960)
Kuhn, A. et al. Expression of endomucin, a novel endothelial sialomucin, in normal and diseased human skin. J. Invest. Dermatol. 119, 1388–1393 (2002)
Kusumbe, A. P., Ramasamy, S. K. & Adams, R. H. Coupling of angiogenesis and osteogenesis by a specific vessel subtype in bone. Nature http://dx.doi.org/10.1038/nature13145 (this issue)
Siekmann, A. F. & Lawson, N. D. Notch signalling limits angiogenic cell behaviour in developing zebrafish arteries. Nature 445, 781–784 (2007)
Hoeck, J. D. et al. Fbw7 controls neural stem cell differentiation and progenitor apoptosis via Notch and c-Jun. Nature Neurosci. 13, 1365–1372 (2010)
Izumi, N. et al. Fbxw7 controls angiogenesis by regulating endothelial Notch activity. PLoS ONE 7, e41116 (2012)
Nakashima, K. et al. The novel zinc finger-containing transcription factor osterix is required for osteoblast differentiation and bone formation. Cell 108, 17–29 (2002)
Hozumi, K. et al. Delta-like 1 is necessary for the generation of marginal zone B cells but not T cells in vivo. Nature Immunol. 5, 638–644 (2004)
Koch, U. et al. Delta-like 4 is the essential, nonredundant ligand for Notch1 during thymic T cell lineage commitment. J. Exp. Med. 205, 2515–2523 (2008)
Brooker, R., Hozumi, K. & Lewis, J. Notch ligands with contrasting functions: Jagged1 and Delta1 in the mouse inner ear. Development 133, 1277–1286 (2006)
Olave, I., Reinberg, D. & Vales, L. D. The mammalian transcriptional repressor RBP (CBF1) targets TFIID and TFIIA to prevent activated transcription. Genes Dev. 12, 1621–1637 (1998)
Gerber, H. P. et al. VEGF couples hypertrophic cartilage remodeling, ossification and angiogenesis during endochondral bone formation. Nature Med. 5, 623–628 (1999)
Engin, F. et al. Dimorphic effects of Notch signaling in bone homeostasis. Nature Med. 14, 299–305 (2008)
Hilton, M. J. et al. Notch signaling maintains bone marrow mesenchymal progenitors by suppressing osteoblast differentiation. Nature Med. 14, 306–314 (2008)
Potente, M., Gerhardt, H. & Carmeliet, P. Basic and therapeutic aspects of angiogenesis. Cell 146, 873–887 (2011)
Wu, X. B. et al. Impaired osteoblastic differentiation, reduced bone formation, and severe osteoporosis in noggin-overexpressing mice. J. Clin. Invest. 112, 924–934 (2003)
Canalis, E., Brunet, L. J., Parker, K. & Zanotti, S. Conditional inactivation of noggin in the postnatal skeleton causes osteopenia. Endocrinology 153, 1616–1626 (2012)
Tylzanowski, P., Mebis, L. & Luyten, F. P. The Noggin null mouse phenotype is strain dependent and haploinsufficiency leads to skeletal defects. Dev. Dyn. 235, 1599–1607 (2006)
Wang, Y. et al. Ephrin-B2 controls VEGF-induced angiogenesis and lymphangiogenesis. Nature 465, 483–486 (2010)
Han, H. et al. Inducible gene knockout of transcription factor recombination signal binding protein-J reveals its essential role in T versus B lineage decision. Int. Immunol. 14, 637–645 (2002)
Murtaugh, L. C., Stanger, B. Z., Kwan, K. M. & Melton, D. A. Notch signaling controls multiple steps of pancreatic differentiation. Proc. Natl Acad. Sci. USA 100, 14920–14925 (2003)
Soleimani, M. & Nadri, S. A protocol for isolation and culture of mesenchymal stem cells from mouse bone marrow. Nature Protocols 4, 102–106 (2009)
Zhu, H. et al. A protocol for isolation and culture of mesenchymal stem cells from mouse compact bone. Nature Protocols 5, 550–560 (2010)
Suzuki, H. et al. Histological evidence of the altered distribution of osteocytes and bone matrix synthesis in klotho-deficient mice. Arch. Histol. Cytol. 68, 371–381 (2005)
Acknowledgements
We thank A. Duarte, F. Radtke and T. Honjo for floxed Dll4 and Rbpj mutant mice, A. Borgscheiper for technical assistance, M. Stehling for endothelial cell sorting, D. Zeuschner for electron microscopy. Funding was provided by the Max Planck Society, the University of Münster, the DFG cluster of excellence ‘Cells in Motion’, and the European Research Council (AdG 339409 AngioBone).
Author information
Authors and Affiliations
Contributions
S.K.R., A.P.K and R.H.A. designed experiments and interpreted results. S.K.R. generated and characterized mouse mutant lines. S.K.R. and A.P.K. did all experiments including FACS, qRT-PCR, bone sectioning and staining, confocal imaging and quantifications. L.W. performed analysis of bone vessel ultrastructure. S.K.R. and R.H.A. wrote the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Extended data figures and tables
Extended Data Figure 1 Schematic representation of key findings.
Organization and role of growing vessels in the regulation of osteogenesis in postnatal long bone. Endothelial columns, which are embedded between segments of forming trabecular bone in the metaphysis, are interconnected by arches at their distal end. Blind-ended, lumen-containing protrusions extend from arches towards growth plate chondrocytes, a key source of VEGF-A. Endothelial Notch signalling promotes endothelial cell proliferation and vessel growth in bone, which is the opposite of its role in other tissues. Notch activity in endothelial cells is also required for endothelial Noggin expression, controls the differentiation of perivascular osteoprogenitor cells and thereby osteogenesis. Endothelial Notch signalling and Noggin also promote chondrocyte maturation and hypertrophy, which affects angiogenesis through VEGF-A expression. These signalling interactions between different cell types couple angiogenesis and osteogenesis.
Extended Data Figure 2 Vessel growth in the postnatal metaphysis.
a, b, Organisation of distal vessels in the metaphysis of 4-week-old tibia. Endothelial cells were visualized by anti-Endomucin (Emcn, red) immunostaining (a) or GFP expression (green) in Cdh5(PAC)-CreERT2 Rosa26-mT/mG double transgenic mice (b) at 4 weeks of age. Note blunt appearance of most distal vessels (top) in proximity of growth plate chondrocytes. Nuclei, blue (DAPI). c, Maximum intensity projection showing the organization of distal, CD31-immunostained (type H) vessels in the tibial metaphysis at the indicated ages. Note emergence of blunt and blind-ended bulb-like protrusions (arrows) from arch vessels (arrowheads) at the distal end of endothelial columns. d, Confocal image showing expression of intercellular adhesion molecule 2 (ICAM2, red), a marker of lumenized vessels, in distal vessels of the tibial metaphysis. Nuclei, blue (DAPI). e, Podocalyxin (Podx, green), a sialoglycoprotein marking the apical surface of endothelial cells and thereby the vascular lumen (arrows), is present on the most distal, CD31-positive (red) vessel structures in the metaphysis. Nuclei, blue (DAPI). f, Transmission electron microscopy confirming the lumenized nature of distal vessels close to growth plate chondrocytes (ch). Yellow arrowheads indicate thin endothelial cells lining vessels. g, Quantitation of EdU labelled (proliferating) endothelial cells in the metaphysis of long bone. EdU+ endothelial cells were predominantly present in columnar vessels and were comparably rare (≤5%) in vascular arches (n = 5 mice from 3 independent experiments). Error bars, ± s.e.m. P values, two-tailed unpaired t-test.
Extended Data Figure 3 Marker expression in arch and bulb endothelial cells.
a, Quantitative real time PCR analysis of transcripts encoding vessel guidance molecules in type H and type L endothelial cells isolated by FACS from 4-week-old femur. Note high levels of transcripts for Netrin-1 (Ntn1), Netrin-3 (Ntn3), Neuropilin 1 (Nrp1), Plexin D1 (Plxnd1), Unc5b and Robo4 in type H relative to type L endothelium (n = 5 mice from 4 independent experiments). Error bars, ± s.e.m. P values, two-tailed unpaired t-test. b–f, Confocal images showing immunostaining of vessel guidance molecules such as Neuropilin 1 (b, Nrp1), Neuropilin 2 (c, Nrp2), Plexin D1 (d), Unc5b (e), Robo4 (f) in metaphyseal (type H) vessels and surrounding mesenchymal cells. High levels of Nrp1 were detected in type H endothelial cells (arrows), but comparably lower expression was also observed in vessel-associated osteoprogenitors and growth plate chondrocytes (ch). Nrp2 is highly expressed in perivascular osteoprogenitors (arrowheads). High expression of PlexinD1 and Unc5b, and lower levels of Robo4 were detected in type H vessel bulbs and columns (arrows). g, Maximum intensity projection showing high expression of VEGFR3 in endothelial bulbs and arches (white arrows) compared to columns. VEGFR3 was absent in the arteries. h, Image showing Dll4 expression in type H endothelial cells forming bulbs and arches in 4-week-old tibial metaphysis. With the exception of arteries (arrows), Dll4 expression was low in vessels of the diaphysis. i, Jag1 expression was detected in endothelial cells (white arrows) and surrounding mesenchymal cells.
Extended Data Figure 4 EC numbers, proliferation and type H vessels in Notch mutants.
a, b, Representative flow-cytometric graph plots showing the quantification of total and EdU-labelled endothelial cells from RbpjiΔEC (a) or Fbxw7iΔEC (b) mutant bones or corresponding littermate controls, as indicated. CD31+ CD45− Ter119− endothelial cells were substantially reduced in RbpjiΔEC mutants (a). Conversely, endothelial cell number and proliferation were increased after inactivation of Fbxw7 (b). c, Maximum intensity projections of EdU-labelled (green fluorescence) tibial metaphysis showing proliferating cells in 4-week-old Notch gain-of-function (Fbxw7iΔEC) and loss-of-function (RbpjiΔEC) mutants and corresponding littermate controls. Endothelial cells were visualized by Emcn immunostaining (red) and nuclei by DAPI (blue). d, Confocal images of Emcn (red) immunostained endothelial distal columns and arches next to growth plate chondrocytes (ch) highlighting the distribution of filopodia (arrowheads). Note that filopodia were directed towards chondrocytes in controls whereas directionality was compromised in RbpjiΔEC mutants. e, Quantitation of filopodia indicating higher variability in RbpjiΔEC samples (n = 6 mice from 4 independent litters). Error bars, ± s.e.m. P values, two-tailed unpaired t-test. f, Quantitative analysis of type H vessels in 4-week-old Fbxw7iΔEC mutants and littermate controls (n = 6 mice from 4 independent litters). Error bars, ± s.e.m. P values, two-tailed unpaired t-test. g, h, Maximum intensity projections of CD31 (green) and Endomucin (red) immunostained vessels in the metaphysis of 4-week-old RbpjiΔEC (g) and Fbxw7iΔEC mutants (h). Note loss of type H vessels and decreased CD31 (green) staining in the RbpjiΔEC metaphysis, while CD31+ vessels were extended after endothelial-cell-specific inactivation of Fbxw7. Arrows indicate arteries.
Extended Data Figure 5 Arterial specification and VEGF receptor expression in Notch mutants.
a, Maximum intensity projections of Emcn (red) and CD31 (green) immunostained sections of 4-week-old tibia showing increase in CD31+ (type H) vessels in Fbxw7iΔEC mice. Mutants also displayed numerous small CD31+ Endomucin− arterioles (white arrows), which were associated with αSMA+ cells (blue arrows in bottom panels). b–d, Confocal images showing 4-week-old RbpjiΔEC or Fbxw7iΔEC mutant, or corresponding littermate control metaphysis after immunostaining for different VEGF receptors. VEGFR2 was highest on control arches, bulbs and arteries and staining was strongly reduced in the RbpjiΔEC metaphysis (b). VEGFR3 immunostaining decorated arches and bulb protrusion but was absent in the arteries. Staining was reduced in RbpjiΔEC mutants but enhanced in Fbxw7iΔEC vessels (c). Expression of VEGFR1 was predominantly found on perivascular mesenchymal and osteoprogenitor cells. Expression in these populations was not appreciably altered (d). e, f, qPCR analysis of sorted endothelial cells from Notch loss-of-function (RbpjiΔEC, e) and gain-of-function (Fbxw7iΔEC, f) mice. In endothelial cells, Notch positively regulated transcripts for the receptors VEGFR2 (Kdr), VEGFR3 (Flt4) and membrane-anchored VEGFR1 (mFlt1). In contrast, expression of soluble Flt1 (sFlt1), a known antagonist of VEGF signalling, was increased in RbpjiΔEC endothelial cells and significantly reduced in Fbxw7iΔEC cells (n = 4 mice from 4 independent litters). Error bars, ± s.e.m. P values, two-tailed unpaired t-test. g, Increased formation of CD31+ (red) vessels in the metaphysis of 4-week-old Notch gain-of-function mice after endothelial-cell-specific overexpression of active Notch (NICDiOE-EC). Nuclei, blue (DAPI). h, Confocal images showing extensive formation of CD31+ (green) Emcn− (red) arterioles (arrows) in the NICDiOE-EC metaphysis. Nuclei, DAPI (blue).
Extended Data Figure 6 Gene expression in Rbpj mutant bone and lung endothelial cells.
a, qPCR analysis of freshly isolated endothelial cells from control and RbpjiΔEC mutant lung or bone. Relative fold mRNA expression of Notch target genes such as Dll4, Jag1, Efnb2, EphB4, Hes1, Hey1, Hey2 is shown. Differently expressed genes are marked in red (n = 4 mice from 4 independent litters). Error bars, ± s.e.m. P values, two-tailed unpaired t-test. b, Quantitative PCR analysis of expression the Notch target gene Hes5 in sorted endothelial cells from 4-week-old wild-type bone and lung (left), and from RbpjiΔEC mutant and littermate control bone samples (right). Note very low expression of Hes5 in lung compared to bone endothelial cells and its reduction in the RbpjiΔEC mutant bone endothelium. Data represent relative fold mRNA expression (n = 4 mice from 4 independent litters). Error bars, ± s.e.m. P values, two-tailed unpaired t-test. c, qPCR results for transcripts encoding intercellular adhesion molecule 1 (Icam1), angiopoietin 2 (Angpt2), VE-cadherin (Cdh5), and CD31 (Pecam1). Data represent relative fold mRNA expression. Differently expressed genes are marked in red (n = 4 mice from 4 independent litters). Error bars, ± s.e.m. P values, two-tailed unpaired t-test. d, Relative fold mRNA expression of cyclin-dependent kinase inhibitor 2A (Cdkn2a), cyclin-dependent kinase inhibitor 1A (Cdkn1a), cyclin-dependent kinase inhibitor 1B (Cdkn1b), cyclin D1 (Ccnd1), cyclin-dependent kinase 2 (Cdk2), and cyclin-dependent kinase 4 (Cdk4) in control or RbpjiΔEC mutant lung and bone endothelial cells (n = 4 mice from 4 independent litters). Error bars, ± s.e.m. P values, two-tailed unpaired t-test.
Extended Data Figure 7 Endothelial Notch controls osteogenesis.
a–i, Analysis of bone parameters in 4-week-old RbpjiΔEC mutants and control littermates. Data on femur length are shown in panel a. Connectivity density (number of connections per unit volume, b), trabecular bone separation (size of space separating trabeculae, c), trabecular number (number of trabeculae per mm length, d) were obtained by μ-CT. Mineral apposition rates (MAR, e) were calculated using calcein double labelling. Data on osteoclast number per bone perimeter (f), osteoclast surface per bone surface (g), osteoid percentage (h) and osteiod thickness (i) are based on histomorphometrical characterization of control and mutant samples (n = 6 mice from 4 independent litters). Error bars, ± s.e.m. P values, two-tailed unpaired t-test. j, Osteoclasts were identified by immunostaining for Calcitonin receptor (green). Maximum intensity projection of Calcitonin receptor stained metaphysis region of tibial sections of Notch gain-of-function (Fbxw7iΔEC) and loss-of-function (RbpjiΔEC) mutants along with their respective littermate controls. Nuclei, DAPI (blue). k–t, Analysis of bone parameters in 4-week-old Fbxw7iΔEC mutants and control littermates. Femur length is shown in k. μ-CT data (l) were used for the analysis of bone density (bone volume/total volume; BV/TV, m), connectivity density (number of connections per unit volume, n), trabecular bone separation (size of space separating trabeculae, o), trabecular number (number of trabeculae per mm length, p), and trabecular thickness (q). Data on osteoclast number per bone perimeter (r), osteoclast surface/bone surface (Osteoclast S/B S) (s), and osteiod thickness (t) are based on histomorphometrical characterization of control and mutant samples (n = 6 mice from 4 independent litters). Error bars, ± s.e.m. P values, two-tailed unpaired t-test.
Extended Data Figure 8 Endothelial Notch controls mesenchymal cell differentiation.
a, Confocal images showing immunostaining for mesenchymal lineage markers Osx (green) and Runx2 (red) in Fbxw7iΔEC and littermate control tibiae, as indicated. Nuclei, DAPI (blue). Note reduction of Osx+ cells and increased Runx2+ population after targeting of Fbxw7 in endothelial cells. b, Analysis of differentiation capacity of primary mesenchymal cells isolated from RbpjiΔEC or control bone. Alizarin Red S staining after 10 days in vitro (10 div) showed that mesenchymal cells isolated from endothelial-cell-specific RbpjiΔEC mutants generated mineral nodules (red arrows) prematurely in comparison to controls. c, d, Maximum intensity projection of stained RbpjiΔEC and control tibiae. Labelling of hypoxic cells with pimonidazole (Pimo, green) showed no appreciable differences between control and RbpjiΔEC samples (c). Likewise, anti-HIF1α immunostaining (green) was comparable in the control and RbpjiΔEC metaphysis (d). Endothelial cells (Emcn antibody staining, red); nuclei, DAPI (blue). e, Representative confocal images showing that endothelial-cell-specific gene targeting of Jagged1 (Jag1) or Delta-like 1 (Dll1) did not lead to appreciable alterations in Jag1iΔEC or Dll1iΔEC mutant metaphyseal vasculature at 4 weeks. Dashed lines indicate vessel columns (C) and distal arches (A). Endothelial cells, CD31 (red); nuclei, DAPI (blue). f, Smaller size of freshly isolated femurs from 4 week-old Dll4iΔEC mutants relative to littermate controls. g, Disrupted organization of vessel arches (A) and columns (C) in the Dll4iΔEC metaphyseal vasculature. Endothelial cells, CD31 (red); nuclei, DAPI (blue). h, Quantitative analysis of size of chondrocyte proliferating zone in littermate control and RbpjiΔEC mutants (n = 6 mice from 4 independent litters). Error bars, ± s.e.m. P values, two-tailed unpaired t-test. i, j, Quantification of Sox9 (i) and VEGF-A (j) fluorescence intensity in immunostained sections of RbpjiΔEC mutants showing decreased signals in mutant growth plates (n = 4 mice from 3 independent litters). Error bars, ± s.e.m. P values, two-tailed unpaired t-test. k, Maximum intensity projection of stained RbpjiΔEC and littermate control tibiae showing decreased VEGF-A immunosignals.
Extended Data Figure 9 Endothelial-cell-dependent regulation of growth plate and bone.
a–c, Quantitative analysis of Fbxw7iΔEC mutant growth plates (a), proliferating zones (b) and maturation/hypertrophy zones (c) relative to control littermates (n = 4 mice from 3 independent litters). Error bars, ± s.e.m. P values, two-tailed unpaired t-test. d, e, Quantitative PCR analysis of Vegfa transcripts level in the bones (without marrow cells) of Notch mutants (RbpjiΔEC and Fbxw7iΔEC) and respective littermate controls (n = 4 mice from 3 independent litters). Error bars, ± s.e.m. P values, two-tailed unpaired t-test. f, g, Maximum intensity projection of stained Fbxw7iΔEC and littermate control tibiae. Note slight reduction of mutant growth plate and maturation/hypertrophy zone (MHZ). EdU (red) labelling (f) marks mitotic chondrocytes in the proliferating zone (PZ). Sox9 (green) immunosignals (g) label maturing and hypertrophic chondrocytes. Dashed lines mark borders of PZ and MHZ. Nuclei, DAPI (blue). h, Maximum intensity projection of tile scanned tibial section of Col1a1-CreERT2 ROSA26-mT/mG double transgenic mice. GFP expression (green) indicates Cre-mediated recombination. i, Maximum intensity projections showing overlapping GFP (green) and Col1α immunostaining (red). j, Maximum intensity projections showing osteoblasts expressing GFP (green). GFP signals were not seen in Emcn immunostained endothelial cells (red). k, Normal arrangement of CD31-stained endothelial columns (C) and arches (A) after Col1a1-CreERT2-mediated inactivation of Rbpj in cells of the osteoblast lineage (RbpjiΔOB). Nuclei, DAPI (blue). l, m, Maximum intensity projections of tile-scanned tibia sections after anti-Emcn (red) immunostaining. Nuclei, DAPI (blue). Note profound disorganization of the Dll4iΔEC metaphyseal vasculature (l) and restoration by simultaneous endothelial-cell-specific overexpression of active Notch in Dll4iΔEC/NICDiOE-EC double mutant mice (m). n, Tile scan confocal images of osteopontin (Opn, green) stained control and Dll4iΔEC/NICDiOE-EC tibiae showing that endothelial NICD expression can rescue trabecular bone defects in the Dll4iΔEC background (see Fig. 2c for comparison). o, Organization of the Emcn-stained metaphyseal vasculature and arrangement of osteoblasts (Osx) in tibia section from 4-week-old control, Dll4iΔEC, NICDiOE-EC, and Dll4iΔEC/NICDiOE-EC mice, as indicated. Nuclei, DAPI (blue). p, Sox9 immunostaining (green) in the epiphyseal region of 4-week-old tibia showing normalized maturing and hypertrophic chondrocytes zone (MHZ) in the growth plate of Dll4iΔEC/NICDiOE-EC mice. Dashed lines mark borders of MHZ. Nuclei, DAPI (blue).
Extended Data Figure 10 Notch and angiocrine Noggin production.
a, Noggin (green) in the wild-type tibial metaphysis was detected in Emcn+ (red) endothelial cells as well as surrounding mesenchymal cells. In contrast, the diaphysis contained Noggin+ haematopoietic cells, whereas only weak staining was seen in sinusoidal blood vessels. b, Panels show higher magnifications of insets in a. Nuclei, DAPI (blue). c, Confocal tile scans of osteopontin-immunostained tibia sections showing partial restoration of trabecular bone formation in 4-week-old RbpjiΔEC mice after administration of recombinant Noggin. Left panels show saline-treated RbpjiΔEC mutants and littermate controls. Nuclei, DAPI (blue). d, e, Noggin treatment restored the bone formation rate (BFR, d) and mineral apposition rate (MAR, e) in RbpjiΔEC long bone to control level (n = 6 mice from 4 independent litters). Data represent mean ± s.e.m. One-way ANOVA was performed along with Bonferroni’s multiple comparison post-hoc test. f, Systemic administration of recombinant Noggin protein reduced the number of Osx+ cells (green) and increased Runx2+ early osteoprogenitors in the RbpjiΔEC metaphysis in comparison to vehicle-treated (saline) mutants. g, Maximum intensity projection of Emcn-immunostained control and RbpjiΔEC tibia sections after treatment with saline or recombinant Noggin, as indicated. Emcn staining intensity was increased in Noggin-treated RbpjiΔEC samples, and the organization of endothelial column and arch structures was partially restored. Dashed lines indicate position of boundaries between endothelial arches (A) and columns (C) as seen in littermate control samples. h, i, Confocal images of VEGF-A (green) immunostained tibia sections showing growth plate chondrocytes in RbpjiΔEC mice after Noggin treatment. Note partial restoration (arrows) of VEGF-A expression in Noggin- but not vehicle control-treated (Saline) RbpjiΔEC mutants (h). Nuclei, DAPI (blue). Quantification data showing fluorescence intensity (in arbitrary units) of VEGF-A expression recovered in the tibial sections of these animals (i) (n = 6 mice from 4 independent litters). Data represent mean ± s.e.m. One-way ANOVA was performed along with Bonferroni’s multiple comparison post-hoc test.
Rights and permissions
About this article
Cite this article
Ramasamy, S., Kusumbe, A., Wang, L. et al. Endothelial Notch activity promotes angiogenesis and osteogenesis in bone. Nature 507, 376–380 (2014). https://doi.org/10.1038/nature13146
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature13146
This article is cited by
-
Photobiomodulation therapy’s impact on angiogenesis and osteogenesis in orthodontic tooth movement: in vitro and in vivo study
BMC Oral Health (2024)
-
MarShie: a clearing protocol for 3D analysis of single cells throughout the bone marrow at subcellular resolution
Nature Communications (2024)
-
Endothelial cells regulate alveolar morphogenesis by constructing basement membranes acting as a scaffold for myofibroblasts
Nature Communications (2024)
-
FOXC1 Promotes Osteoblastic Differentiation of Bone Marrow Mesenchymal Stem Cells via the Dnmt3b/CXCL12 Axis
Biochemical Genetics (2024)
-
LOX-1 regulation of H-type vascular endothelial cell regeneration in hyperglycemia
Acta Diabetologica (2024)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
